Efficiency of Phage φ6 for Biocontrol of Pseudomonas syringae pv. syringae: An in Vitro Preliminary Study

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Preparation of Phage φ6 and Enrichment
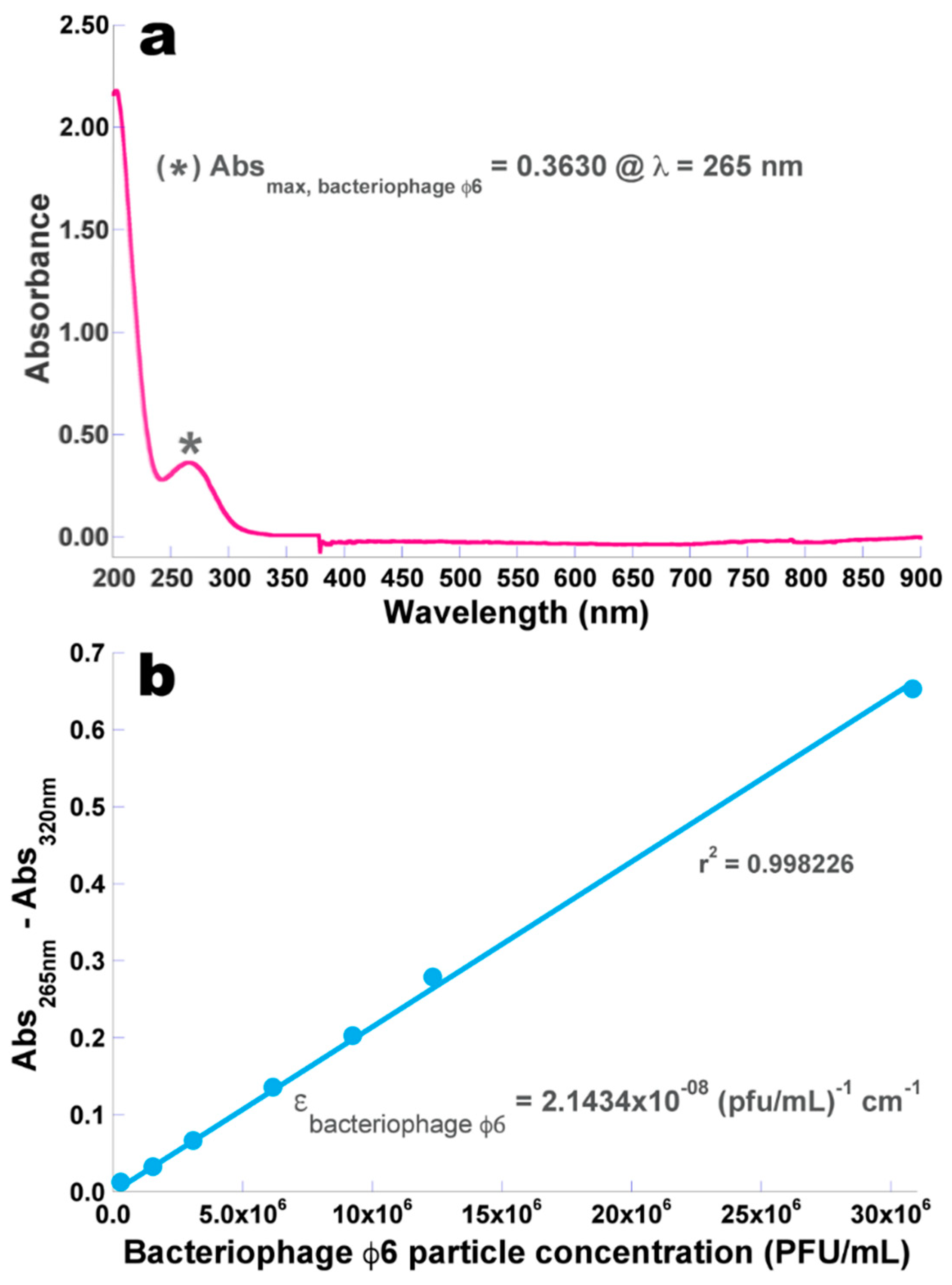
2.3. Determination of the Molar Extinction Coefficient of the Isolated Phage φ6 Particles
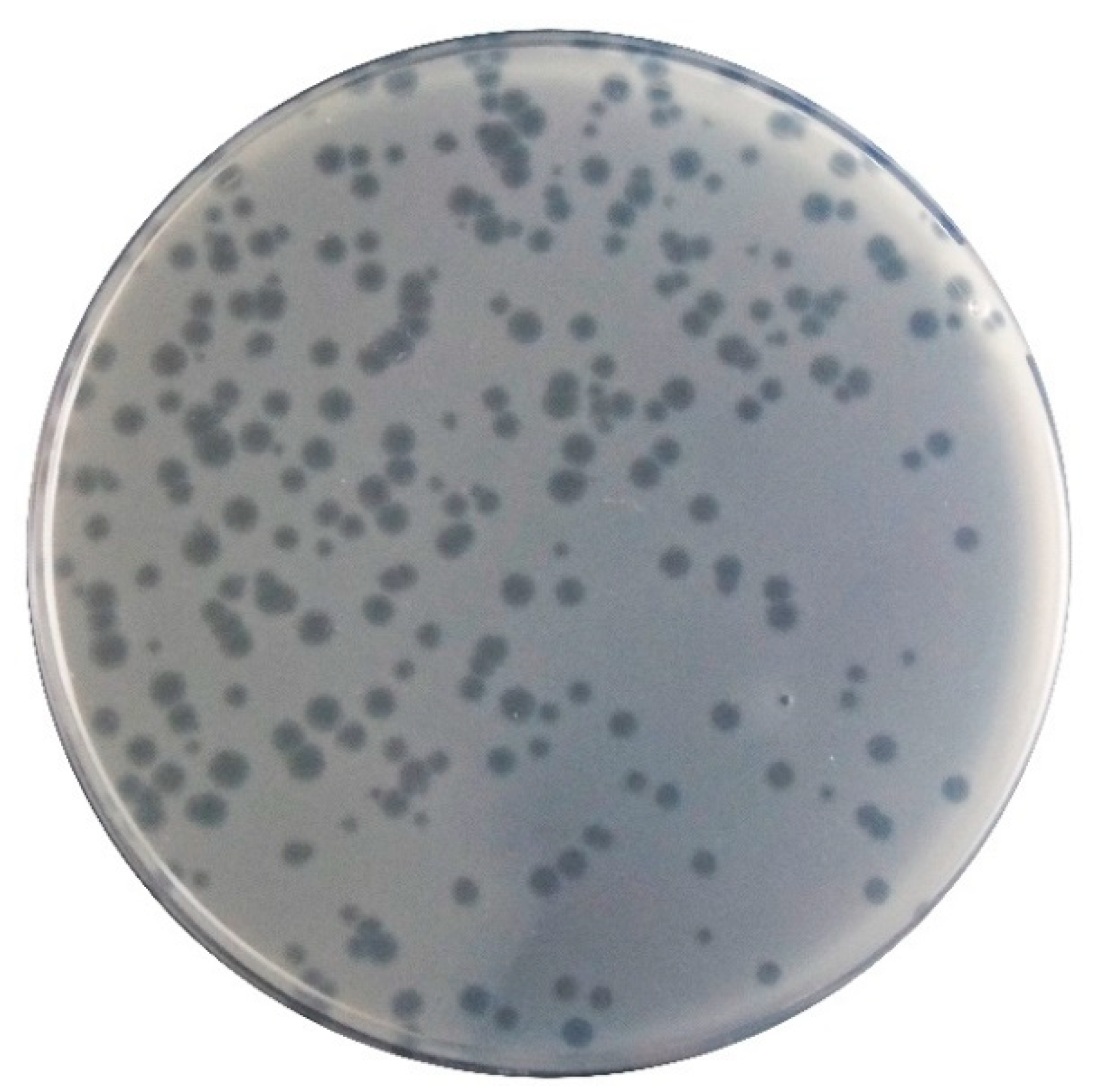
2.4. Phage φ6 Host Range: Spot Test and Efficiency of Plating (EOP) Assays
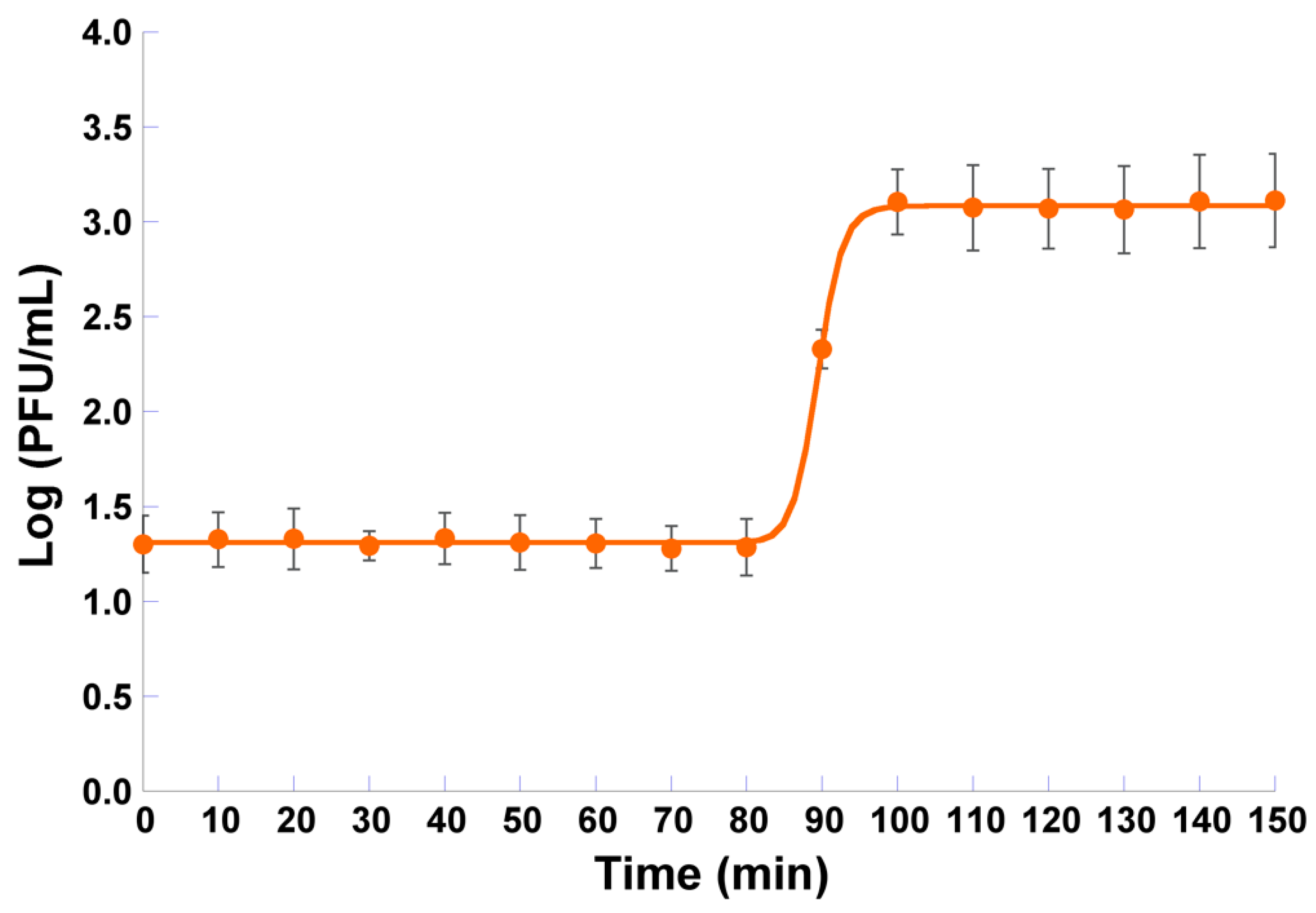
2.5. One-Step Growth Curve
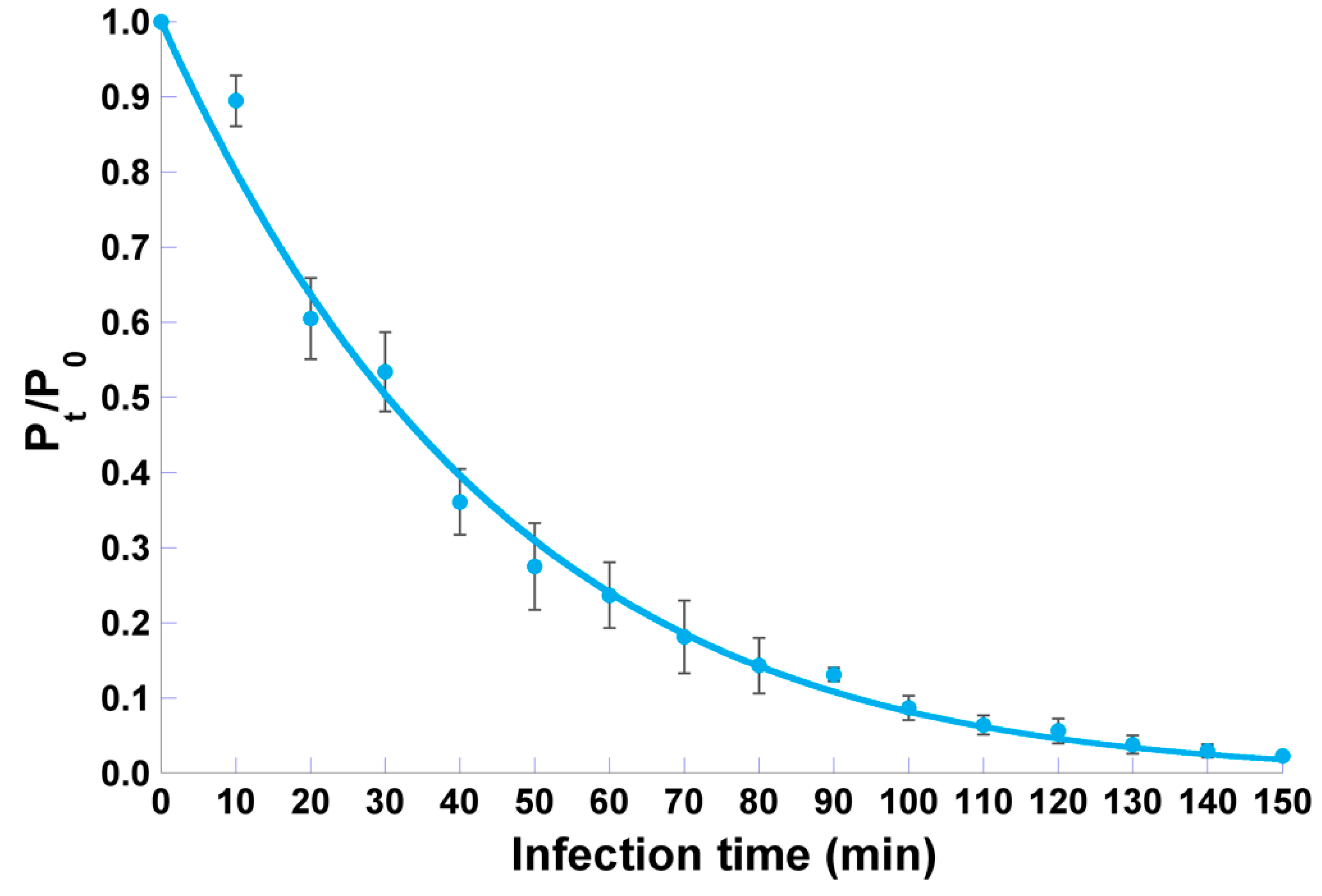
2.6. Adsorption Curve
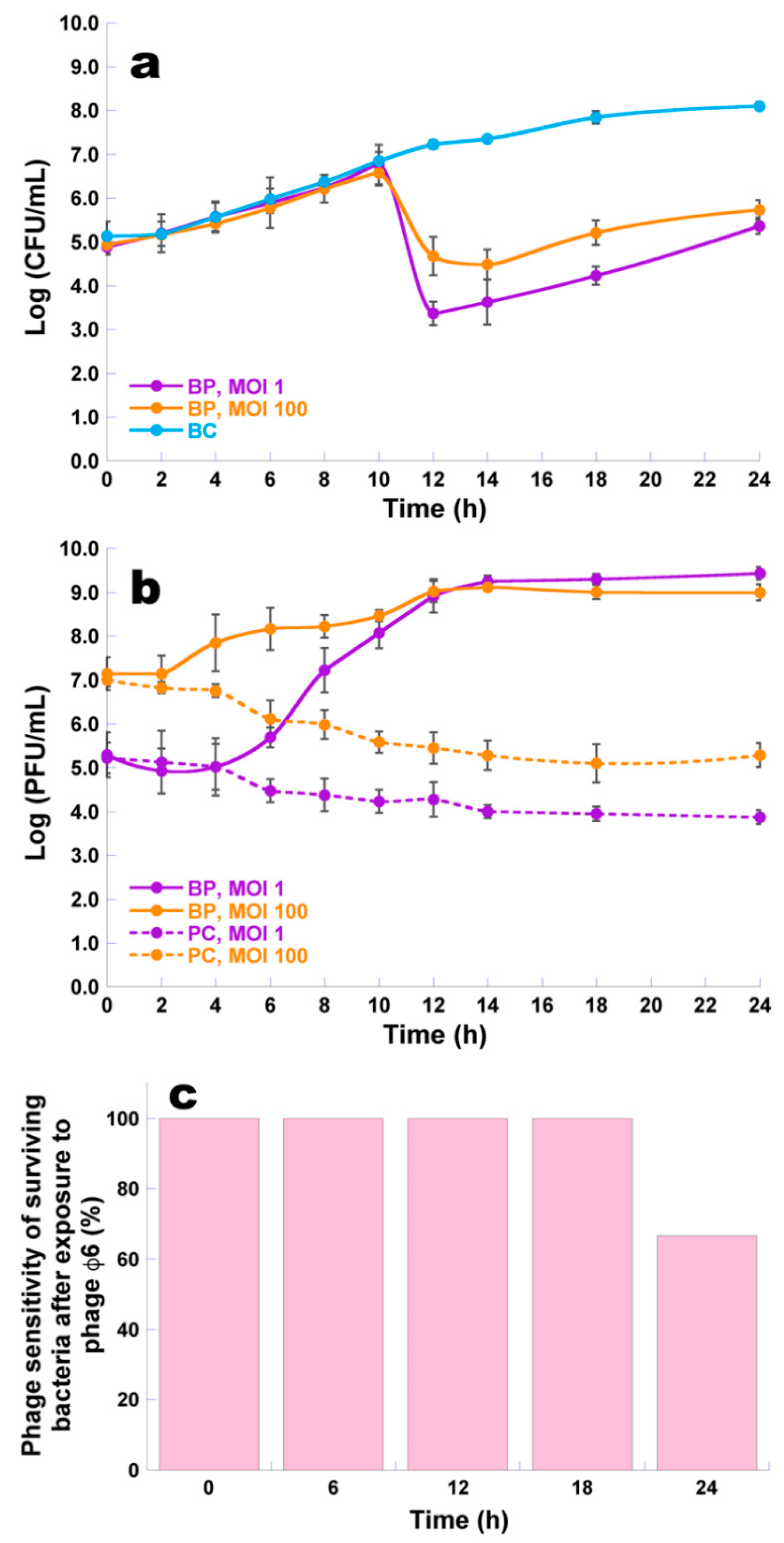
2.7. Bacterial Kill Curves
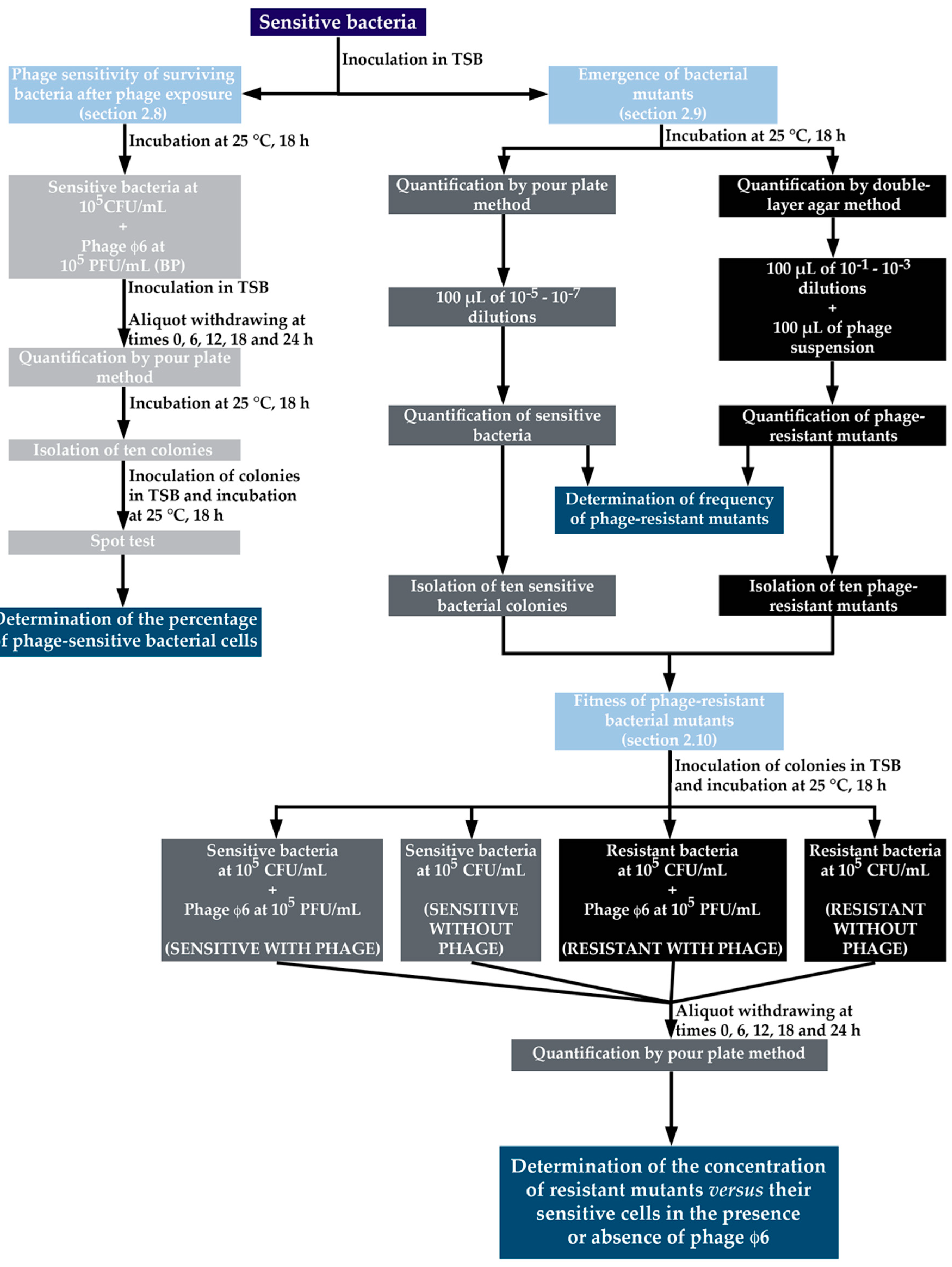
2.8. Phage Sensitivity of Surviving Bacteria after Phage Exposure
2.9. Isolation of Phage-Resistant Mutants and Determination of the Frequency of Emergence of Phage-Resistant Bacterial Mutants
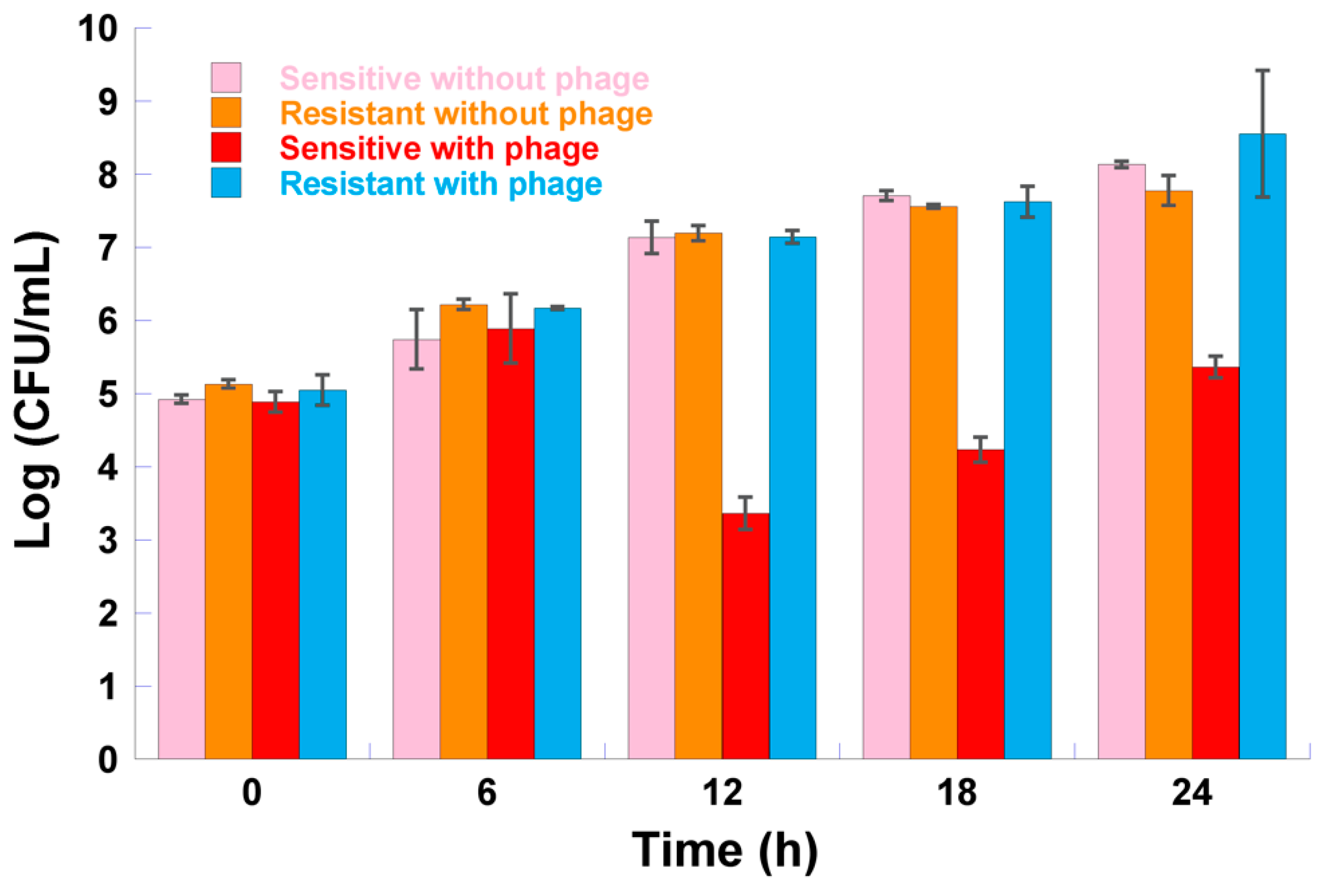
2.10. Fitness of Phage-Resistant Bacterial Mutants
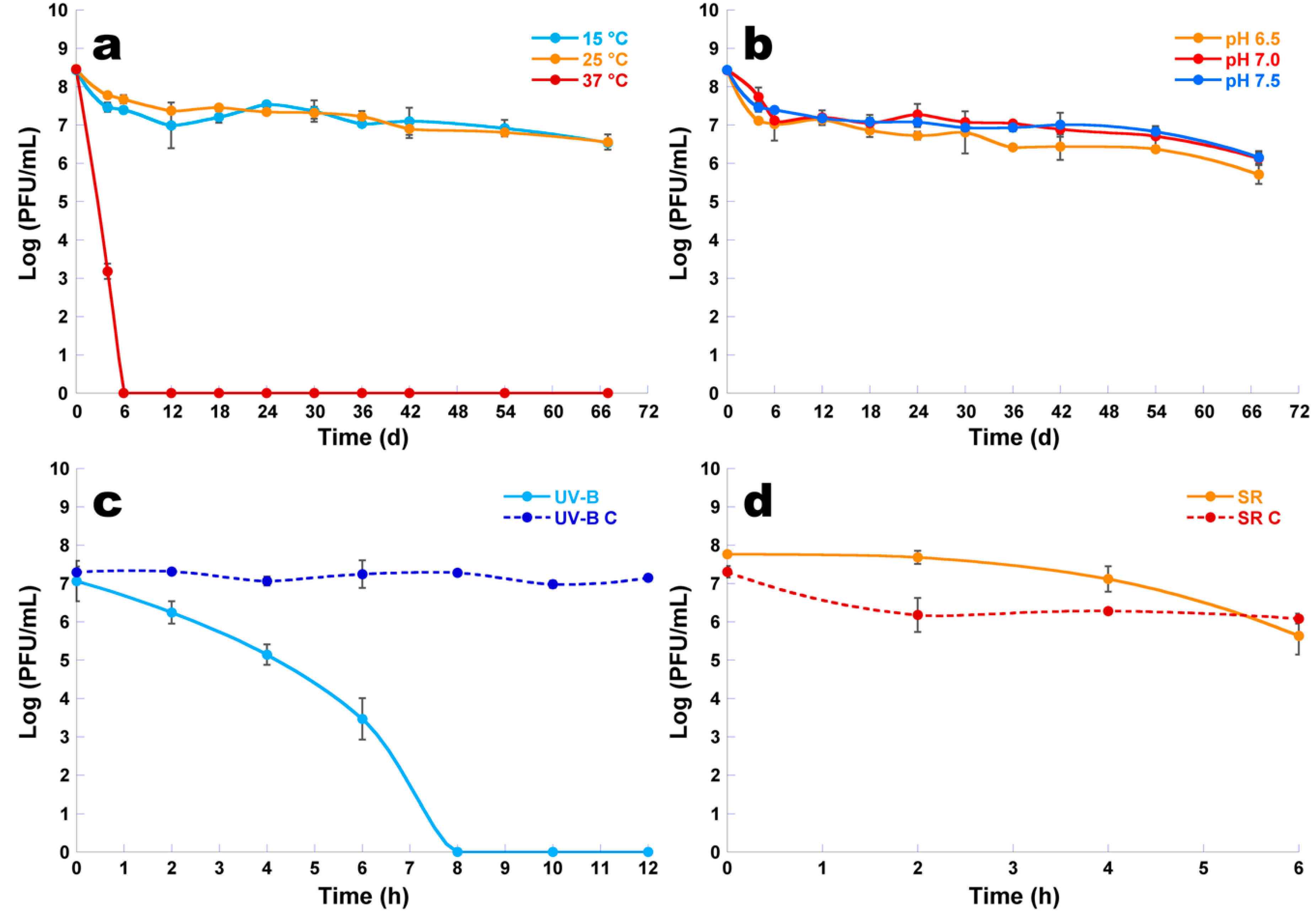
2.11. Assessment of the Effect of Environmental Factors upon Phage φ6 Viability
2.11.1. pH Experiments
2.11.2. Temperature Experiments
2.11.3. UV-B Irradiation Experiments
2.11.4. Solar Radiation Experiments
2.12. Statistical Analyses
3. Results
3.1. Phage Preparation and Enrichment
3.2. Determination of the Molar Extinction Coefficient of the Isolated Phage φ6 Particles
3.3. Phage Host Range and Efficiency of Plating (EOP)
3.4. One-Step Growth Curve Analysis
3.5. Adsorption Curve
3.6. Bacterial Kill Curves and Host Sensitivity to Phage φ6
3.7. Determination of the Frequency of Emergence of Phage-Resistant Bacterial Mutants
3.8. Fitness of Phage-Resistant Mutants
3.9. Assessment of the Effect of Environmental Factors upon Phage φ6 Viability
3.9.1. Temperature Experiments
3.9.2. pH Experiments
3.9.3. UV-B Experiments
3.9.4. Solar Radiation Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bradbury, J.F. Guide to Plant Pathogenic Bacteria; CAB International: Farnham Royal, Slough, 1986; ISBN 0851985572. [Google Scholar]
- Kennelly, M.M.; Cazorla, F.M.; de Vicente, A.; Ramos, C.; Sundin, G.W. Pseudomonas syringae diseases of fruit trees: Progress toward understanding and control. Plant Dis. 2007, 91, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Ivanović, Ž.; Perović, T.; Popović, T.; Blagojević, J.; Trkulja, N.; Hrnčić, S. Characterization of Pseudomonas syringae pv. syringae, causal agent of citrus blast of mandarin in Montenegro. Plant Pathol. J. 2017, 33, 21–33. [Google Scholar] [PubMed]
- Pscheidt, J.W.; Ocamb, C.M. Disease Management Handbook; Oregon State University Extension Service/OSU Extension Catalog; Pacific Northwest (PNW): Oregon State University, Corvallis, OR, USA, 2018. [Google Scholar]
- Altimira, F.; Yanez, C.; Bravo, G.; Gonzalez, M.; Rojas, L.A.; Seeger, M. Characterization of copper-resistant bacteria and bacterial communities from copper-polluted agricultural soils of central Chile. BMC Microbiol. 2012, 12, 193. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Cunha, Â.; Gomes, N.C.M.; Alves, E.; Costa, L.; Faustino, M.A.F. Phage therapy and photodynamic therapy: Low environmental impact approaches to inactivate microorganisms in fish farming plants. Mar. Drugs 2009, 7, 268–313. [Google Scholar] [CrossRef] [PubMed]
- Rios, A.C.; Moutinho, C.G.; Pinto, F.C.; Del Fiol, F.S.; Jozala, A.; Chaud, M.V.; Vila, M.M.D.C.; Teixeira, J.A.; Balcao, V.M. Alternatives to overcoming bacterial resistances: State-of-the-art. Microbiol. Res. 2016, 191, 51–80. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.; Del Fiol, F.S.; Balcão, V.M. Prospects for the use of new technologies to combat multidrug-resistant bacteria. Front. Pharmacol. 2019, 10, 692. [Google Scholar] [CrossRef]
