Development and Evaluation of a Duo Zaire ebolavirus Real-Time RT-PCR Assay Targeting Two Regions within the Genome

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
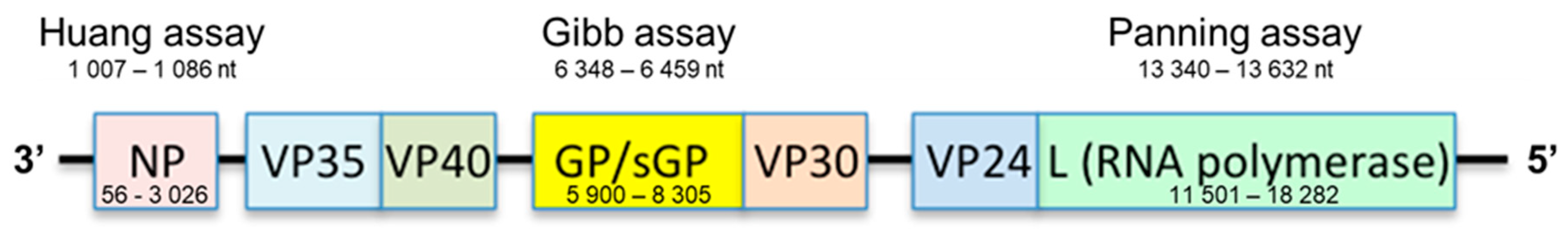
2.1. In Silico Analysis
2.2. Mekambo EBOV RNA
2.3. EBOV RNA Transcript for LoD Calculation
2.4. RT-qPCR Assays
2.5. Specificity
2.6. Cepheid GeneXpert Open Cartridge Development
3. Results
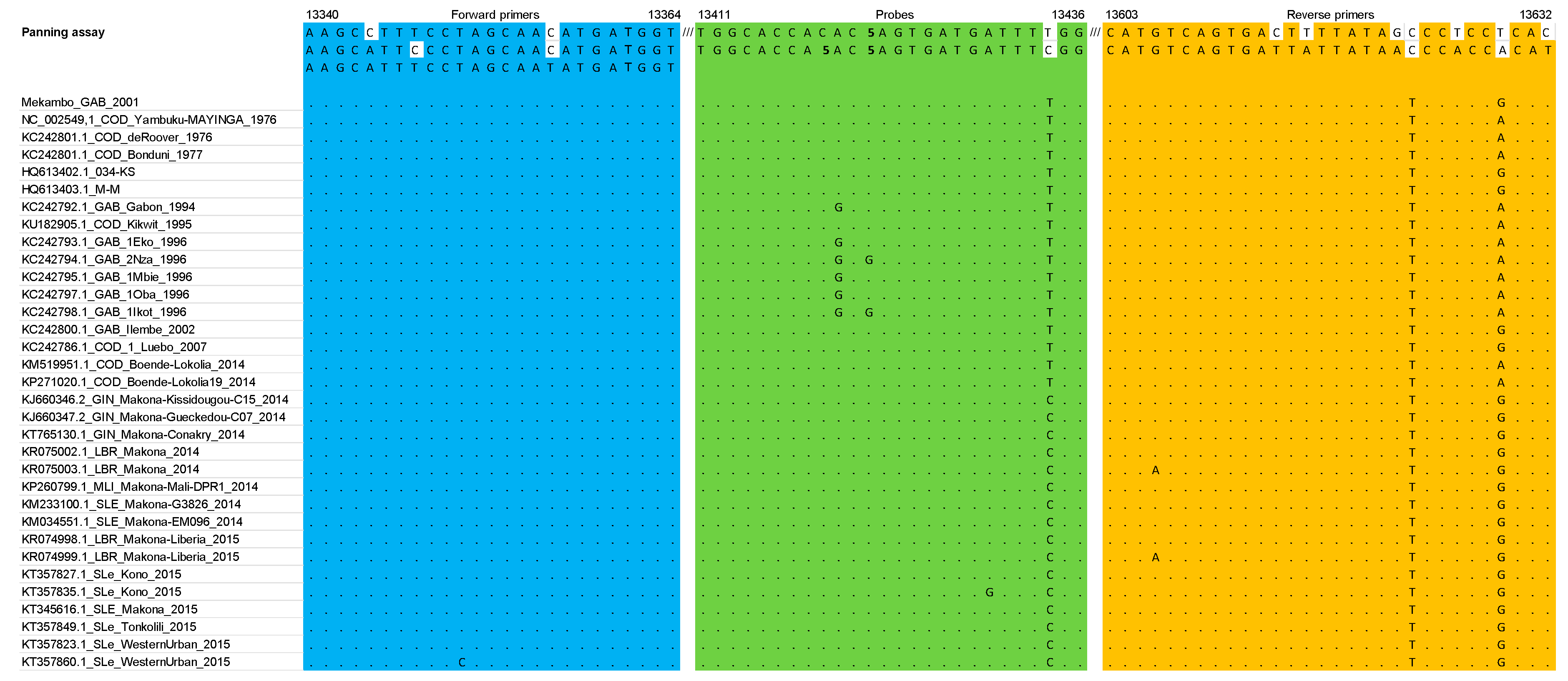
3.1. RT-qPCR Primers and Probe Matched against EBOV Multiple Sequence Alignment: In Silico Analysis
3.2. Study Using the Mekambo EBOV RNA
3.3. LoD Calculation Using the EBOV Synthetic RNA
3.4. Specificity of the Duo Gibb + Huang Assay
3.5. Transfer of the Duo Gibb + Huang Assay onto the Flex-03 Cartridge and Validation on the GeneXpert (Cepheid)
4. Discussion
4.1. In Silico Analysis
4.2. Sensitivity of the Duo Gibb + Huang Assay
4.3. Transfer of the Duo Gibb + Huang Assay onto the Flex-03 Cartridge and Validation on the GeneXpert (Cepheid)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bukreyev, A.A.; Chandran, K.; Dolnik, O.; Dye, J.M.; Ebihara, H.; Leroy, E.M.; Mühlberger, E.; Netesov, S.V.; Patterson, J.L.; Paweska, J.T.; et al. Discussions and Decisions of the 2012–2014 International Committee on Taxonomy of Viruses (ICTV) Filoviridae Study Group, January 2012–June 2013. Arch. Virol. 2014, 159, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Becker, S.; Ebihara, H.; Geisbert, T.W.; Johnson, K.M.; Kawaoka, Y.; Lipkin, W.I.; Negredo, A.I.; Netesov, S.V.; Nichol, S.T.; et al. Proposal for a Revised Taxonomy of the Family Filoviridae: Classification, Names of Taxa and Viruses, and Virus Abbreviations. Arch. Virol. 2010, 155, 2083–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, M.E.; White, M.E.; Dayrit, M.M.; Hayes, C.G.; Ksiazek, T.G.; Burans, J.P. Seroepidemiological Study of Filovirus Related to Ebola in the Philippines. Lancet 1991, 337, 425–426. [Google Scholar] [CrossRef] [Green Version]
- 2014–2016 Ebola Outbreak Distribution in West Africa. Available online: https://www.cdc.gov/vhf/ebola/history/2014-2016-outbreak/distribution-map.html (accessed on 30 September 2019).
