Phenotypic Diversity of Lactobacillus casei Group Isolates as a Selection Criterion for Use as Secondary Adjunct Starters

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Milk Fermentation and Bacterial Acidifying Performance Analyses
2.3. Analysis of Volatile Compounds
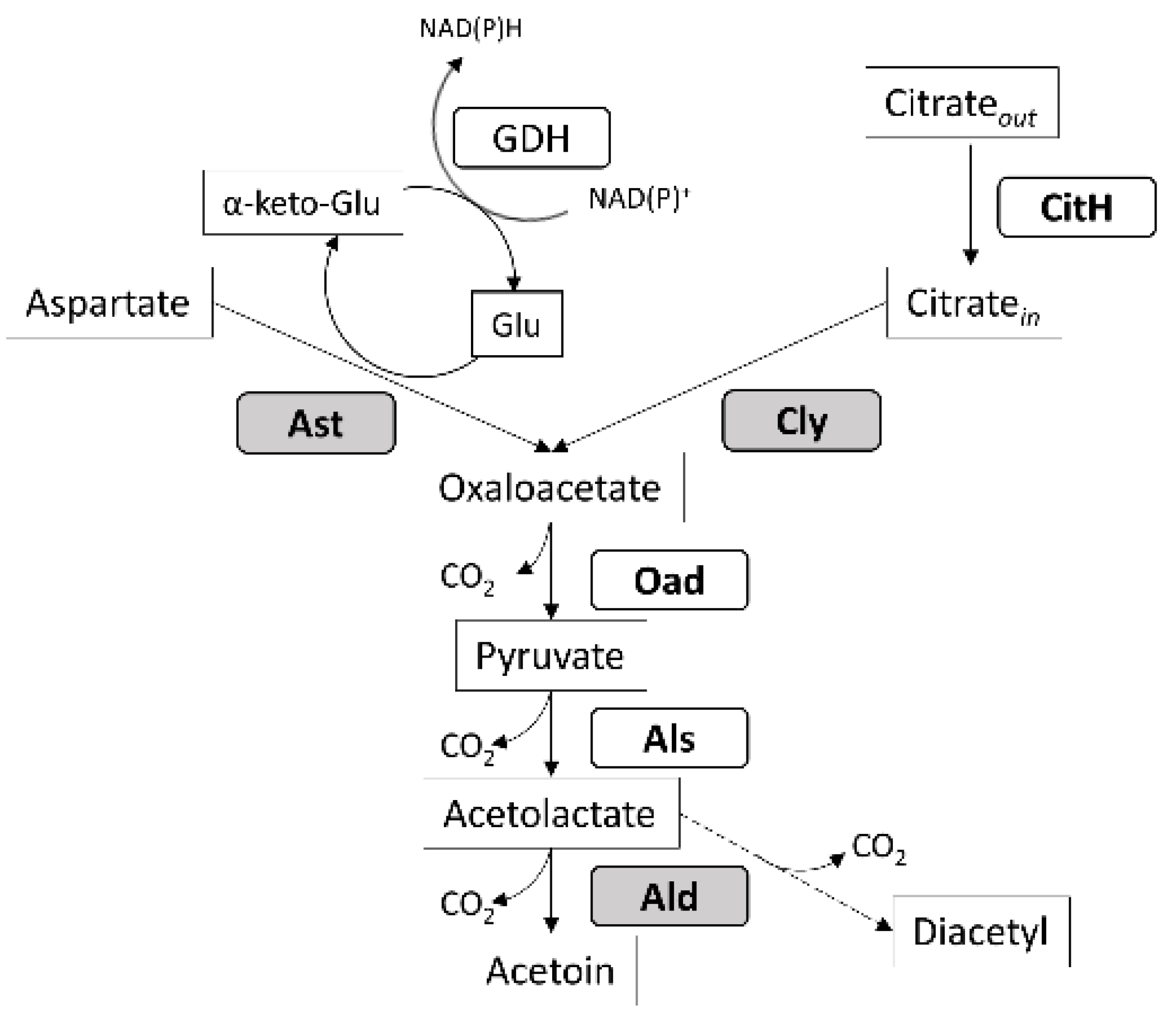
2.4. Screening for Genes Involved in Acetoin Metabolic Pathway
2.5. RNA Extraction and cDNA Synthesis
2.6. Relative Expression of Ast, Cly and Ald Genes
2.7. Statistical Analyses
3. Results and Discussion
3.1. Acidifying Properties of the Strains in Milk
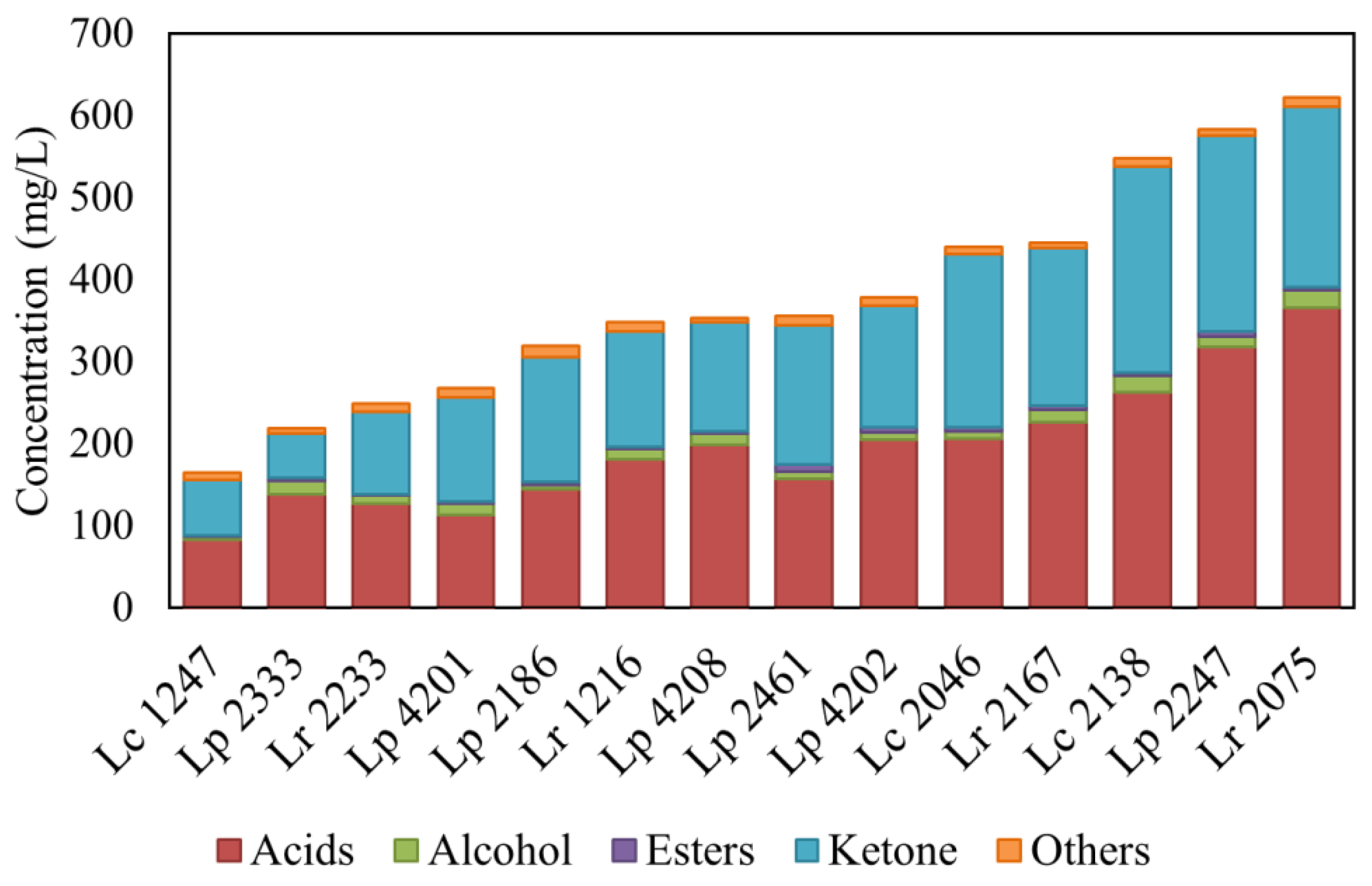
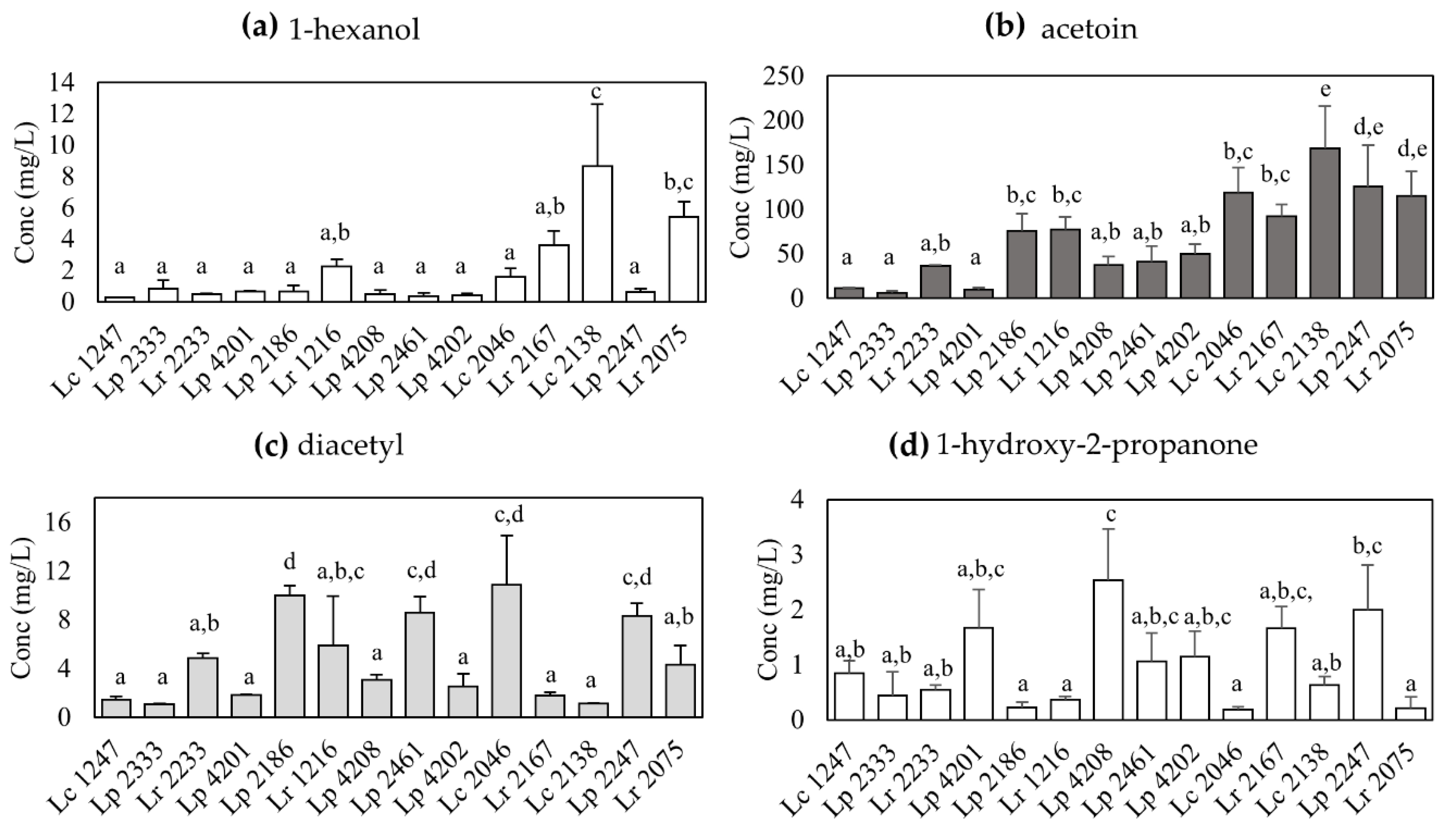
3.2. Production of Volatile Compounds during Milk Fermentation
3.3. Distribution and Expression of Genes Involved in Acetoin Production
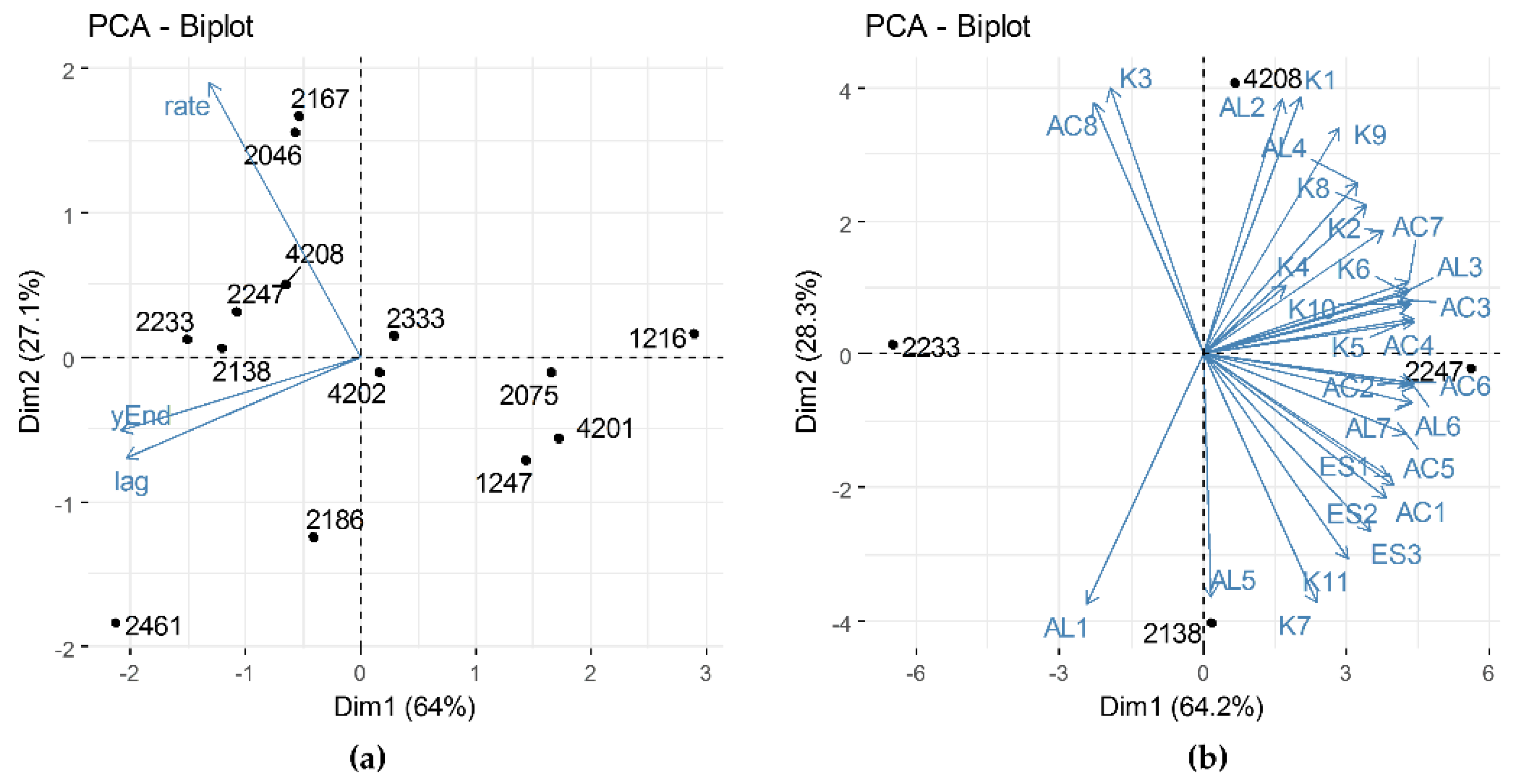
3.4. Features-Based Selection of the Strains for Use as Adjunct Cultures
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Grappin, R.; Beuvier, E. Possible implications of milk pasteurization on the manufacture and sensory quality of ripened cheese. Int. Dairy J. 1997, 7, 751–761. [Google Scholar] [CrossRef]
- Crow, V.; Curry, B.; Hayes, M. The ecology of non-starter lactic acid bacteria (NSLAB) and their use as adjuncts in New Zealand Cheddar. Int. Dairy J. 2001, 11, 275–283. [Google Scholar] [CrossRef]
