Accelerating the Biodegradation of High-Density Polyethylene (HDPE) Using Bjerkandera adusta TBB-03 and Lignocellulose Substrates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Fungi
2.2. Preparation of HDPE
2.3. Enzyme Production and Measurement of Enzyme Activity
2.4. Degradation Assays
2.5. Raman Spectroscopy
2.6. Scanning Electron Microscopy Observation
2.7. Statistical Tests
3. Results and Discussion
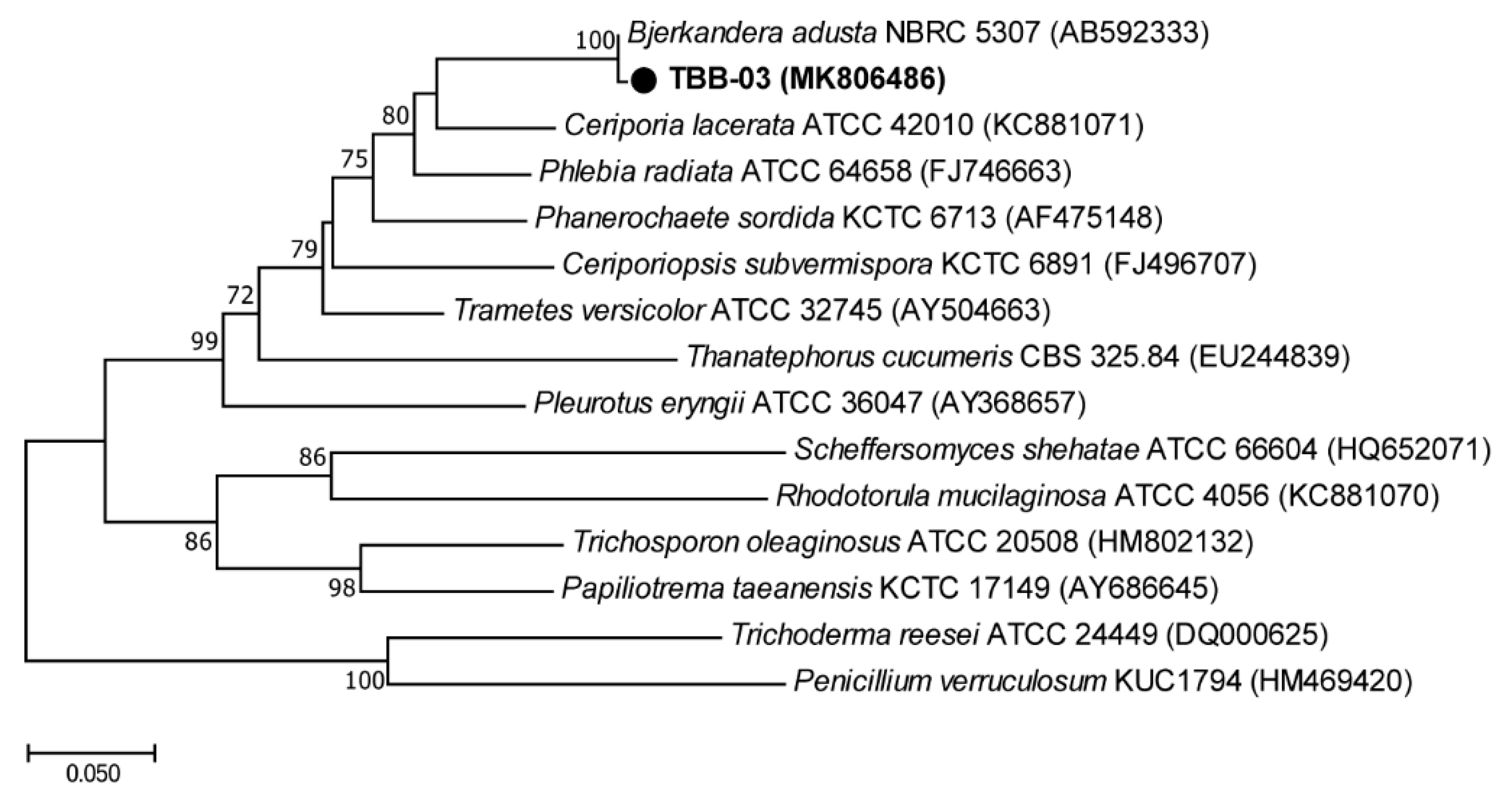
3.1. Isolation and Identification of Laccase-Producing Fungal Strains