- Cherpillod, P.; Schibler, M.; Vieille, G.; Cordey, S.; Mamin, A.; Vetter, P.; Kaiser, L. Ebola Virus Disease Diagnosis by Real-Time RT-PCR: A Comparative Study of 11 Different Procedures. J. Clin. Virol. 2016, 77, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Thirion, L.; Pezzi, L.; Corcostegui, I.; Dubot-Pérès, A.; Falchi, A.; de Lamballerie, X.; Charrel, R.N. Development and Evaluation of a Duo Chikungunya Virus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Viruses 2019, 11, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acid. Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Gibb, T.R.; Norwood, D.A.; Woollen, N.; Henchal, E.A. Development and Evaluation of a Fluorogenic 5′ Nuclease Assay to Detect and Differentiate between Ebola Virus Subtypes Zaire and Sudan. J. Clin. Microbiol. 2001, 39, 4125–4130. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wei, H.; Wang, Y.; Shi, Z.; Raoul, H.; Yuan, Z. Rapid Detection of Filoviruses by Real-Time TaqMan Polymerase Chain Reaction Assays. Virol. Sin. 2012, 27, 273–277. [Google Scholar] [CrossRef]
- Panning, M.; Laue, T.; Olschlager, S.; Eickmann, M.; Becker, S.; Raith, S.; Courbot, M.-C.G.; Nilsson, M.; Gopal, R.; Lundkvist, A.; et al. Diagnostic Reverse-Transcription Polymerase Chain Reaction Kit for Filoviruses Based on the Strain Collections of All European Biosafety Level 4 Laboratories. J. Infect. Dis. 2007, 196, S199–S204. [Google Scholar] [CrossRef] [Green Version]
- Biava, M.; Colavita, F.; Marzorati, A.; Russo, D.; Pirola, D.; Cocci, A.; Petrocelli, A.; Delli Guanti, M.; Cataldi, G.; Kamara, T.A.; et al. Evaluation of a Rapid and Sensitive RT-QPCR Assay for the Detection of Ebola Virus. J. Virol. Methods 2018, 252, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Dedkov, V.G.; Magassouba, N.; Safonova, M.V.; Bodnev, S.A.; Pyankov, O.V.; Camara, J.; Sylla, B.; Agafonov, A.P.; Maleev, V.V.; Shipulin, G.A. Sensitive Multiplex Real-Time RT-QPCR Assay for the Detection of Filoviruses. Health Secur. 2018, 16, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Ro, Y.-T.; Ticer, A.; Carrion, R.; Patterson, J.L. Rapid Detection and Quantification of Ebola Zaire Virus by One-Step Real-Time Quantitative Reverse Transcription-Polymerase Chain Reaction. Microbiol. Immunol. 2017, 61, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieger, T.; Kerber, R.; El Halas, H.; Pallasch, E.; Duraffour, S.; Günther, S.; Ölschläger, S. Evaluation of RealStar Reverse Transcription-Polymerase Chain Reaction Kits for Filovirus Detection in the Laboratory and Field. J. Infect. Dis. 2016, 214, S243–S249. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Sun, Y.; Kargbo, B.; Zhang, C.; Feng, H.; Lu, H.; Liu, W.; Wang, C.; Hu, Y.; Deng, Y.; et al. Detection of Zaire Ebola Virus by Real-Time Reverse Transcription-Polymerase Chain Reaction, Sierra Leone, 2014. J. Virol. Methods 2015, 222, 62–65. [Google Scholar] [CrossRef]
