Characterization of Salinivibrio socompensis sp. nov., A New Halophilic Bacterium Isolated from the High-Altitude Hypersaline Lake Socompa, Argentina

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Culture Media and Growth Conditions
2.3. Taxophylogenomic Characterization
2.3.1. Phylogenetic Analysis Based on 16S rRNA Gene Sequence Comparison
2.3.2. Phylogenomic Comparative Analysis
2.3.3. Average Nucleotide Identity (ANI) and n-silico DNA–DNA Hybridization (DDH)
2.4. Phenotypic Characterization
2.4.1. Morphology and Motility
2.4.2. Physiological Characteristics
2.4.3. Biochemical Characteristics
2.4.4. Nutritional Characteristics
2.4.5. Chemotaxonomic Characterization
3. Results and Discussion
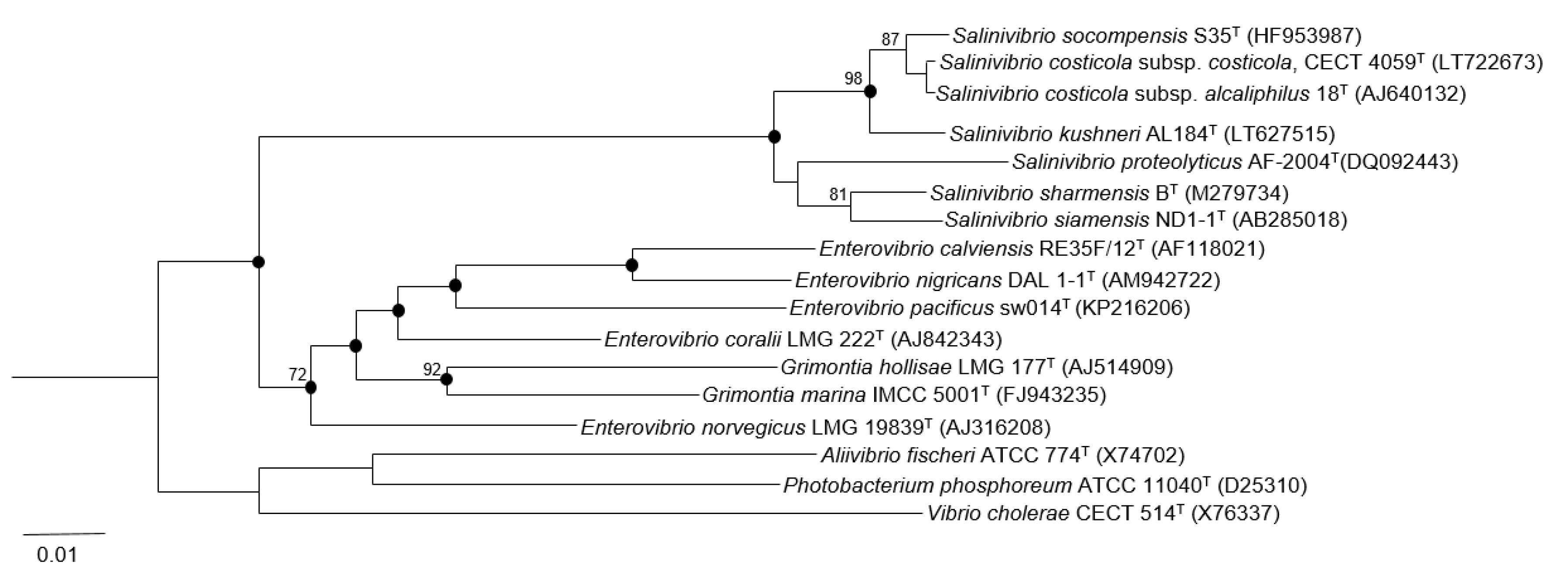
3.1. Phylogenetic Analysis Based on 16S rRNA Gene Sequence Comparison
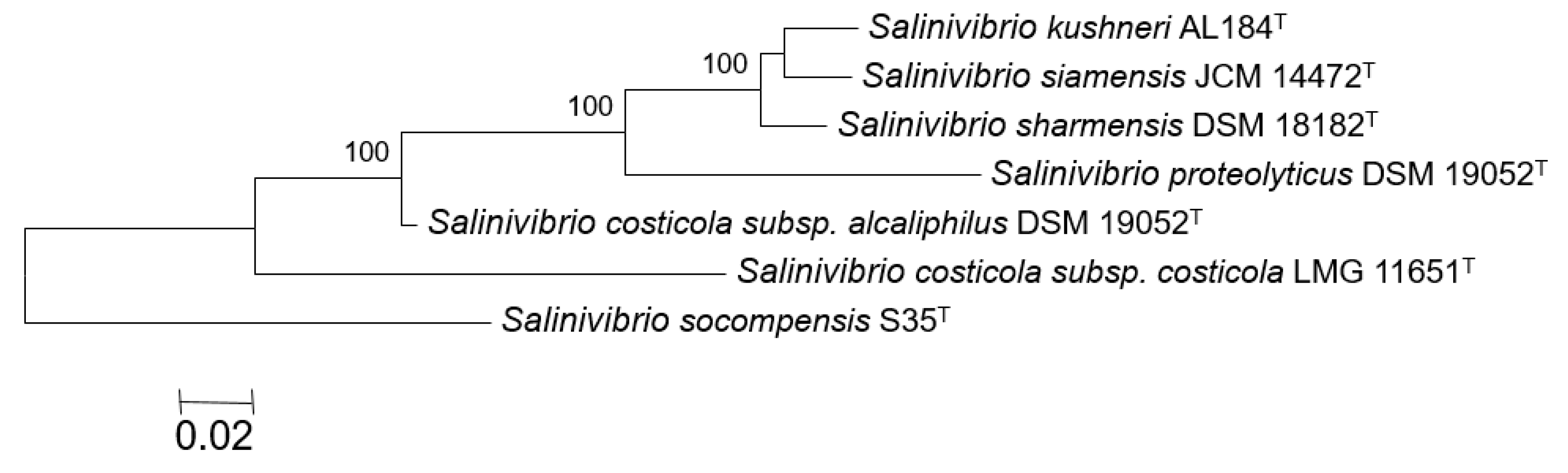
3.2. Phylogenomic Comparative Analysis
3.3. ANI Values and In-Silico DNA–DNA Hybridization (GGDC)
3.4. Phenotypic Characterization
3.5. Chemotaxonomic Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ventosa, A. Unusual micro-organisms from unusual habitats: Hypersaline environments. In Prokaryotic Diversity: Mechanisms and Significance; Cambridge University Press: Cambridge, UK, 2006; pp. 223–254. [Google Scholar]
- de la Haba, R.R.; Sánchez-Porro, C.; Márquez, M.C.; Ventosa, V. Taxonomy of halophiles. In Extremophiles Handbook; Springer: Heidelberg, Germany, 2010; pp. 255–309. [Google Scholar]
- Amoozegar, M.A.; Siroosi, M.; Atashgahi, S.; Smidt, H.; Ventosa, A. Systematics of haloarchaea and biotechnological potential of their hydrolytic enzymes. Microbiology 2017, 163, 623–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira Obayashi, U.; Nobutsugu Hiraoka, M.; Keiko Kita, K.; Hiroshi Nakajima, O. Inventors process for producing restriction enzyme. U.S. Patent US 4724209, 9 February 1988. [Google Scholar]
- Fu, X.T.; Kim, S.M. Agarase: Review of major sources, categories, purification method, enzyme characteristics and applications. Mar. Drugs 2010, 8, 200–218. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; Geueke, B.; Delgado, O.; Coleman, J.; Hatti-Kaul, R. β-galactosidase from a cold-adapted bacterium: Purification, characterization and application for lactose hydrolysis. Appl. Microbiol. Biotechnol. 2002, 58, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A. Salinivibrio. In Bergey’s Manual of Systematics of Archaea and Bacteria; Springer: New York, NY, USA, 2005; Volume 2, pp. 552–556. [Google Scholar]
- Smith, F.B. An investigation of a taint in rib bones of bacon. The determination of halophilic vibrios (n. spp.). Proc. R. Soc. Qd. 1938, 49, 29–52. [Google Scholar]
- Ventosa, A.; Quesada, E.; Rodríguez-Valera, F.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Numerical taxonomy of moderately halophilic Gram-negative rods. J. Gen. Microbiol. 1982, 128, 1959–1968. [Google Scholar] [CrossRef]