- Vieira, A.; Silva, Y.J.; Cunha, Â.; Gomes, N.C.M.; Ackermann, H.W.; Almeida, A. Phage therapy to control multidrug-resistant Pseudomonas aeruginosa skin infections: In vitro and ex vivo experiments. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 3241–3249. [Google Scholar] [CrossRef]
- Silva, Y.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, A.; Calado, R.; Gomes, N.; Pardo, M.; Hernandez, I.; Almeida, A. Phage Therapy as an approach to prevent Vibrio anguillarum infections in fish larvae production. PLoS ONE 2014, 9, e114197. [Google Scholar] [CrossRef]
- Silva, Y.; Moreirinha, C.; Pereira, C.; Costa, L.; Rocha, R.J.M.; Cunha, Â.; Gomes, N.C.M.; Calado, R.; Almeida, A. Biological control of Aeromonas salmonicida infection in juvenile Senegalese sole (Solea senegalensis) with phage AS-A. Aquaculture 2016, 450, 225–233. [Google Scholar] [CrossRef]
- Pereira, C.; Silva, Y.J.; Santos, A.L.; Cunha, Â.; Gomes, N.C.M.; Almeida, A. Bacteriophages with potential for inactivation of fish pathogenic bacteria: Survival, host specificity and effect on bacterial community structure. Mar. Drugs 2011, 9, 2236–2255. [Google Scholar] [CrossRef]
- Park, S.; Nakai, T. Bacteriophage control of Pseudomonas plecoglossicida infection in ayu. Dis. Aquat. Organ. 2003, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.; Harper, D.; Burch, D.; Anggard, E.; Soothill, J. Topical treatment of Pseudomonas aeruginosa otitis of dogs with a bacteriophage mixture: A before/after clinical trial. Vet. Microbiol. 2010, 146, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.J.; Wu, J.; Lee, H.J.; Jo, E.J.; Murugaiyan, S.; Chung, E.; Lee, S.W. Biocontrol potential of a lytic bacteriophage PE204 against bacterial wilt of tomato. J. Microbiol. Biotechnol. 2012, 22, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.; Kim, M.; Lee, D.; Lee, Y.; Park, J.; Youn, H.; Lee, H.; Yang, S.; Cho, Y.; Lee, J.; et al. Research in veterinary science use of bacteriophage for biological control of Salmonella Enteritidis infection in chicken. Res. Vet. Sci. 2012, 93, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Frampton, R.A.; Pitman, A.R.; Fineran, P.C. Advances in bacteriophage-mediated control of plant pathogens. Int. J. Microbiol. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; Lojkowska, E. Isolation and characterization of novel soilborne lytic bacteriophages infecting Dickeya spp. biovar 3 (‘D. solani’). Plant Pathol. 2014, 63, 758–772. [Google Scholar] [CrossRef]
- Prior, S.E.; Andrews, A.J.; Nordeen, R.O.; Priori, S.E. Characterization of bacteriophages of Pseudomonas syringae pv. tomato. J. Ark. Acad. Sci. 2007, 61, 14. [Google Scholar]
- Vidaver, A.K.; Koski, R.K.; Van Etten, J.L. Bacteriophage phi6: A lipid-containing virus of Pseudomonas phaseolicola. J. Virol. 1973, 11, 799–805. [Google Scholar]
- Mindich, L.; Qiao, X.; Qiao, J.; Onodera, S.; Romantschuk, M.; Hoogstraten, D. Isolation of additional bacteriophages with genomes of segmented double- stranded RNA. J. Bacteriol. 1999, 181, 4505–4508. [Google Scholar]
- Qiao, X.; Sun, Y.; Qiao, J.; Di Sanzo, F.; Mindich, L. Characterization of Φ2954, a newly isolated bacteriophage containing three dsRNA genomic segments. BMC Microbiol. 2010, 10, 55. [Google Scholar] [CrossRef]
- Nordeen, R.O.; Morgan, M.K.; Currier, T.C. Isolation and partial characterization of bacteriophages of the phytopathogen Pseudomonas syringae. Appl. Environ. Microbiol. 1983, 45, 1890–1898. [Google Scholar] [PubMed]