- Colonna, A.; Durham, C. Factors affecting consumers’ preferences for and purchasing decisions regarding pasteurized and raw milk specialty cheeses. J. Dairy Sci. 2011, 94, 5217–5226. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Montel, M.C.; Buchin, S.; Mallet, A.; Delbes-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Smid, E.J.; Kleerebezem, M. Production of aroma compounds in lactic fermentations. Annu. Rev. Food Sci. Technol. 2014, 5, 313–326. [Google Scholar] [CrossRef]
- Pereira, G.V.D.M.; Neto, D.P.D.C.; Junqueira, A.C.D.O.; Karp, S.G.; Letti, L.A.J.; Júnior, A.I.M.; Soccol, C.R. A Review of Selection Criteria for Starter Culture Development in the Food Fermentation Industry. Food Rev. Int. 2019, 36, 135–167. [Google Scholar] [CrossRef]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef]
- El Soda, M.; Madkor, S.; Tong, P.S. Adjunct cultures: Recent developments and potential significance to the cheese industry. J. Dairy Sci. 2000, 83, 609–619. [Google Scholar] [CrossRef]
- Tungjaroenchai, W.; White, C.H.; Holmes, W.E.; Drake, M.A. Influence of adjunct cultures on volatile free fatty acids in reduced-fat edam cheeses. J. Dairy Sci. 2004, 87, 3224–3234. [Google Scholar] [CrossRef] [Green Version]
- Di Cagno, R.; De Pasquale, I.; De Angelis, M.; Buchin, S.; Rizzello, C.G.; Gobbetti, M. Use of microparticulated whey protein concentrate, exopolysaccharide-producing Streptococcus thermophilus, and adjunct cultures for making low-fat Italian Caciotta-type cheese. J. Dairy Sci. 2014, 97, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Gatti, M.; Bottari, B.; Lazzi, C.; Neviani, E.; Mucchetti, G. Invited review: Microbial evolution in raw-milk, long-ripened cheeses produced using undefined natural whey