- Kamekura, M.; Wallace, R.; Hipkiss, A.R.; Kushner, D.J. Growth of Vibrio costicola and other moderate halophiles in a chemically defined minimal medium. Can. J. Microbiol. 1985, 31, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Diversity of halophilic microorganisms: Environments, phylogeny, physiology, and applications. J. Ind. Microbiol. Biotechnol. 2002, 28, 56–63. [Google Scholar] [CrossRef]
- Oren, A. Halophilic Microorganisms and their Environments. In Cellular Origin and Life in Extreme Habitats, vol. 5; Kluwer Academic Press: London, UK, 2002. [Google Scholar]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of moderately halophilic aerobic bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar]
- Mellado, E.; Moore, E.R.B.; Nieto, J.J.; Ventosa, A. Analysis of 16S rRNA gene sequences of Vibrio costicola strains: Description of Salinivibrio costicola gen. nov., comb. nov. Int. J. Syst. Bacteriol. 1996, 46, 817–821. [Google Scholar] [CrossRef]
- Huang, C.Y.; García, J.L.; Patel, B.K.C.; Cayol, J.L.; Baresi, L.; Mah, R.A. Salinivibrio costicola subsp. vallismortis subsp. nov., a halotolerant facultative anaerobe from Death Valley, and emended description of Salinivibrio costicola. Int. J. Syst. Evol. Microbiol. 2000, 50, 615–622. [Google Scholar]
- Romano, I.; Gambacorta, A.; Lama, L.; Nicolaus, B.; Giordano, A. Salinivibrio costicola subsp. alcaliphilus subsp. nov., a haloalkaliphilic aerobe from Campania Region (Italy). Syst. Appl. Microbiol. 2005, 28, 34–42. [Google Scholar] [PubMed]
- Amoozegar, M.A.; Schumann, P.; Hajighasemi, M.; Fatemi, A.Z.; Karbalaei-Heidari, H.R. Salinivibrio proteolyticus sp. nov., a moderately halophilic and proteolytic species from a hypersaline lake in Iran. Int. J. Syst. Evol. Microbiol. 2008, 58, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Chamroensaksri, N.; Tanasupawat, S.; Akaracharanya, A.; Visessanguan, W.; Kudo, T.; Itoh, T. Salinivibrio siamensis sp. nov., from fermented fish (pla-ra) in Thailand. Int. J. Syst. Evol. Microbiol. 2009, 59, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Romano, I.; Orlando, P.; Gambacorta, A.; Nicolaus, B.; Dipasquale, L.; Pascual, J.; Giordano, A.; Lama, L. Salinivibrio sharmensis sp. nov., a novel haloalkaliphilic bacterium from a saline lake in Ras Mohammed Park (Egypt). Extremophiles 2011, 15, 213–220. [Google Scholar] [CrossRef] [PubMed]
- López-Hermoso, C.; de la Haba, R.R.; Sánchez-Porro, C.; Ventosa, A. Salinivibrio kushneri sp. nov., a moderately halophilic bacterium isolated from salterns. Syst. Appl. Microbiol. 2018, 41, 159–166. [Google Scholar] [CrossRef]
- López-Hermoso, C.; de la Haba, R.R.; Sánchez-Porro, C.; Ventosa, A. Emended description of Salinivibrio proteolyticus, including Salinivibrio costicola subsp. vallismortis and five new isolates. Int. J. Syst. Evol. Microbiol. 2018, 68, 1599–1607. [Google Scholar] [CrossRef]