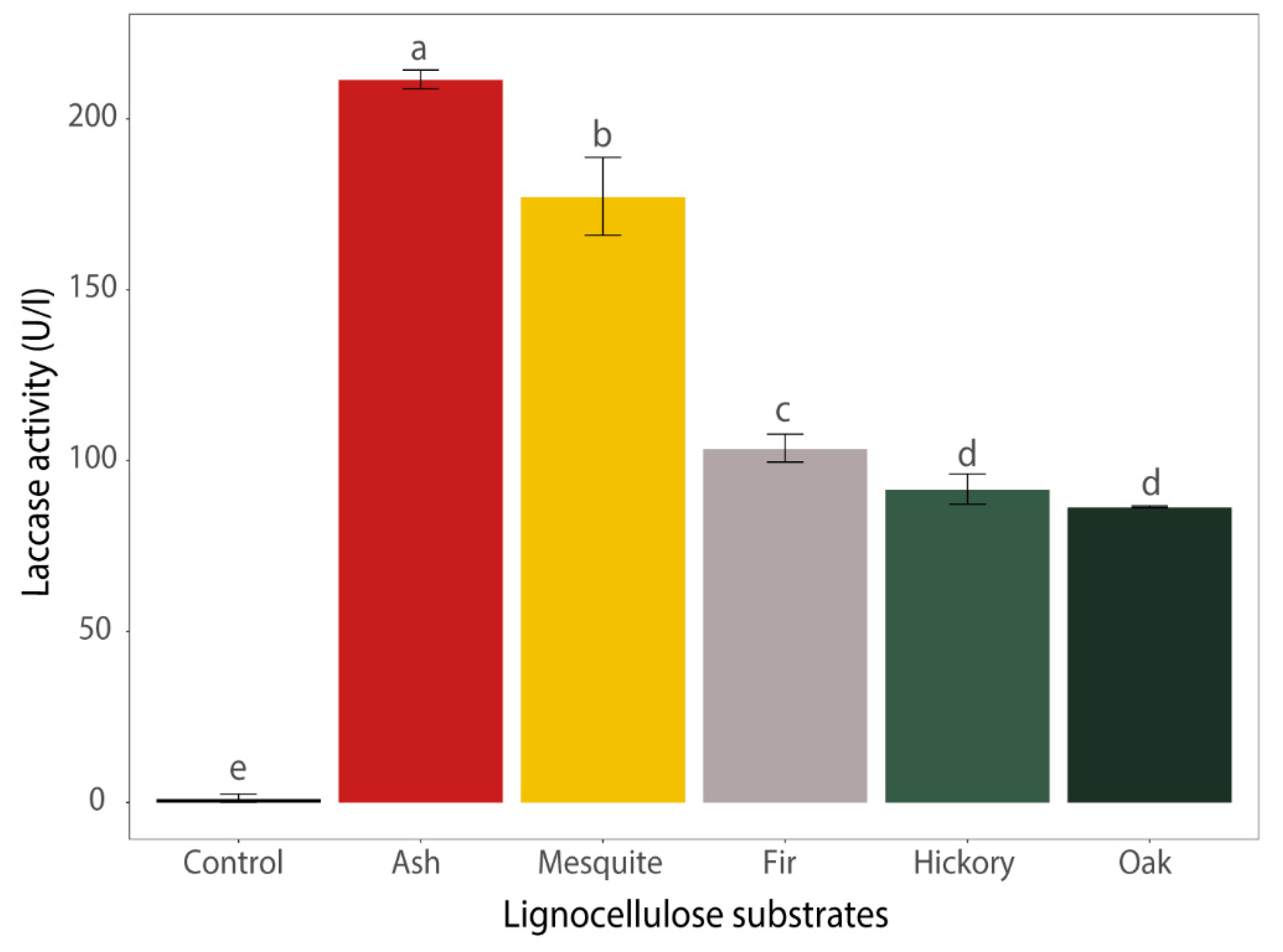
3.2. Enzyme Production During the Cultivation on Media Containing Different Lignocellulose Substrates
3.3. Chemical, Structural, and Morphological Changes of HDPE
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological degradation of plastics: a comprehensive review. Biotechnol. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef] [PubMed]
- Statistical Research Institute. Korean Social Trends; 2018. Available online: http://sri.kostat.go.kr (accessed on 12 December 2018).
