Requirement of lmo1930, a Gene in the Menaquinone Biosynthesis Operon, for Esculin Hydrolysis and Lithium Chloride Tolerance in Listeria monocytogenes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Mutant Library Construction and Mutant Screening
2.3. Genetic Complementation
3. Results and Discussion
3.1. Insertional Inactivation of lmo1930 Abolished Esculin Hydrolysis and Resulted in Impaired Growth
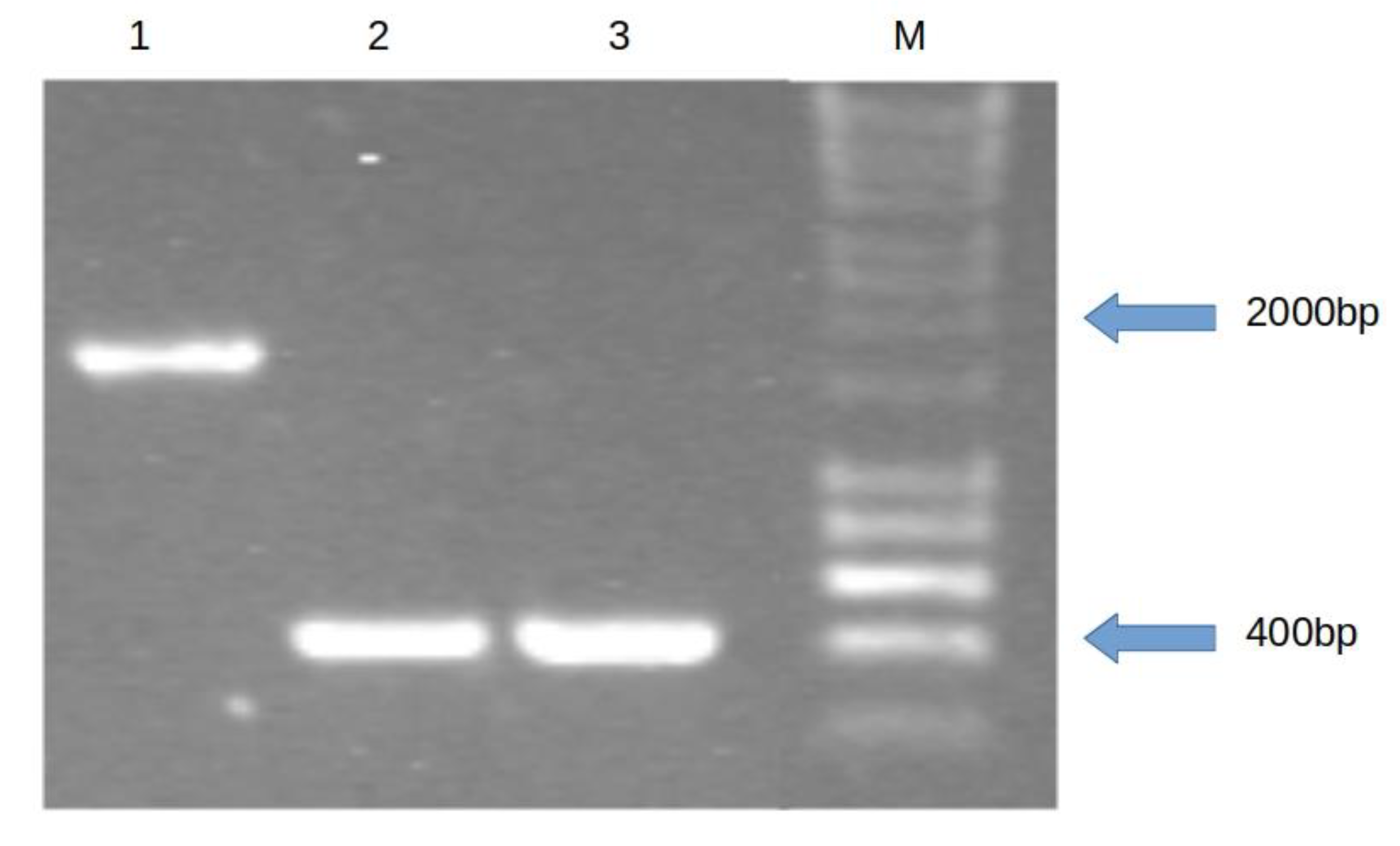
3.2. Genetic Complementation Confirms Role of lmo1930 in Esculin Hydrolysis
3.3. Inactivation of lmo1930 Markedly Reduces Tolerance to LiCl and Compromises Growth of L. Monocytogenes on LiCl-Containing Selective Media Such as MOX and RAPID’L.mono agar
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States-Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Zunabovic, M.; Domig, K.J.; Kneifel, W. Practical relevance of methodologies for detecting and tracing of Listeria monocytogenes in ready-to-eat foods and manufacture environments—A review. LWT Food Sci. Technol. 2011, 44, 351–362. [Google Scholar] [CrossRef]
- Azizoglu, R.O.; Gorski, L.; Kathariou, S. Isolation of Listeria monocytogenes from food and water: Official and experimental protocols. Curr. Protoc. Microbiol. 2014, 33, 1–19. [Google Scholar]
- Magalhães, R.; Mena, C.; Ferreira, V.; Almeida, G.; Silva, J.; Teixeira, P. Traditional Methods for Isolation of Listeria monocytogenes. In Listeria monocytogenes Methods and Protocols; Humana Press: New York, NY, USA, 2014; pp. 15–30. [Google Scholar]
- Anthoni, J.; Humeau, C.; Maia, E.R.; Chebil, L.; Engasser, J.M.; Ghoul, M. Enzymatic synthesis of oligoesculin: Structure and biological activities characterizations. Eur. Food Res. Technol. 2010, 231, 571–579. [Google Scholar] [CrossRef]
- Husain Qadri, S.M.; Johnson, S. Rapid tests for esculin hydrolysis by anaerobic bacteria. Antonie Van Leeuwenhoek 1981, 47, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Hitchins, A.D.; Jinneman, K.; Chen, Y. Detection of Listeria monocytogenes in foods and environmental samples, and enumeration of Listeria monocytogenes in foods. In Bacteriological Analytical Manual; U.S. Food and Drug Administration: Gaithersburg, MD, USA, 2016. [Google Scholar]
- Cao, M.; Bitar, A.P.; Marquis, H. A mariner-based transposition system for Listeria monocytogenes. Appl. Environ. Microbiol. 2007, 73, 2758–2761. [Google Scholar] [CrossRef] [PubMed]
- Parsons, C.; Lee, S.; Jayeola, V.; Kathariou, S. Novel cadmium resistance determinant in Listeria monocytogenes. Appl. Environ. Microbiol. 2017, 83, e02580-16. [Google Scholar] [CrossRef] [PubMed]