- López-Hermoso, C.; de la Haba, R.R.; Sánchez-Porro, C.; Papke, R.T.; Ventosa, A. Assessment of MultiLocus Sequence Analysis as a valuable tool for the classification of the genus Salinivibrio. Front. Microbiol. 2017, 8, 1107. [Google Scholar]
- Gorriti, M.F.; Dias, G.M.; Chimetto, L.A.; Trindade-Silva, A.E.; Silva, B.S.; Mesquita, M.M.; Gregoracci, G.B.; Farias, M.E.; Thompson, C.C.; Thompson, F.L. Genomic and phenotypic attributes of novel salinivibrios from stromatolites, sediment and water from a high altitude lake. BMC Genom. 2014, 15, 473. [Google Scholar] [CrossRef]
- Toneatti, D.M.; Albarracín, V.H.; Flores, M.R.; Polerecky, L.; Farías, M.E. Stratified bacterial diversity along physico-chemical gradients in high-altitude modern stromatolites. Front. Microbiol. 2017, 8, 646. [Google Scholar] [CrossRef]
- Kurth, D.; Amadio, A.; Ordoñez, O.F.; Albarracín, V.H.; Gärtnerm, W.; Farías, M.E. Arsenic metabolism in high altitude modern stromatolites revealed by metagenomic analysis. Sci. Rep. 2017, 7, 1024. [Google Scholar] [CrossRef]
- Farias, M.E.; Rascovan, N.; Toneatti, D.M.; Albarracin, V.H.; Flores, M.R.; Poiré, D.G.; Collavino, M.M.; Aguilar, A.M.; Vazquez, M.P.; Polerecky, L. The discovery of stromatolites developing at 3570 m above sea level in a high-altitude volcanic lake Socompa, Argentinean Andes. PLoS ONE 2013, 8, e53497. [Google Scholar] [CrossRef] [PubMed]
- Allwood, A.C.; Walter, M.R.; Kamber, B.S.; Marshall, C.P.; Burch, I.W. Stromatolite reef from the early Archaean era of Australia. Nature 2006, 441, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Burne, R.V.; Moore, L.S. Microbialites; organosedimentary deposits of benthic microbial communities. Palaios 1987, 2, 241–254. [Google Scholar] [CrossRef]
- Schopf, J.W. Fossil evidence of Archaean life. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 869–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [PubMed]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar; Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; et al. ARB: A software environment for sequence data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Fitch, W.M. Toward defining the course of evolution: Minimum change for a specific tree topology. Syst. Biol. 1971, 20, 406–416. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ. Prepr. 2016, 4, e1900v1. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2002, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, H.; Hasegawa, M. Multiple comparisons of loglikelihoods with applications to phylogenetic inference. Mol. Biol. Evol. 1999, 16, 1114–1116. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Lee, I.; Kim, Y.O.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2015, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Cowan, S.T.; Steel, K.J. Manual for the Identification of Medical Bacteria, 3rd ed.; Cambridge University Press: London, UK, 1965. [Google Scholar]