- Jääskeläinen, A.J.; Moilanen, K.; Aaltonen, K.; Putkuri, N.; Sironen, T.; Kallio-Kokko, H.; Vapalahti, O. Development and Evaluation of a Real-Time EBOV-L-RT-QPCR for Detection of Zaire Ebolavirus. J. Clin. Virol. 2015, 67, 56–58. [Google Scholar] [CrossRef]
- Trombley, A.R.; Wachter, L.; Garrison, J.; Buckley-Beason, V.A.; Jahrling, J.; Hensley, L.E.; Schoepp, R.J.; Norwood, D.A.; Goba, A.; Fair, J.N.; et al. Comprehensive Panel of Real-Time TaqMan Polymerase Chain Reaction Assays for Detection and Absolute Quantification of Filoviruses, Arenaviruses, and New World Hantaviruses. Am. J. Trop. Med. Hyg. 2010, 82, 954–960. [Google Scholar] [CrossRef]
- Drosten, C.; Göttig, S.; Schilling, S.; Asper, M.; Panning, M.; Schmitz, H.; Günther, S. Rapid Detection and Quantification of RNA of Ebola and Marburg Viruses, Lassa Virus, Crimean-Congo Hemorrhagic Fever Virus, Rift Valley Fever Virus, Dengue Virus, and Yellow Fever Virus by Real-Time Reverse Transcription-PCR. J. Clin. Microbiol. 2002, 40, 2323–2330. [Google Scholar] [CrossRef] [Green Version]
- Dedkov, V.G.; Magassouba, N.F.; Safonova, M.V.; Deviatkin, A.A.; Dolgova, A.S.; Pyankov, O.V.; Sergeev, A.A.; Utkin, D.V.; Odinokov, G.N.; Safronov, V.A.; et al. Development and Evaluation of a Real-Time RT-PCR Assay for the Detection of Ebola Virus (Zaire) during an Ebola Outbreak in Guinea in 2014-2015. J. Virol. Methods 2016, 228, 26–30. [Google Scholar] [CrossRef]
- Templer, S.P.; Seiverth, B.; Baum, P.; Stevens, W.; Seguin-Devaux, C.; Carmona, S. Improved Sensitivity of a Dual-Target HIV-1 Qualitative Test for Plasma and Dried Blood Spots. J. Clin. Microbiol. 2016, 54, 1877–1882. [Google Scholar] [CrossRef] [Green Version]
- Sizmann, D.; Glaubitz, J.; Simon, C.O.; Goedel, S.; Buergisser, P.; Drogan, D.; Hesse, M.; Kröh, M.; Simmler, P.; Dewald, M.; et al. Improved HIV-1 RNA Quantitation by COBAS AmpliPrep/COBAS TaqMan HIV-1 Test, v2.0 Using a Novel Dual-Target Approach. J. Clin. Virol. 2010, 49, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Damond, F.; Avettand-Fenoel, V.; Collin, G.; Roquebert, B.; Plantier, J.C.; Ganon, A.; Sizmann, D.; Babiel, R.; Glaubitz, J.; Chaix, M.L.; et al. Evaluation of an Upgraded Version of the Roche Cobas AmpliPrep/Cobas TaqMan HIV-1 Test for HIV-1 Load Quantification. J. Clin. Microbiol. 2010, 48, 1413–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinsky, B.A.; Sahoo, M.K.; Sandlund, J.; Kleman, M.; Kulkarni, M.; Grufman, P.; Nygren, M.; Kwiatkowski, R.; Baron, E.J.; Tenover, F.; et al. Analytical Performance Characteristics of the Cepheid GeneXpert Ebola Assay for the Detection of Ebola Virus. PLoS ONE 2015, 10, e0142216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen van Vuren, P.; Grobbelaar, A.; Storm, N.; Conteh, O.; Konneh, K.; Kamara, A.; Sanne, I.; Paweska, J.T. Comparative Evaluation of the Diagnostic Performance of the Prototype Cepheid GeneXpert Ebola Assay. J. Clin. Microbiol. 2016, 54, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semper, A.E.; Broadhurst, M.J.; Richards, J.; Foster, G.M.; Simpson, A.J.; Logue, C.H.; Kelly, J.D.; Miller, A.; Brooks, T.J.; Murray, M.; et al. Performance of the GeneXpert Ebola Assay for Diagnosis of Ebola Virus Disease in Sierra Leone: A Field Evaluation Study. PLoS Med. 2016, 13, e1001980. [Google Scholar] [CrossRef]