- Shimao, M. Biodegradation of plastics. Curr. Opin. Biotechnol. 2001, 12, 242–247. [Google Scholar] [CrossRef]
- Zhang, X.M.; Elkoun, S.; Ajji, A.; Huneault, M.A. Oriented structure and anisotropy properties of polymer blown films: HDPE, LLDPE and LDPE. Polymer 2004, 45, 217–229. [Google Scholar] [CrossRef]
- Sivan, A. New perspectives in plastic biodegradation. Curr. Opin. Biotechnol. 2011, 22, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Ojha, N.; Pradhan, N.; Singh, S.; Barla, A.; Shrivastava, A.; Khatua, P.; Rai, V.; Bose, S. Evaluation of HDPE and LDPE degradation by fungus, implemented by statistical optimization. Sci. Rep. 2017, 7, 39515. [Google Scholar] [CrossRef] [PubMed]
- Romero, E.; Speranza, M.; Garcia-Guinea, J.; Martinez, A.T.; Martinez, M.J. An anamorph of the white-rot fungus Bjerkandera adusta capable of colonizing and degrading compact disc components. FEMS Microbiol. Lett. 2007, 275, 122–129. [Google Scholar] [CrossRef]
- El-Batal, A.I.; ElKenawy, N.M.; Yassin, A.S.; Amin, M.A. Laccase production by Pleurotus ostreatus and its application in synthesis of gold nanoparticles. Biotechnol. Rep. 2015, 5, 31–39. [Google Scholar] [CrossRef]
- Crowe, J.D.; Olsson, S. Induction of laccase activity in Rhizoctonia solani by antagonistic Pseudomonas fluorescens strains and a range of chemical treatments. Appl. Environ. Microbiol. 2001, 67, 2088–2094. [Google Scholar] [CrossRef]
- J White, T.; Bruns, T.; Lee, S.; Taylor, J.; A Innis, M.; H Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academic Press Inc.: New York, NY, USA, 1990; Volume 31, pp. 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Blanquez, P.; Sarra, M.; Vicent, M.T. Study of the cellular retention time and the partial biomass renovation in a fungal decolourisation continuous process. Water Res. 2006, 40, 1650–1656. [Google Scholar] [CrossRef]
- Kang, B.R.; Kim, M.S.; Lee, T.K. Unveiling of Concealed Processes for the Degradation of Pharmaceutical Compounds by Neopestalotiopsis sp. Microorganisms 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Vilar, F.C.; Siqueira-Soares Rde, C.; Finger-Teixeira, A.; de Oliveira, D.M.; Ferro, A.P.; da Rocha, G.J.; Ferrarese Mde, L.; dos Santos, W.D.; Ferrarese-Filho, O. The acetyl bromide method is faster, simpler and presents best recovery of lignin in different herbaceous tissues than Klason and thioglycolic acid methods. PLoS ONE 2014, 9, e110000. [Google Scholar] [CrossRef] [PubMed]
- Margot, J.; Bennati-Granier, C.; Maillard, J.; Blanquez, P.; Barry, D.A.; Holliger, C. Bacterial versus fungal laccase: potential for micropollutant degradation. AMB Express 2013, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Touahar, I.E.; Haroune, L.; Ba, S.; Bellenger, J.P.; Cabana, H. Characterization of combined cross-linked enzyme aggregates from laccase, versatile peroxidase and glucose oxidase, and their utilization for the elimination of pharmaceuticals. Sci. Total Environ. 2014, 481, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Tien, M.; Kirk, T.K. Lignin peroxidase of Phanerochaete chrysosporium. In Methods in Enzymology; Academic Press: San Diego, CA, USA, 1988; Volume 161, pp. 238–249. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. 2013, Volume 3, p. 201. Available online: https://www.r-project.org/ (accessed on 11 March 2019).