- Kovacs, N. Identification of Pseudomonas pyocyanea by the oxidase reaction. Nature 1956, 178, 703. [Google Scholar] [CrossRef] [PubMed]
- Koser, S.A. Utilization of the salts of organic acids by the colon-aerogenes group. J. Bacteriol. 1923, 8, 493–523. [Google Scholar] [PubMed]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids; MIDI Technical Note 101; MIDI Inc.: Newark, DE, USA, May 1990. [Google Scholar]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Osawa, I.; Adachi, H.; Kawada, M. Vibrio japonicus sp. nov., a novel member of the Nereis clade in the genus Vibrio isolated from the coast of Japan. PLoS ONE 2017, 12, e0172164. [Google Scholar] [CrossRef] [PubMed]
- Paek, J.; Shin, J.H.; Shin, Y.; Park, I.S.; Kim, H.; Kook, J.K.; Kang, S.S.; Kim, D.S.; Park, K.H.; Chang, Y.H. Vibrio injenensis sp. nov., isolated from human clinical specimens. Antonie Van Leeuwenhoek 2017, 110, 145–152. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ANIb | ANIm | orthoANI | GGDC | |
|---|---|---|---|---|
| Strains | Strain S35T | |||
| S. costicola subsp. costicola LMG 11651T | 87.8 | 88.9 | 88.1 | 35 |
| S. costicola subsp. alcaliphilus DSM 16359T | 87.9 | 88.6 | 88.2 | 35 |
| S. proteolyticus DSM 19052T | 78.1 | 84.6 | 78.6 | 22 |
| S. sharmensis DSM 18182T | 80.4 | 84.5 | 80.8 | 23 |
| S. siamensis JCM 14472T | 80.6 | 84.6 | 80.9 | 24 |
| S. kushneri AL184T | 80.6 | 84.7 | 81.0 | 24 |
| Characteristic | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| NaCl range (% w/v) | 3–20 | 0.5–20 a | 2–25 b | 1–17 c | 1–22 d | 6–16 e | 2–20 f |
| NaCl optimum (% w/v) | 7–7.5 | 10 a | 10 b | 5 c | 10 d | 10 e | 7.5 f |
| pH range | 6–10 | 5–10 a | 7–10.5 b | 5–9.5 c | 5–9 d | 6–10 e | 5–10 f |
| pH optimum | 8 | 7.5 a | 9 b | 8 c | 8 d | 9 e | 7.2–7.4 f |
| Temperature range (°C) | 10–42 | 5–45 a | 10–40 b | 10–45 c | 10–47 d | 25–40 e | 17–49 f |
| Temperature optimum (°C) | 37 | 37 a | 30 b | 35 c | 37 d | 35 e | 37 f |
| Hydrolysis of starch | − | + | − | + | + | + | + |
| Hydrolysis of Tween 80 | − | + | − | + | − | − | − |
| Hydrolysis of casein | − | − | + | − | − | + | − |
| Hydrolysis of aesculin | − | + | + | − | − | + | − |
| Production of indole | − | − | − | − | + | − | − |
| Methyl red test | + | − | − | − | − | − | − |
| Nitrate and nitrite reduction | + | + | + | − | + | + | + |
| Arginine decarboxylase | − | + | + | − | + | + | − |
| Acid production from carbohydrates: | |||||||
| D-fructose | + | + | + | − | + | + | + |
| D-galactose | + | − | + | − | − | − | − |
| Lactose | + | − | + | − | − | − | − |
| Maltose | − | − | + | + | + | + | − |
| Mannitol | − | + | + | + | + | + | − |