- Smith, A.R.W.; Zamze, S.E.; Hignett, R.C. Morphology and hydrolytic activity of A7, a typing phage of Pseudomonas syringae pv. morsprunorum. Microbiology 1994, 140, 905–913. [Google Scholar] [CrossRef]
- Rombouts, S.; Volckaert, A.; Venneman, S.; Declercq, B.; Vandenheuvel, D.; Allonsius, C.N.; Van Malderghem, C.; Jang, H.B.; Briers, Y.; Noben, J.P.; et al. Characterization of novel bacteriophages for biocontrol of bacterial blight in leek caused by Pseudomonas syringae pv. porri. Front. Microbiol. 2016, 7, 279. [Google Scholar] [CrossRef] [PubMed]
- Di Lallo, G.; Evangelisti, M.; Mancuso, F.; Ferrante, P.; Marcelletti, S.; Tinari, A.; Superti, F.; Migliore, L.; D’Addabbo, P.; Frezza, D.; et al. Isolation and partial characterization of bacteriophages infecting Pseudomonas syringae pv. actinidiae, causal agent of kiwifruit bacterial canker. J. Basic Microbiol. 2014, 54, 1210–1221. [Google Scholar] [PubMed]
- Yu, J.; Lim, J.; Song, Y.; Heu, S.; Kim, G.; Koh, Y.; Oh, C. Isolation and characterization of bacteriophages against Pseudomonas syringae pv. actinidiae causing bacterial canker disease in kiwifruit. J. Microbiol. Biotechnol. 2016, 26, 385–393. [Google Scholar]
- Frampton, R.; Taylor, C.; Holguin Moreno, A.; Visnovsky, S.; Petty, N.; Pitman, A.; Fineran, P. Identification of bacteriophages for biocontrol of the kiwifruit canker phytopathogen Pseudomonas syringae pv. actinidiae. Appl. Environ. Microbiol. 2014, 80, 2216–2228. [Google Scholar] [CrossRef]
- Frampton, R.; Acedo, E.L.; Young, V.L.; Chen, D.; Tong, B.; Taylor, C.; Easingwood, R.A.; Pitman, A.R.; Kleffmann, T.; Bostina, M.; et al. Genome, proteome and structure of a T7-like bacteriophage of the kiwifruit canker phytopathogen Pseudomonas syringae pv. actinidiae. Viruses 2015, 7, 3361–3379. [Google Scholar] [CrossRef]
- Park, J.; Lim, J.A.; Yu, J.G.; Oh, C.S. Genomic features and lytic activity of the bacteriophage PPPL-1 effective against Pseudomonas syringae pv. actinidiae, a cause of bacterial canker in kiwifruit. J. Microbiol. Biotechnol. 2018, 28, 1542–1546. [Google Scholar]
- Yin, Y.; Ni, P.; Deng, B.; Wang, S.; Xu, W.; Wang, D. Isolation and characterisation of phages against Pseudomonas syringae pv. actinidiae. Acta Agric. Scand. Sect. B Soil Plant Sci. 2019, 69, 199–208. [Google Scholar] [CrossRef]
- Mindich, L.; Nemhauser, I.; Gottlieb, P.; Romantschuk, M.; Carton, J.; Frucht, S.; Strassman, J.; Bamford, D.H.; Kalkkinen, N. Nucleotide sequence of the large double-stranded RNA segment of bacteriophage phi 6: Genes specifying the viral replicase and transcriptase. J. Virol. 1988, 62, 1180–1185. [Google Scholar]
- Wickner, R.B. Double-stranded RNA virus replication and packaging. J. Biol. Chem. 1993, 268, 3797–3800. [Google Scholar] [PubMed]
- Wei, H.; Cheng, R.H.; Berriman, J.; Rice, W.J.; Stokes, D.L.; Katz, A.; Morgan, D.G.; Gottlieb, P. Three-dimensional structure of the enveloped bacteriophage Φ12: An incomplete T = 13 lattice is superposed on an enclosed T = 1 shell. PLoS ONE 2009, 4, e6850. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, S.; Shen, W.; Zhao, X.; Shen, M.; Tan, Y.; Li, G.; Li, M.; Wang, J.; Hu, F.; et al. Characterization of the first double-stranded RNA bacteriophage infecting Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 38795. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dennehy, J.J. Differential bacteriophage mortality on exposure to copper. Appl. Environ. Microbiol. 2011, 77, 6878–6883. [Google Scholar] [CrossRef] [PubMed]