starters. J. Dairy Sci. 2014, 97, 573–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertani, G.; Levante, A.; Lazzi, C.; Bottari, B.; Gatti, M.; Neviani, E. Dynamics of a natural bacterial community under technological and environmental pressures: The case of natural whey starter for Parmigiano Reggiano cheese. Food Res. Int. 2020, 129, 108860. [Google Scholar] [CrossRef]
- Bottari, B.; Levante, A.; Neviani, E.; Gatti, M. How the fewest become the greatest. L. casei’s impact on long ripened cheeses. Front. Microbiol. 2018, 9, 2866. [Google Scholar] [CrossRef] [Green Version]
- Toh, H.; Oshima, K.; Nakano, A.; Takahata, M.; Murakami, M.; Takaki, T.; Nishiyama, H.; Igimi, S.; Hattori, M.; Morita, H. Genomic adaptation of the Lactobacillus casei group. PLoS ONE 2013, 8, e75073. [Google Scholar] [CrossRef]
- Broadbent, J.R.; Neeno-Eckwall, E.C.; Stahl, B.; Tandee, K.; Cai, H.; Morovic, W.; Horvath, P.; Heidenreich, J.; Perna, N.T.; Barrangou, R.; et al. Analysis of the Lactobacillus casei supragenome and its influence in species evolution and lifestyle adaptation. BMC Genom. 2012, 13, 533–551. [Google Scholar] [CrossRef] [Green Version]
- Bove, C.G.; De Dea Lindner, J.; Lazzi, C.; Gatti, M.; Neviani, E. Evaluation of genetic polymorphism among Lactobacillus rhamnosus non-starter Parmigiano Reggiano cheese strains. Int. J. Food Microbiol. 2011, 144, 569–572. [Google Scholar] [CrossRef]
- Bancalari, E.; Savo Sardaro, M.L.; Levante, A.; Marseglia, A.; Caligiani, A.; Lazzi, C.; Neviani, E.; Gatti, M. An integrated strategy to discover Lactobacillus casei group strains for their potential use as aromatic starters. Food Res. Int. 2017, 100, 682–690. [Google Scholar] [CrossRef]