| Sucrose | + | + | + | + | + | − | + |
| D-xylose | − | + | + | − | − | − | − |
| Utilization as sole carbon and energy source of the: | |||||||
| Amygdalin | + | − | − | + | + | + | − |
| D-arabinose | − | − | − | + | + | − | − |
| Cellobiose | + | − | − | + | + | + | − |
| D-fructose | − | − | − | − | + | + | − |
| D-glucose | + | − | − | + | + | + | + |
| Maltose | + | − | − | + | + | + | + |
| D-mannose | − | + | + | + | + | + | + |
| Melibiose | − | − | + | − | − | − | − |
| Sucrose | + | − | − | + | + | + | + |
| Starch | + | − | − | + | + | + | − |
| D-trehalose | − | − | − | + | + | − | − |
| D-xylose | + | − | − | + | + | + | − |
| Butanol | − | + | + | − | − | − | − |
| Ethanol | − | − | + | + | + | + | + |
| Glycerol | + | − | − | + | + | + | + |
| Mannitol | − | + | + | + | + | + | + |
| Methanol | − | − | + | − | + | − | + |
| Propranolol | − | − | + | − | + | + | + |
| Sorbitol | − | − | − | + | − | − | − |
| Xylitol | − | − | − | − | + | + | − |
| Benzoate | − | + | − | + | + | + | − |
| Butyrate | − | − | − | + | + | + | + |
| Citrate | + | − | − | + | − | + | − |
| Fumarate | − | − | − | + | + | + | − |
| Hypurate | − | − | − | + | + | + | + |
| Succinate | + | − | − | + | + | − | + |
| Valerate | + | - | - | - | + | + | + |
| Utilization as sole carbon, nitrogen and energy source of: | |||||||
| Alanine | + | - | - | + | + | + | + |
| Arginine | + | - | - | + | + | + | + |
| Aspartic acid | - | - | - | + | + | - | + |
| Cysteine | - | - | - | + | + | - | + |
| Glutamine | - | - | - | + | + | - | - |
| L-phenylalanine | - | - | - | + | + | + | + |
| Ornithine | + | - | - | + | + | + | + |
| Serine | - | - | - | - | + | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galisteo, C.; Sánchez-Porro, C.; de la Haba, R.R.; López-Hermoso, C.; Fernández, A.B.; Farias, M.E.; Ventosa, A. Characterization of Salinivibrio socompensis sp. nov., A New Halophilic Bacterium Isolated from the High-Altitude Hypersaline Lake Socompa, Argentina. Microorganisms 2019, 7, 241. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080241
Galisteo C, Sánchez-Porro C, de la Haba RR, López-Hermoso C, Fernández AB, Farias ME, Ventosa A. Characterization of Salinivibrio socompensis sp. nov., A New Halophilic Bacterium Isolated from the High-Altitude Hypersaline Lake Socompa, Argentina. Microorganisms. 2019; 7(8):241. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080241
Chicago/Turabian StyleGalisteo, Cristina, Cristina Sánchez-Porro, Rafael R. de la Haba, Clara López-Hermoso, Ana B. Fernández, María E. Farias, and Antonio Ventosa. 2019. "Characterization of Salinivibrio socompensis sp. nov., A New Halophilic Bacterium Isolated from the High-Altitude Hypersaline Lake Socompa, Argentina" Microorganisms 7, no. 8: 241. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7080241