- McCollum, J.T.; Cronquist, A.B.; Silk, B.J.; Jackson, K.A.; O’Connor, K.A.; Cosgrove, S.; Gossack, J.P.; Parachini, S.S.; Jain, N.S.; Ettestad, P.; et al. Multistate outbreak of listeriosis associated with cantaloupe. N. Engl. J. Med. 2013, 369, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Gaul, L.K.; Farag, N.H.; Shim, T.; Kingsley, M.A.; Silk, B.J.; Hyytia-Trees, E. Hospital-Acquired Listeriosis Outbreak Caused by Contaminated Diced Celery—Texas, 2010. Clin. Infect. Dis. 2013, 56, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, P.; Qu, T.; Chen, Y.; Hua, X.; Shi, K.; Yu, Y. Identification of Novel Conjugative Plasmids with Multiple Copies of fosB that Confer High-Level Fosfomycin Resistance to Vancomycin-Resistant Enterococci. Front. Microbiol. 2017, 8, 1541. [Google Scholar] [CrossRef] [PubMed]
- Horton, R.M.; Hunt, H.D.; Ho, S.N.; Pullen, J.K.; Pease, L.R. Engineering hybrid genes without the use of restriction enzymes: Gene splicing by overlap extension. Gene 1989, 77, 61–68. [Google Scholar] [CrossRef]
- Lauer, P.; Chow, M.Y.N.; Loessner, M.J.; Portnoy, A.; Calendar, R. Construction, characterization, and use of two Listeria monocytogenes site-specific phage integration vectors. J. Bacteriol. 2002, 184, 4177–4186. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.M.; Cole, M.B.; Legan, J.D.; Slade, L.; Schaffner, D.W. Solute-specific effects of osmotic stress on Staphylococcus aureus. J. Appl. Microbiol. 2005, 98, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.H.; McClain, D. Improved Listeria monocytogenes selective agar. Appl. Environ. Microbiol. 1986, 52, 1215–1217. [Google Scholar] [PubMed]
- Stritzker, J.; Janda, J.; Schoen, C.; Taupp, M.; Pilgrim, S.; Gentschev, I.; Schreier, P.; Geginat, G.; Goebel, W. Growth, virulence, and immunogenicity of Listeria monocytogenes aro mutants. Infect. Immun. 2004, 72, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Arana, A.; Dussurget, O.; Nikitas, G.; Sesto, N.; Guet-Revillet, H.; Balestrino, D.; Loh, E.; Gripenland, J.; Tiensuu, T.; Vaitkevicius, K.; et al. The Listeria transcriptional landscape from saprophytism to virulence. Nature 2009, 459, 950–956. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parsons, C.; Jahanafroozi, M.; Kathariou, S. Requirement of lmo1930, a Gene in the Menaquinone Biosynthesis Operon, for Esculin Hydrolysis and Lithium Chloride Tolerance in Listeria monocytogenes. Microorganisms 2019, 7, 539. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110539
Parsons C, Jahanafroozi M, Kathariou S. Requirement of lmo1930, a Gene in the Menaquinone Biosynthesis Operon, for Esculin Hydrolysis and Lithium Chloride Tolerance in Listeria monocytogenes. Microorganisms. 2019; 7(11):539. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110539
Chicago/Turabian StyleParsons, Cameron, Midya Jahanafroozi, and Sophia Kathariou. 2019. "Requirement of lmo1930, a Gene in the Menaquinone Biosynthesis Operon, for Esculin Hydrolysis and Lithium Chloride Tolerance in Listeria monocytogenes" Microorganisms 7, no. 11: 539. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7110539