- Bancalari, E.; Bernini, V.; Bottari, B.; Neviani, E.; Gatti, M. Application of Impedance Microbiology for Evaluating Potential Acidifying Performances of Starter Lactic Acid Bacteria to Employ in Milk Transformation. Front. Microbiol. 2016, 7, 1628. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.N. The changes produced by the growth of bacteria in the molecular concentration and electrical conductivity of culture media. J. Exp. Med. 1899, 4, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Bashir, R. Electrical/electrochemical impedance for rapid detection of foodborne pathogenic bacteria. Biotechnol. Adv. 2008, 26, 135–150. [Google Scholar] [CrossRef]
- Bancalari, E.; D’Incecco, P.; Savo Sardaro, M.L.; Neviani, E.; Pellegrino, L.; Gatti, M. Impedance microbiology to speed up the screening of lactic acid bacteria exopolysaccharide production. Int. J. Food Microbiol. 2019, 306, 108268. [Google Scholar] [CrossRef]
- Pogacic, T.; Maillard, M.B.; Leclerc, A.; Hervé, C.; Chuat, V.; Valence, F.; Thierry, A. Lactobacillus and Leuconostoc volatilomes in cheese conditions. Appl. Microbiol. Biotechnol. 2016, 100, 2335–2346. [Google Scholar] [CrossRef]
- Barbieri, G.; Bolzoni, L.; Careri, M.; Mangia, A.; Parolari, G.; Spagnoli, S.; Virgili, R. Study of the volatile fraction of Parmesan cheese. J. Agric. Food Chem. 1994, 42, 1170–1176. [Google Scholar] [CrossRef]