- Louvado, A.; Coelho, F.J.R.C.; Domingues, P.; Santos, A.L.; Gomes, N.C.M.; Almeida, A.; Cunha, Â. Isolation of surfactant-resistant pseudomonads from the estuarine surface microlayer. J. Microbiol. Biotechnol. 2012, 22, 283–291. [Google Scholar] [CrossRef]
- Oliveira, V.; Gomes, N.C.M.; Almeida, A.; Silva, A.M.S.; Simões, M.M.Q.; Smalla, K.; Cunha, Â. Hydrocarbon contamination and plant species determine the phylogenetic and functional diversity of endophytic degrading bacteria. Mol. Ecol. 2014, 23, 1392–1404. [Google Scholar] [CrossRef]
- Callanan, J.; Stockdale, S.R.; Shkoporov, A.; Draper, L.A.; Ross, R.P.; Hill, C. RNA phage biology in a metagenomic era. Viruses 2018, 10, 386. [Google Scholar] [CrossRef]
- Mäntynen, S.; Sundberg, L.R.; Poranen, M.M. Recognition of six additional cystoviruses: Pseudomonas virus phi6 is no longer the sole species of the family Cystoviridae. Arch. Virol. 2018, 163, 1117–1124. [Google Scholar] [CrossRef]
- Adams, M.H. Bacteriophages; Interscience Publishers, Inc.: New York, NY, USA, 1959. [Google Scholar]
- Rios, A.; Vila, M.; Lima, R.; Del Fiol, F.; Tubino, M.; Teixeira, J.; Balcão, V. Structural and functional stabilization of bacteriophage particles within the aqueous core of a W/O/W multiple emulsion: A potential biotherapeutic system for the inhalational treatment of bacterial pneumonia. Process Biochem. 2018, 64, 177–192. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Cunha, Â.; Delgadillo, I.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Bacteriophages with potential to inactivate Salmonella Typhimurium: Use of single phage suspensions and phage cocktails. Virus Res. 2016, 220, 179–192. [Google Scholar] [CrossRef]
- Kutter, E. Phage host range and efficiency of plating. In Bacteriophage: Methods and Protocols; Clokie, M.R.K.A.M., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 141–149. ISBN 978-1-58829-682-5. [Google Scholar]
- Melo, D.R.; Sillankorva, S.; Ackermann, H.; Kropinski, A.M.; Azeredo, J.; Cerca, N. Isolation and characterization of a new Staphylococcus epidermidis broad-spectrum bacteriophage. J. Gen. Virol. 2014, 95, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Mateus, L.; Costa, L.; Silva, Y.J.; Pereira, C.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424–425, 167–173. [Google Scholar] [CrossRef]
- Stuer-Lauridsen, B.; Janzen, T.; Schnabl, J.; Johansen, E. Identification of the host determinant of two prolate-headed phages infecting Lactococcus lactis. Virology 2003, 309, 10–17. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, I.N. Bacteriophage adsorption rate and optimal lysis time. Genetics 2008, 180, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.B.; Carvalho, C.; Azeredo, J.; Ferreira, E.C. Population dynamics of a Salmonella lytic phage and its host: Implications of the host bacterial growth rate in modelling. PLoS ONE 2014, 9, e102507. [Google Scholar] [CrossRef]
- García, R.; Latz, S.; Romero, J.; Higuera, G.; García, K.; Bastías, R. Bacteriophage production models: An overview. Front. Microbiol. 2019, 10, 1–7. [Google Scholar] [CrossRef]
- Filippov, A.; Sergueev, K.V.; He, Y.; Huang, X.Z.; Gnade, B.T.; Mueller, A.J.; Fernandez-Prada, C.; Nikolich, M.P. Bacteriophage-resistant mutants in Yersinia pestis: Identification of phage receptors and attenuation for mice. PLoS ONE 2011, 6, e25486. [Google Scholar] [CrossRef]
- Moore, L.W. Pseudomonas syringae: Disease and ice nucleation activity. In Ornamentals Northwest Archives; Department of Botany and Plant Pathology, Oregon State University: Corvallis, OR, USA, 1988; Volume 12, pp. 3–16. [Google Scholar]