- Floudas, D.; Binder, M.; Riley, R.; Barry, K.; Blanchette, R.A.; Henrissat, B.; Martinez, A.T.; Otillar, R.; Spatafora, J.W.; Yadav, J.S.; et al. The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef]
- Heinfling, A.; Martinez, M.J.; Martinez, A.T.; Bergbauer, M.; Szewzyk, U. Purification and characterization of peroxidases from the dye-decolorizing fungus Bjerkandera adusta. FEMS Microbiol. Lett. 1998, 165, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Haritash, A.K.; Kaushik, C.P. Biodegradation aspects of Polycyclic Aromatic Hydrocarbons (PAHs): A review. J. Hazard. Mater. 2009, 169, 1–15. [Google Scholar] [CrossRef]
- Dhouib, A.; Hamza, M.; Zouari, H.; Mechichi, T.; Hmidi, R.; Labat, M.; Martinez, M.J.; Sayadi, S. Screening for Ligninolytic Enzyme Production by Diverse Fungi from Tunisia. World J. Microbiol. Biotechnol. 2005, 21, 1415–1423. [Google Scholar] [CrossRef]
- de Souza, C.G.; Tychanowicz, G.K.; de Souza, D.F.; Peralta, R.M. Production of laccase isoforms by Pleurotus pulmonarius in response to presence of phenolic and aromatic compounds. J. Basic Microbiol. 2004, 44, 129–136. [Google Scholar] [CrossRef]
- Periasamy, R.; Palvannan, T. Optimization of laccase production by Pleurotus ostreatus IMI 395545 using the Taguchi DOE methodology. J. Basic Microbiol. 2010, 50, 548–556. [Google Scholar] [CrossRef]
- Adekunle, A.E.; Guo, C.; Liu, C.-Z. Lignin-Enhanced Laccase Production from Trametes versicolor. Waste Biomass Valorization 2017, 8, 1061–1066. [Google Scholar] [CrossRef]
- Janusz, G.; Kucharzyk, K.H.; Pawlik, A.; Staszczak, M.; Paszczynski, A.J. Fungal laccase, manganese peroxidase and lignin peroxidase: gene expression and regulation. Enzyme Microb. Technol. 2013, 52, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Andriani, A.; Tachibana, S. Lignocellulosic materials as solid support agents for Bjerkandera adusta SM46 to enhance polycyclic aromatic hydrocarbon degradation on sea sand and sea water media. Biocatal. Agric. Biotechnol. 2016, 8. [Google Scholar] [CrossRef]
- Bagewadi, Z.K.; Mulla, S.I.; Ninnekar, H.Z. Optimization of laccase production and its application in delignification of biomass. Int. J. Recycl. Org. Waste Agric. 2017, 6, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Brijwani, K.; Rigdon, A.; Vadlani, P.V. Fungal laccases: production, function, and applications in food processing. Enzyme Res. 2010, 2010, 149748. [Google Scholar] [CrossRef] [PubMed]
- Toran, J.; Blanquez, P.; Caminal, G. Comparison between several reactors with Trametes versicolor immobilized on lignocellulosic support for the continuous treatments of hospital wastewater. Bioresour. Technol. 2017, 243, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rodriguez, C.E.; Marco-Urrea, E.; Caminal, G. Naproxen degradation test to monitor Trametes versicolor activity in solid-state bioremediation processes. J. Hazard Mater. 2010, 179, 1152–1155. [Google Scholar] [CrossRef] [PubMed]
- Tellez-Tellez, M.; Fernandez, F.J.; Montiel-Gonzalez, A.M.; Sanchez, C.; Diaz-Godinez, G. Growth and laccase production by Pleurotus ostreatus in submerged and solid-state fermentation. Appl. Microbiol. Biotechnol. 2008, 81, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Aydinoglu, T.; Sargin, S. Production of laccase from Trametes versicolor by solid-state fermentation using olive leaves as a phenolic substrate. Bioprocess Biosyst. Eng. 2013, 36, 215–222. [Google Scholar] [CrossRef]
- Pathak, V.M.; Navneet. Review on the current status of polymer degradation: a microbial approach. Bioresour. Bioprocess. 2017, 4, 15. [Google Scholar] [CrossRef]