- McSweeney, P.L.H. Biochemistry of cheese ripening: Introduction and overview. Cheese Chem. Phys. Microbiol. 2004, 1, 347–360. [Google Scholar]
- Le Bars, D.; Yvon, M. Formation of diacetyl and acetoin by Lactococcus lactis via aspartate catabolism. J. Appl. Microbiol. 2008, 104, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Lo, R.; Ho, V.T.T.; Bansal, N.; Turner, M.S. The genetic basis underlying variation in production of the flavour compound diacetyl by Lactobacillus rhamnosus strains in milk. Int. J. Food Microbiol. 2018, 265, 30–39. [Google Scholar] [CrossRef]
- Kieronczyk, A.; Skeie, S.; Langsrud, T.; Le Bars, D.; Yvon, M. The nature of aroma compounds produced in a cheese model by glutamate dehydrogenase positive Lactobacillus INF15D depends on its relative aminotransferase activities towards the different amino acids. Int. Dairy J. 2004, 14, 227–235. [Google Scholar] [CrossRef]
- Tanous, C.; Kieronczyk, A.; Helinck, S.; Chambellon, E.; Yvon, M. Glutamate dehydrogenase activity: A major criterion for the selection of flavour-producing lactic acid bacteria strains. Antonie Van Leeuwenhoek 2002, 82, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.G.; Withers, S.E.; Brechany, E.Y.; Banks, J.M. Glutamate dehydrogenase activity in lactobacilli and the use of glutamate dehydrogenase-producing adjunct Lactobacillus spp. cultures in the manufacture of cheddar cheese. J. Appl. Microbiol. 2006, 101, 1062–1075. [Google Scholar] [CrossRef] [PubMed]