- Sistrom, M.; Park, D.; O’Brien, H.E.; Wang, Z.; Guttman, D.S.; Townsend, J.P.; Turner, P.E. Genomic and gene-expression comparisons among phage-resistant type-IV pilus mutants of Pseudomonas syringae pathovar phaseolicola. PLoS ONE 2015, 10, e0144514. [Google Scholar] [CrossRef]
- McGrane, R.; Beattie, G.A. Pseudomonas syringae pv. syringae regulates multiple stages of plant colonization via the bacteriophytochrome BphP1. MBio 2017, 8, e01178-17. [Google Scholar]
- He, R.; Liu, P.; Jia, B.; Xue, S.; Wang, X.; Hu, J.; Al Shoffe, Y.; Gallipoli, L.; Mazzaglia, A.; Balestra, G.; et al. Genetic diversity of Pseudomonas syringae pv. actinidiae strains from different geographic Regions in China. Phytopathology 2019, 109, 347–357. [Google Scholar]
- Vasebi, Y.; Khakvar, R.; Faghihi, M.M.; Vinatzer, A. Genomic and pathogenic properties of Pseudomonas syringae pv. syringae strains isolated from apricot in East Azerbaijan province. Iran. Biocatal. Agric. Biotechnol. 2019, 19, 101167. [Google Scholar]
- Park, S.C.; Nakai, T. infection in ayu Plecoglossus altivelis. Dis. Aquat. Organ. 2003, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Characterization and in vitro evaluation of new bacteriophages for the biocontrol of Escherichia coli. Virus Res. 2017, 227, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Pereira, C.; Santos, L.; Klumpp, J.; Almeida, A. Potential of phage cocktails in the inactivation of Enterobacter cloacae-An in vitro study in a buffer solution and in urine samples. Virus Res. 2016, 211, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, T.; Sawada, H. Genome analysis of the kiwifruit canker pathogen Pseudomonas syringae pv. actinidiae biovar 5. Sci. Rep. 2016, 6, 21399. [Google Scholar] [CrossRef] [PubMed]
- McCann, H.C.; Li, L.; Liu, Y.; Li, D.; Pan, H.; Zhong, C.; Rikkerink, E.H.A.; Templeton, M.D.; Straub, C.; Colombi, E.; et al. Origin and evolution of the kiwifruit canker pandemic. Genome Biol. Evol. 2017, 9, 932–944. [Google Scholar] [CrossRef]
- Ferris, M.T.; Joyce, P.; Burch, C.L. High frequency of mutations that expand the host range of an RNA virus. Genet. Soc. Am. 2007, 1022, 1013–1022. [Google Scholar] [CrossRef]
- Simpson, D.J. The Discovery and Application of Bacteriophage Receptor Binding Proteins. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2016. [Google Scholar]
- Moldovan, R.; Chapman-McQuiston, E.; Wu, X.L. On kinetics of phage adsorption. Biophys. J. 2007, 93, 303–315. [Google Scholar] [CrossRef]
- Ceyssens, P. Isolation and Characterization of Lytic Bacteriophages Infecting Pseudomonas aeruginosa. Ph.D. Thesis, Katholieke Inuversiteit Leuven, Leuven, Belgium, 2009. [Google Scholar]
- Hyman, P.; Abedon, S.T. Practical methods for determining phage growth parameters. In Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Humana Press: Totowa, NJ, USA, 2009; Volume 501, ISBN 9781603271646. [Google Scholar]
- Storms, Z.J.; Sauvageau, D. Modeling tailed bacteriophage adsorption: Insight into mechanisms. Virology 2015, 485, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, H.; McKean, K.; Wang, I. Phage fitness may help predict phage therapy efficacy. Bacteriophage 2014, 4, e964081. [Google Scholar] [CrossRef]
- ChiHsin, H.; ChongYi, L.; JongKang, L.; ChanShing, L. Control of the eel (Anguilla japonica) pathogens, Aeromonas hydrophila and Edwardsiella tarda, by bacteriophages. J. Fish. Soc. Taiwan 2000, 27, 21–31. [Google Scholar]