- Raftery, P.; Condell, O.; Wasunna, C.; Kpaka, J.; Zwizwai, R.; Nuha, M.; Fallah, M.; Freeman, M.; Harris, V.; Miller, M.; et al. Establishing Ebola Virus Disease (EVD) diagnostics using GeneXpert technology at a mobile laboratory in Liberia: Impact on outbreak response, case management and laboratory systems strengthening. PLoS Negl. Trop. Dis. 2018, 12, e0006135. [Google Scholar] [CrossRef]
- Mbala-Kingebeni, P.; Villabona-Arenas, C.J.; Vidal, N.; Likofata, J.; Nsio-Mbeta, J.; Makiala-Mandanda, S.; Mukadi, D.; Mukadi, P.; Kumakamba, C.; Djokolo, B.; et al. Rapid Confirmation of the Zaire Ebola Virus in the Outbreak of the Equateur Province in the Democratic Republic of Congo: Implications for Public Health Interventions. Clin. Infect. Dis. 2019, 68, 330–333. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Target | Amplicon | Primers/Probe | Sequence (5′-3′) |
|---|---|---|---|---|
| Gibb | GP | 112 bp | EBOGP1D-fwd | TGGGCTGAAAAYTGCTACAATC |
| EBOGP1D-rev | GTGCCGSTATGTKCACAAAG | |||
| EBOGP1DZ-Prb | FAM-CTACCAGCAGCGCCAGACGG-TAMRA | |||
| Huang | NP | 80 bp | enp-F | GCAGAGCAAGGACTGATTCA |
| Enp-F2 | GCAGAGCAAGGACTGATACA | |||
| enp-R | ATTTTCCGTTTGATGCGAAC | |||
| enp-Probe | FAMa-CAACAGCTTGGCAATCAGTTGGACA-TAMRA | |||
| Panning | L | 293 bp | FiloA2.2 | AAGCCTTTCCTAGCAACATGATGGT |
| FiloA2.3 | AAGCATTCCCTAGCAACATGATGGT | |||
| FiloA2.4 | AAGCATTTCCTAGCAATATGATGGT | |||
| FiloB | CATGTCAGTGATTATTATAACCCACCACAT | |||
| Filo B-Prime | CATGTCAGTGATTATTATAAYCCACCRCAT | |||
| Filo B-Ra | CATGTCAGTGACTTTTATAGCCCTCCTCAC | |||
| FAMEBOSu | FAMb-TGGCACCAIACIAGTGATGATTTCGG-BHQ1 | |||
| FAMEBOg | FAMb-TGGCACCACACIAGTGATGATTTTGG-BHQ1 |
| Gibb et al. [9] | Huang et al. [10] | Panning et al. [11] | Standard Protocol Mekambo EBOV RNA | Standard Protocol EBOV Synthetic RNA | RealStar® Filovirus Altona Diagnostics | |
|---|---|---|---|---|---|---|
| Reverse Transcription | 55 °C/45 min | 50 °C/30 min | 50 °C/30 min | 50 °C/15 min | 50 °C/15 min | 55 °C/20 min |
| Denaturation | 94 °C/1 min | 94 °C/5 min | 95 °C/1.5 min | 95 °C/2 min | 95 °C/2 min | 95 °C/2 min |
| Cycling | 40 | 45 | 45 | 40 | 40 | 45 |
| Denaturation | 94 °C/15 s | 94 °C/15 s | 95 °C/15 s | 95 °C/15 s | 95 °C/15 s | 95 °C/15 s |
| Amplification | 60 °C/30 s | 60 °C/1 min | 72 °C/20 s | 60 °C/1 min | 60 °C/1 min | 72 °C/15 s |
| RNA volume | 5 µL | 1–3 µL | 3 µL | 5 µL | 10 µL | 10 µL |
| Total volume | 50 µL | 23–25 µL | 25 µL | 25 µL | 30 µL | 30 µL |
| Primers concentration | 0.5 µM | 0.4 µM | 0.2 µM Forward 0.3 µM Reverse | 0.4 µM Gibb 0.4 µM Huang 0.2 µM Panning | 0.625 µM Gibb 0.5 µM Huang | - |
| Probes concentration | 0.2 µM | 0.1 µM | 0.0667 µM | 0.16 µM Gibb and Huang | 0.25 µM Gibb | - |
| 0.2 µM each Panning | 0.125 µM Huang |
| Genus | Virus | Acronyms | Strain (Viral Load TCID50/mL) | Reference (a) |
|---|---|---|---|---|