- Peralta, G.H.; Wolf, I.V.; Perotti, M.C.; Bergamini, C.V.; Hynes, E.R. Formation of volatile compounds, peptidolysis and carbohydrate fermentation by mesophilic lactobacilli and streptoccocci cultures in a cheese extract. Dairy Sci. Technol. 2016, 96, 603–621. [Google Scholar] [CrossRef] [Green Version]
- Monnet, C.; Schmitt, P.; Divies, C. Diacetyl production in milk by an α-acetolactic acid accumulating strain of Lactococcus lactis ssp. lactis biovar. diacetylactis. J. Dairy Sci. 1994, 77, 2916–2924. [Google Scholar] [CrossRef]
- Monnet, C.; Corrieu, G. Selection and properties of Lactobacillus mutants producing α-acetolactate. J. Dairy Sci. 1998, 81, 2096–2102. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Species | Cheese | Cheese Ripening Time (Months) |
|---|---|---|---|
| 1247 | L. casei | Parmigiano Reggiano | 12 |
| 2138 | L. casei | Grana Padano | 6 |
| 2046 | L. casei | Grana Padano | 2 |
| 2247 | L. paracasei | Parmigiano Reggiano | 1 |
| 2333 | L. paracasei | Parmigiano Reggiano | 6 |
| 2186 | L. paracasei | Grana Padano | 9 |
| 2461 | L. paracasei | Grana Padano | 2 |
| 4201 | L. paracasei | Pecorino toscano | 2 |
| 4202 | L. paracasei | Pecorino toscano | 2 |
| 4208 | L. paracasei | Pecorino toscano | 2 |
| 2233 | L. rhamnosus | Parmigiano Reggiano | 1 |
| 1216 | L. rhamnosus | Parmigiano Reggiano | 12 |
| 2167 | L. rhamnosus | Grana Padano | 9 |
| 2075 | L. rhamnosus | Grana Padano | 2 |
| Target Name | Primer | Sequence (5′-3′) 1 | Tm (°C) | PCR Product Size (bp) | Efficiency (E) |
|---|---|---|---|---|---|
| alpha-acetolactate decarboxylase (ald) | Ald_F | TTCGAAGCCAAGAATATGSCC | 55 | 156 | 0.93 |