- Pasharawipas, T.; Manopvisetcharean, J.; Flegel, T. Phage treatment of Vibrio harveyi: A general concept of protection against bacterial infection. Res. J. Microbiol. 2011, 6, 560–567. [Google Scholar] [CrossRef]
- Prasad, Y.; Kumar, D.; Sharma, A. Lytic bacteriophages specific to Flavobacterium columnare rescue catfish, Clarias batrachus (Linn.) from columnaris disease. J. Environ. Biol. 2011, 32, 161–168. [Google Scholar] [PubMed]
- Nakai, T. Application of bacteriophages for control of infectious diseases in aquaculture. In Bacteriophages in the Control of Food and Waterborne Pathogens; Sabour, P., Griffiths, M., Eds.; ASM Press: Washington, DC, USA, 2010; pp. 257–272. [Google Scholar]
- Silva, Y.; Costa, L.; Pereira, C.; Cunha, A.; Calado, R.; Gomes, N.; Almeida, A. Influence of environmental variables in the efficiency of phage therapy in aquaculture. Microb. Biotechnol. 2014, 7, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.; Pereira, C.; Moreirinha, C.; Salvio, R.; Lopes, A.; Wang, D.; Almeida, A. New insights on phage efficacy to control Aeromonas salmonicida in aquaculture systems: An in vitro preliminary study. Aquaculture 2018, 495, 970–982. [Google Scholar] [CrossRef]
- Arisaka, F.; Kanamaru, S.; Leiman, P.; Rossmann, M.G. The tail lysozyme complex of bacteriophage T4. Int. J. Biochem. Cell Biol. 2003, 35, 16–21. [Google Scholar] [CrossRef]
- Abedon, S.T. Lysis from without. Bacteriophage 2011, 1, 46–49. [Google Scholar] [CrossRef]
- Lopes, A.; Pereira, C.; Almeida, A. Sequential combined effect of phages and antibiotics on the inactivation of E. coli. Microorganisms 2018, 6, 125. [Google Scholar] [CrossRef]
- Levin, B.R.; Bull, J.J. Population and evolutionary dynamics of phage therapy. Nat. Rev. Microbiol. 2004, 2, 20–24. [Google Scholar] [CrossRef]
- Gill, J.J.; Hyman, P. Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef]
- Wright, R.C.; Friman, V.P.; Smith, M.C.M.; Brockhurst, M.A. Cross-resistance is modular in bacteria–phage interactions. PLoS Biol. 2018, 16, e2006057. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Shen, M.; Yang, Y.; Le, S.; Li, M.; Wang, J.; Zhao, Y.; Tan, Y.; Hu, F.; Lu, S. Adaptation of Pseudomonas aeruginosa to phage PaP1 predation via O-antigen polymerase mutation. Front. Microbiol. 2018, 9, 1170. [Google Scholar] [CrossRef] [PubMed]
- Bohannan, B.J.M.; Travisano, M.; Lenski, R.E. Epistatic interactions can lower the cost of resistance to multiple consumers. Evolution 1999, 53, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Brockhurst, M.A.; Buckling, A.; Rainey, P.B. The effect of a bacteriophage on diversification of the opportunistic bacterial pathogen, Pseudomonas aeruginosa. Proc. Biol. Sci. 2005, 272, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Lennon, J.T.; Khatana, S.A.M.; Marston, M.F.; Martiny, J.B.H. Is there a cost of virus resistance in marine cyanobacteria? ISME J. 2007, 1, 300–312. [Google Scholar] [CrossRef]
- Quance, M.A.; Travisano, M. Effects of temperature on the fitness cost of resistance to bacteriophage T4 in Escherichia coli. Evolution 2009, 63, 1406–1416. [Google Scholar] [CrossRef]
- Meaden, S.; Paszkiewicz, K.; Koskella, B. The cost of phage resistance in a plant pathogenic bacterium is context-dependent. Evolution 2015, 69, 1321–1328. [Google Scholar] [CrossRef]
- Sandeep, K. Bacteriophage precision drug against bacterial infections. Curr. Sci. 2006, 90, 631–633. [Google Scholar]
- Scott, A.E.; Timms, A.R.; Connerton, P.L.; Carrillo, C.L.; Radzum, K.A.; Connerton, I.F. Genome dynamics of Campylobacter jejuni in response to bacteriophage predation. PLoS Pathog. 2007, 3, 1142–1151. [Google Scholar] [CrossRef]