| Phlebovirus | Toscana virus | TOSV | UVE/TOSV/2014/FR/5904 (10 8,22) | 001v-02452 |
| Sandfly Fever Sicilian virus | SFSV | UVE/SFSV/1943/IT/Sabin (10 6,82) | 001v-EVA77 | |
| Flavivirus | Japanese encephalitis virus | JEV | UVE/JEV/2009/LA/CNS769 (10 5,57) | 001v-02217 |
| Saint-Louis encephalitis virus | SLEV | UVE/SLEV/UNK/US/MSI-7 (10 4,82) | 001v-EVA128 | |
| Tick-borne encephalitis virus | TBEV | UVE/TBEV/2013/FR/32.11 WT-PCR (10 8,82) | 001v-02352 | |
| West-Nile virus | WNV | UVE/WNV/2008/US/R94224 (10 7,32) | 001v-02224 | |
| Yellow Fever virus | YFV | UVE/YFV/UNK/XX/French neurotropic R94224 (10 7,32) | 001v-02226 | |
| Usutu virus | USUV | UVE/USUV/1959/ZA/SAAR-1776 (10 5,32) | 001v-EVA138 | |
| Murray Valley virus | MVEV | UVE/MVEV/UNK/AU/3329 (10 4,32) | 001v-EVA145 | |
| Zika virus | ZIKV | UVE/ZIKV/1947/UG/MR766 (10 4,32) | 001v-EVA143 | |
| Dengue virus | DENV-1 | UVE/DENV-1/2013/NC/CNR_17132 (10 7,57) | 001v-03151 | |
| Alphavirus | Venezuelan equine encephalitis virus | VEEV | UVE/VEEV/UNK/XX/TC83 vaccine (10 9,42) | 001v-EVA1459 |
| Western equine encephalitis virus | WEEV | UVE/WEEV/UNK/XX/47a (10 8,32) | 001v-EVA1479 | |
| Eastern equine encephalitis virus | EEEV | UVE/EEEV/1999/XX/H178_99 (10 7,82) | 001v-EVA1480 | |
| O’nyong-nyong virus | ONNV | UVE/ONNV/UNK/SN/Dakar 234 (10 4,22) | 001v-EVA1044 | |
| Chikungunya virus | CHIKV | UVE/CHIKV/2017/FR/45625-26 (10 6,16) | 001v-03433 | |
| Semliki Forest virus | SFV | UVE/SFV/UNK/XX/1745 (10 4,42) | 001v-02468 | |
| Sindbis virus | SINV | UVE/SINV/UNK/EG/Egypt 339 (10 4,32) | 001v-02469 | |
| Filovirus | Marburg virus | MBGV | Popp | n/a |
| Marburg virus | MBGV | Musoke | n/a | |
| Nairovirus | Crimean-Congo hemorrhagic fever virus | CCHF | Unk | n/a |
| Replicate | RNA Copies/Reaction | ||||
|---|---|---|---|---|---|
| 16.8 | 12.6 | 8.4 | 4.2 | 1.7 | |
| Flex-C#1 | 37.1 | 38.7 | 39.6 | negative | negative |
| Flex-C#2 | 36.3 | 36.0 | 38.9 | 38.0 | negative |
| Flex-C#3 | 36.4 | 36.9 | 39.2 | negative | negative |
| Flex-C#4 | 38.4 | 37.6 | 37.9 | negative | negative |
| Flex-C#5 | 36.2 | 38.2 | negative | 36.7 | negative |
| Flex-C#6 | 36.5 | 39.3 | negative | 38.0 | negative |
| Flex-C#7 | 39.7 | 37.8 | negative | negative | negative |
| Flex-C#8 | 36.1 | 36.2 | negative | negative | negative |
| Mean | 37.1 (1.3) | 37.6 (1.2) | 38.9 (0.6) | 37.6 (0.6) | - |
| GIBB et al. [9] | Replicate | RNA Copies/µL | ||||||
| 1 | 0.75 | 0.5 | 0.2 | 0.1 | 0.075 | 0.05 | ||
| 1 | 36.57 | 36.66 | 37.28 | 38.76 | >40 | >40 | >40 | |
| 2 | 36.59 | 35.99 | 37.25 | 38.43 | >40 | >40 | >40 | |
| 3 | 37.02 | 36.62 | 36.98 | >40 | 38.36 | >40 | >40 | |
| 4 | 36.47 | 37.07 | >40 | 38.27 | >40 | >40 | >40 | |
| 5 | 38.47 | 36.17 | >40 | 38.27 | >40 | >40 | >40 | |
| 6 | 37.49 | >40 | 38.54 | >40 | 38.54 | >40 | >40 | |
| 7 | >40 | 38.53 | 37.59 | >40 | 38.52 | 39.85 | >40 | |
| Mean Ct (SD) | 37.10 (0.7) | 36.84 (0.8) | 37.53 (0.5) | 38.43 (0.2) | 38.47 (0.1) | 39.85 | >40 | |
| HUANG et al. [10] | Replicate | RNA Copies/µL | ||||||