| Ald_R | GCYTCRGGTCAAAAAGAATT | ||||
| aspartate aminotransferase (ast) | Ast_F | CAATAACTRATMCGCGCATG | 50 | 143 | 1.13 |
| Ast_R | TTCCCGAAYATYAAGCG | ||||
| citrate lyase (subunit alpha) (cly) | Cly_F | GACAGGTTYTTGATYCCCATT | 55 | 148 | 1.22 |
| Cly_R | AACTMGTGGCGTCAATTCA | ||||
| 16S rRNA | TBA_FW 2 | CGGCAACGAGCGCAACCC | 60 | 130 | 0.99 |
| TBA_RV 2 | CCATTGTAGCACGTGTGTAGCC |
| Species | Origin | Strain | Lag | Rate | yEnd |
|---|---|---|---|---|---|
| L. casei | Parmigiano Reggiano cheese | 1247 | 3.43 ± 0.32 e | 0.76 ± 0.02 a | 28.09 ± 0.56 b,c |
| L. casei | Grana Padano cheese | 2046 | 3.18 ± 0.26 d,e | 1.43 ± 0.01 e | 29.51 ± 0.10 c,d |
| L. casei | Grana Padano cheese | 2138 | 7.11 ± 0.04 g | 1.19 ± 0.01 d | 29.30 ± 0.30 c,d |
| L. paracasei | Grana Padano cheese | 2186 | 3.00 ± 0.13 d,e | 0.80 ± 0.05 b | 30.37 ± 0.38 c,d |
| L. paracasei | Parmigiano Reggiano cheese | 2247 | 6.09 ± 0.17 a | 0.61 ± 0.01a | 23.01 ± 0.17 a |
| L. paracasei | Parmigiano Reggiano cheese | 2333 | 2.38 ± 0.15 c,d | 1.03 ± 0.00 c | 29.62 ± 0.02 c,d |
| L. paracasei | Grana Padano cheese | 2461 | 8.63 ± 0.74 h | 0.86 ± 0.03 b | 30.91 ± 0.06 d |
| L. paracasei | Pecorino Toscano cheese | 4201 | 2.12 ± 0.38 c | 0.75 ± 0.02 a | 28.24 ± 0.44 b,c |
| L. paracasei | Pecorino Toscano cheese | 4202 | 4.78 ± 0.34 f | 0.84 ± 0.01 b | 21.70 ± 0.30 a |
| L. paracasei | Pecorino Toscano cheese | 4208 | 5.31 ± 0.88 f | 1.22 ± 0.01 d | 29.19 ± 0.82 c,d |
| L. rhamnosus | Parmigiano Reggiano cheese | 1216 | 1.20 ± 0.07 a,b | 0.80 ± 0.00 b | 26.69 ± 0.42 b |