- Lenz, R.; Enders, K.; Stedmon, C.A.; Mackenzie, D.M.A.; Nielsen, T.G. A critical assessment of visual identification of marine microplastic using Raman spectroscopy for analysis improvement. Marine Pollut. Bull. 2015, 100, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Hiejima, Y.; Kida, T.; Takeda, K.; Igarashi, T.; Nitta, K.-h. Microscopic structural changes during photodegradation of low-density polyethylene detected by Raman spectroscopy. Polym. Degrad. Stab. 2018, 150, 67–72. [Google Scholar] [CrossRef]
- Kida, T.; Hiejima, Y.; Nitta, K.-h. Raman Spectroscopic Study of High-density Polyethylene during Tensile Deformation. Int. J. Recycl. Org. Waste Agric. 2016, 1, 1–6. [Google Scholar] [CrossRef]
- Fayolle, B.; Colin, X.; Audouin, L.; Verdu, J. Mechanism of degradation induced embrittlement in polyethylene. Polym. Degrad. Stab. 2007, 92, 231–238. [Google Scholar] [CrossRef]
- Taddei, P.; Monti, P.; Simoni, R. Vibrational and thermal study on the in vitro and in vivo degradation of a bioabsorbable periodontal membrane: Vicryl Periodontal Mesh (Polyglactin 910). J. Mater. Sci. Mater. Med. 2002, 13, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Ołdak, D.; Kaczmarek, H.; Buffeteau, T.; Sourisseau, C. Photo- and Bio-Degradation Processes in Polyethylene, Cellulose and their Blends Studied by ATR-FTIR and Raman Spectroscopies. J. Mater. Sci. 2005, 40, 4189–4198. [Google Scholar] [CrossRef]
- Sangeetha Devi, R.; Rajesh Kannan, V.; Nivas, D.; Kannan, K.; Chandru, S.; Robert Antony, A. Biodegradation of HDPE by Aspergillus spp. from marine ecosystem of Gulf of Mannar, India. Marine Pollut. Bull. 2015, 96, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Da Luz, J.M.; Paes, S.A.; Ribeiro, K.V.; Mendes, I.R.; Kasuya, M.C. Degradation of Green Polyethylene by Pleurotus ostreatus. PLoS ONE 2015, 10, e0126047. [Google Scholar] [CrossRef] [PubMed]
- Longe, L.F.; Couvreur, J.; Leriche Grandchamp, M.; Garnier, G.; Allais, F.; Saito, K. Importance of Mediators for Lignin Degradation by Fungal Laccase. ACS Sustain. Chem. Eng. 2018, 6, 10097–10107. [Google Scholar] [CrossRef]
- Munk, L.; Andersen, M.L.; Meyer, A.S. Influence of mediators on laccase catalyzed radical formation in lignin. Enzyme Microb. Technol. 2018, 116, 48–56. [Google Scholar] [CrossRef]
- Roth, S.; Spiess, A.C. Laccases for biorefinery applications: a critical review on challenges and perspectives. Bioprocess Biosyst. Eng. 2015, 38, 2285–2313. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raman shift, cm−1 | Phase | Mode |
|---|---|---|
| 1064 | Crystalline, trans chain | νas (C–C) |
| 1130 | Crystalline, trans chain | νs (C–C) |
| 1295 | Crystalline | τ (CH2) |
| 1416 | Crystalline (orthorhombic) | δ (CH2) |
| 1440 | Amorphous trans (intermediate) | δ (CH2) |
| 1460 | Amorphous | δ (CH2) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, B.R.; Kim, S.B.; Song, H.A.; Lee, T.K. Accelerating the Biodegradation of High-Density Polyethylene (HDPE) Using Bjerkandera adusta TBB-03 and Lignocellulose Substrates. Microorganisms 2019, 7, 304. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090304
Kang BR, Kim SB, Song HA, Lee TK. Accelerating the Biodegradation of High-Density Polyethylene (HDPE) Using Bjerkandera adusta TBB-03 and Lignocellulose Substrates. Microorganisms. 2019; 7(9):304. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090304
Chicago/Turabian StyleKang, Bo Ram, Soo Bin Kim, Hyun A Song, and Tae Kwon Lee. 2019. "Accelerating the Biodegradation of High-Density Polyethylene (HDPE) Using Bjerkandera adusta TBB-03 and Lignocellulose Substrates" Microorganisms 7, no. 9: 304. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090304