| 1 | 0.75 | 0.5 | 0.2 | 0.1 | 0.075 | 0.05 | ||
| 1 | 37.59 | >40 | 38.14 | >40 | 38.60 | >40 | >40 | |
| 2 | 37.92 | 38.02 | >40 | 40 | >40 | >40 | >40 | |
| 3 | 37.24 | 38.42 | 39.62 | >40 | 39.09 | >40 | >40 | |
| 4 | 37.30 | 38.48 | >40 | >40 | >40 | 37.95 | >40 | |
| 5 | 38.35 | 37.39 | >40 | >40 | >40 | 39.98 | >40 | |
| 6 | 37.89 | >40 | 38.07 | >40 | >40 | 39.98 | >40 | |
| 7 | 37.48 | 37.48 | 39.13 | 39.31 | 39.92 | >40 | >40 | |
| Mean Ct (SD) | 37.68 (0.4) | 37.96 (0.5) | 38.74 (0.7) | 39.66 | 39.20 (0.5) | 39.30 (1.0) | >40 | |
| DUO GIBB + HUANG (this study) | Replicate | RNA Copies/µL | ||||||
| 1 | 0.75 | 0.5 | 0.2 | 0.1 | 0.075 | 0.05 | ||
| 1 | NT | 32.14 | 33.04 | 34.89 | 36.40 | 32.98 | >40 | |
| 2 | NT | 33.01 | 32.78 | 34.93 | 36.24 | 33.17 | >40 | |
| 3 | NT | 32.87 | 33.10 | 34.03 | >40 | 33.80 | 38.27 | |
| 4 | NT | 32.83 | 33.20 | 34.88 | >40 | 33.48 | >40 | |
| 5 | NT | 32.77 | 36.61 | 34.67 | 37.45 | > 40 | >40 | |
| 6 | NT | 32.28 | 33.53 | 35.63 | 35.34 | 32.95 | >40 | |
| 7 | NT | 32.71 | 32.74 | >40 | 35.43 | >40 | >40 | |
| 8 | NT | 32.59 | 32.84 | 34.64 | 35.18 | 33.41 | >40 | |
| 9 | NT | 31.98 | 33.40 | 34.04 | 35.39 | 33.97 | >40 | |
| 10 | NT | 32.44 | 33.12 | 35.12 | >40 | 33.01 | >40 | |
| 11 | NT | 33.11 | 33.33 | 34.67 | 35.88 | 33.83 | >40 | |
| 12 | NT | 32.27 | 32.73 | 35.36 | 35.36 | 33.56 | >40 | |
| 13 | NT | 32.19 | 33.45 | >40 | 38.37 | 36.71 | >40 | |
| 14 | NT | 32.41 | 33.04 | 34.29 | >40 | 34.74 | >40 | |
| 15 | NT | 33.06 | 33.29 | 34.48 | 36.04 | 34.18 | >40 | |
| 16 | NT | 32.47 | 32.41 | 35.69 | >40 | >40 | 34.65 | |
| 17 | NT | 32.07 | 32.93 | 34.70 | >40 | 33.18 | >40 | |
| 18 | NT | 32.22 | 33.15 | >40 | 35.86 | 32.29 | >40 | |
| 19 | NT | 34.00 | 33.46 | 36.39 | 35.78 | 36.10 | >40 | |
| 20 | NT | 32.38 | 34.32 | >40 | 33.92 | 33.17 | >40 | |
| 21 | NT | 33.12 | 33.05 | 34.57 | 36.72 | >40 | > 40 | |
| 22 | NT | 32.44 | 32.55 | 34.31 | >40 | >40 | >40 | |
| 23 | NT | 33.08 | 32.23 | >40 | > 40 | >40 | >40 | |
| Mean Ct (SD) | 32.63 (0.5) | 33.23 (0.8) | 34.85 (0.6) | 35.96 (1.0) | 33.80 (1.1) | 36.46 (1.8) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thirion, L.; Charrel, R.N.; Boehmann, Y.; Corcostegui, I.; Raoul, H.; de Lamballerie, X. Development and Evaluation of a Duo Zaire ebolavirus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Microorganisms 2019, 7, 652. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120652
Thirion L, Charrel RN, Boehmann Y, Corcostegui I, Raoul H, de Lamballerie X. Development and Evaluation of a Duo Zaire ebolavirus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Microorganisms. 2019; 7(12):652. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120652
Chicago/Turabian StyleThirion, Laurence, Remi N. Charrel, Yannik Boehmann, Iban Corcostegui, Hervé Raoul, and Xavier de Lamballerie. 2019. "Development and Evaluation of a Duo Zaire ebolavirus Real-Time RT-PCR Assay Targeting Two Regions within the Genome" Microorganisms 7, no. 12: 652. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120652