| L. rhamnosus | Grana Padano cheese | 2075 | 1.98 ± 0.12 b,c | 0.86 ± 0.01 b | 28.14 ± 0.18 b,c |
| L. rhamnosus | Grana Padano cheese | 2167 | 3.61 ± 0.27 e | 1.45 ± 0.01 e | 29.17 ± 0.28 c,d |
| L. rhamnosus | Parmigiano Reggiano cheese | 2233 | 6.69 ± 0.25 g | 1.22 ± 0.01 d | 29.88 ± 0.06 c,d |
| Target | Strains | Ratio | Ratio SD | Efficiency |
|---|---|---|---|---|
| ald | 2138 | 1.108 | 0.012 | 0.93 |
| 2333 | ||||
| ast | 2138 | 1.743 | 0.015 | 1.13 |
| 2333 | ||||
| cly | 2138 | 0.324 | 0.014 | 1.22 |
| 2333 | ||||
| 16S rRNA | 2138 | 1.000 | 0.012 | 0.99 |
| 2333 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levante, A.; Bancalari, E.; Tambassi, M.; Lazzi, C.; Neviani, E.; Gatti, M. Phenotypic Diversity of Lactobacillus casei Group Isolates as a Selection Criterion for Use as Secondary Adjunct Starters. Microorganisms 2020, 8, 128. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010128
Levante A, Bancalari E, Tambassi M, Lazzi C, Neviani E, Gatti M. Phenotypic Diversity of Lactobacillus casei Group Isolates as a Selection Criterion for Use as Secondary Adjunct Starters. Microorganisms. 2020; 8(1):128. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010128
Chicago/Turabian StyleLevante, Alessia, Elena Bancalari, Martina Tambassi, Camilla Lazzi, Erasmo Neviani, and Monica Gatti. 2020. "Phenotypic Diversity of Lactobacillus casei Group Isolates as a Selection Criterion for Use as Secondary Adjunct Starters" Microorganisms 8, no. 1: 128. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010128