- Hooton, S.P.T.; Atterbury, R.J.; Connerton, I.F. Application of a bacteriophage cocktail to reduce Salmonella Typhimurium U288 contamination on pig skin. Int. J. Food Microbiol. 2011, 151, 157–163. [Google Scholar] [CrossRef]
- Pereira, C.; Moreirinha, C.; Rocha, R.J.M.; Calado, R.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Application of bacteriophages during depuration reduces the load of Salmonella Typhimurium in cockles. Food Res. Int. 2016, 90, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Moreirinha, C.; Teles, L.; Rocha, R.J.M.; Calado, R.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Application of phage therapy during bivalve depuration improves Escherichia coli decontamination. Food Microbiol. 2017, 61, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Pereira, C.; Gomes, A.; Almeida, A. Efficiency of single phage suspensions and phage cocktail in the inactivation of Escherichia coli and Salmonella Typhimurium: An in vitro preliminary study. Microorganisms 2019, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages—Review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Pirisi, A. Phage therapy—advantages over antibiotics? Lancet 2000, 356, 1418. [Google Scholar] [CrossRef]
- Leverentz, B.; Conway, W.S.; Alavidze, Z.; Janisiewicz, W.J.; Fuchs, Y.; Camp, M.J.; Chighladze, E.; Sulakvelidze, A. Examination of bacteriophage as a biocontrol method for Salmonella on fresh-cut fruit: A model study. J. Food Prot. 2001, 64, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Leverentz, B.; Conway, W.S.; Janisiewicz, W.; Camp, M.J. Optimizing concentration and timing of a phage spray application to reduce Listeria monocytogenes on honeydew melon tissue. J. Food Prot. 2004, 67, 1682–1686. [Google Scholar] [CrossRef] [PubMed]
- Nakai, T.; Sugimoto, R.; Park, K.H.; Matsuoka, S.; Mori, K.; Nishioka, T.; Maruyama, K. Protective effects of bacteriophage on experimental Lactococcus garvieae infection in yellowtail. Dis. Aquat. Organ. 1999, 37, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Endersen, L.; Buttimer, C.; Nevin, E.; Coffey, A.; Neve, H.; Oliveira, H.; Lavigne, R.; O’Mahony, J. Investigating the biocontrol and anti-biofilm potential of a three phage cocktail against Cronobacter sakazakii in different brands of infant formula. Int. J. Food Microbiol. 2017, 253, 1–11. [Google Scholar] [CrossRef]
- Cornelissen, J.; Sibma, F.; Van Logtestijn, R.; Broekman, R.; Thompson, K. Leaf pH as a plant trait: Species-driven rather than soil-driven variation. Funct. Ecol. 2011, 25, 449–455. [Google Scholar] [CrossRef]
- Masoero, G.; Cugnetto, A. The raw pH in plants: A multifaceted parameter. J. Agron. Res. 2018, 1, 18. [Google Scholar] [CrossRef]
- Nasser, A.M.; Oman, S.D. Quantitative assessment of the inactivation of pathogenic and indicator viruses in natural water sources. Water Res. 1999, 33, 1748–1752. [Google Scholar] [CrossRef]
- Olson, M.R.; Axler, R.P.; Hicks, R.E. Effects of freezing and storage temperature on MS2 viability. J. Virol. Methods 2004, 122, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Tey, B.T.; Ooi, S.T.; Yong, K.C.; Yeen, M.; Ng, T.; Ling, T.C.; Siang Tan, W. Production of fusion m13 phage bearing the di-sulphide constrained peptide sequence (C-WSFFSNI-C) that interacts with hepatitis B core antigen. Afr. J. Biotechnol. 2009, 8, 268–273. [Google Scholar]
- Fox, A. Renewed fears as Psa devastates european orchards, 2011. Available online: http://www.stuff.co.nz/business/farming/5050782/Renewed-fears-as-PSA-devastates-European-orchards (accessed on 1 May 2019).
- CABI. Pseudomonas syringae pv. actinidiae (bacterial canker of kiwifruit), 2015. Available online: https://www.cabi.org/isc/datasheet/45002 (accessed on 1 May 2019).
- Das, Q.; Islam, M.R.; Marcone, M.F.; Warriner, K.; Diarra, M.S. Potential of berry extracts to control foodborne pathogens. Food Control 2017, 73, 650–662. [Google Scholar] [CrossRef]
- Kiwifruit Vine Health. Psa-V Seasonal management wall chart 2018-19. Available online: https://www.kvh.org.nz/vdb/document/99346 (accessed on 1 May 2019).
- Wommack, K.E.; Hill, R.T.; Muller, T.A.; Colwell, R.R. Effects of sunlight on bacteriophage viability and structure. Appl. Environ. Microbiol. 1996, 62, 1336–1341. [Google Scholar] [PubMed]
- Lytle, C.D.; Sagripanti, J.L. Predicted inactivation of viruses of relevance to biodefense by solar radiation. J. Virol. 2005, 79, 14244–14252. [Google Scholar] [CrossRef] [PubMed]
- Mojica, K.D.A.; Brussaard, C.P.D. Factors affecting virus dynamics and microbial host-virus interactions in marine environments. FEMS Microbiol. Ecol. 2014, 89, 495–515. [Google Scholar] [CrossRef] [Green Version]
- Hotze, E.M.; Badireddy, A.R.; Chellam, S.; Wiesner, M.R. Mechanisms of bacteriophage inactivation via singlet oxygen generation in UV illuminated fullerol suspensions. Environ. Sci. Technol. 2009, 43, 6639–6645. [Google Scholar] [CrossRef]
- Rule Wigginton, K.; Menin, L.; Montoya, J.P.; Kohn, T. Oxidation of virus proteins during UV (254) and singlet oxygen mediated inactivation. Environ. Sci. Technol. 2010, 44, 5437–5443. [Google Scholar] [CrossRef]
- Balcao, V.M.; Barreira, S.V.P.; Nunes, T.M.; Chaud, M.V.; Tubino, M.; Vila, M.M.D.C. Carbohydrate hydrogels with stabilized phage particles for bacterial biosensing: Bacterium diffusion studies. Appl. Biochem. Biotechnol. 2014, 172, 1194–1214. [Google Scholar] [CrossRef] [PubMed]
- Balcão, V.M.; Glasser, C.A.; Chaud, M.V.; del Fiol, F.S.; Tubino, M.; Vila, M.M.D.C. Biomimetic aqueous-core lipid nanoballoons integrating a multiple emulsion formulation: A suitable housing system for viable lytic bacteriophages. Colloids Surfaces B Biointerfaces 2014, 123, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Balcao, V.M.; Vila, M.M.D.C. Structural and functional stabilization of protein entities: State-of-the-art. Adv. Drug Deliv. Rev. 2015, 93, 25–41. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Spot Test | EOP (%) |
|---|---|---|
| Pseudomonas syringae pv. syringae DSM 21482 | + | 100 (host) |
| Pseudomonas syringae pv. actinidiae CRA-FRU 8.43 | − | 0 |
| Pseudomonas syringae pv. actinidiae CRA-FRU 12.54 | + | 101.3 |
| Pseudomonas syringae pv. actinidiae CRA-FRU 14.10 | + | 96.8 |
| Pseudomonas aeruginosa ATCC 27853 | − | 0 |
| Pseudomonas aeruginosa | − | 0 |
| Pseudomonas gingeri | − | 0 |
| Pseudomonas putida JQ619028 | − | 0 |
| Pseudomonas putida JQ824856 | − | 0 |
| Pseudomonas sp. JX047434 | − | 0 |
| Pseudomonas sp. EF627998 | − | 0 |
| Pseudomonas sp. AF411853 | − | 0 |
| Pseudomonas sp. HF679142 | − | 0 |
| Pseudomonas sp. AB772943 | − | 0 |
| Pseudomonas sp. EU306338 | − | 0 |
| Pseudomonas sp. AY332207 | − | 0 |
| Pseudomonas sp. JN033360 | − | 0 |
| Pseudomonas stutzeri EU167940 | − | 0 |
| Pseudomonas rhodesiae JX994152 | − | 0 |
| Escherichia coli ATCC 13706 | − | 0 |
| Escherichia coli ATCC 25922 | − | 0 |
| Salmonella Typhimurium ATCC 13311 | − | 0 |
| Salmonella Typhimurium ATCC 14028 | − | 0 |
| Aeromonas hydrophila ATCC 7966 | − | 0 |
| Vibrio parahaemolyticus DSM 27657 | − | 0 |
| Sample Volume of Concentrated Phage Suspension (µL) | Final Volume of Dilution (µL) | Number of Phage Particles in the Sample Volume of Concentrated Phage Suspension | Phage Particle Concentration (PFU/mL) | Abs265 nm | Abs320 nm | Abs265 nm–Abs320 nm |
|---|---|---|---|---|---|---|
| 5 | 3000 | 9.2500 × 105 | 3.0833 × 105 | 0.0130 | 0.0000 | 0.0130 |
| 10 | 3000 | 1.8500 × 106 | 6.1666 × 105 | 0.0110 | 0.0110 | 0.0000 |
| 25 | 3000 | 4.6250 × 106 | 1.5416 × 106 | 0.0340 | 0.0010 | 0.0330 |
| 50 | 3000 | 9.2500 × 106 | 3.0833 × 106 | 0.0710 | 0.0040 | 0.0670 |
| 100 | 3000 | 1.8500 × 107 | 6.1666 × 106 | 0.1510 | 0.0150 | 0.1360 |
| 150 | 3000 | 2.7750 × 107 | 9.2500 × 106 | 0.2300 | 0.0270 | 0.2030 |
| 200 | 3000 | 3.7000 × 107 | 1.2333 × 107 | 0.3180 | 0.0390 | 0.2790 |
| 500 | 3000 | 9.2500 × 107 | 3.0833 × 107 | 0.7640 | 0.1110 | 0.6530 |
| Control Sample (CFU/mL) | Sample Treated with Phage φ6 (CFU/mL) | Frequency of Phage-Resistant Bacterial Mutants |
|---|---|---|
| (1.47 ± 0.19) × 108 | (1.75 ± 0.12) × 105 | (1.20 ± 0.62) × 10−3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, L.A.M.; Pereira, C.; Frazão, C.; Balcão, V.M.; Almeida, A. Efficiency of Phage φ6 for Biocontrol of Pseudomonas syringae pv. syringae: An in Vitro Preliminary Study. Microorganisms 2019, 7, 286. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090286
Pinheiro LAM, Pereira C, Frazão C, Balcão VM, Almeida A. Efficiency of Phage φ6 for Biocontrol of Pseudomonas syringae pv. syringae: An in Vitro Preliminary Study. Microorganisms. 2019; 7(9):286. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090286
Chicago/Turabian StylePinheiro, Larindja A. M., Carla Pereira, Carolina Frazão, Victor M. Balcão, and Adelaide Almeida. 2019. "Efficiency of Phage φ6 for Biocontrol of Pseudomonas syringae pv. syringae: An in Vitro Preliminary Study" Microorganisms 7, no. 9: 286. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090286