Salicylic Acid-Producing Endophytic Bacteria Increase Nicotine Accumulation and Resistance against Wildfire Disease in Tobacco Plants


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Plant Materials and Isolation of EB
2.2. Measurement of SA Production in Isolated EB and Host Plants
2.3. Growth of Tobacco Species Inoculated with SA-Producing EB
2.4. Disease Assay of Pst-Infected Tobacco Species
2.5. Pathogenicity Test of the Isolated SA-Producing EB
2.6. Reisolation and Confirmation of SA-Producing EB
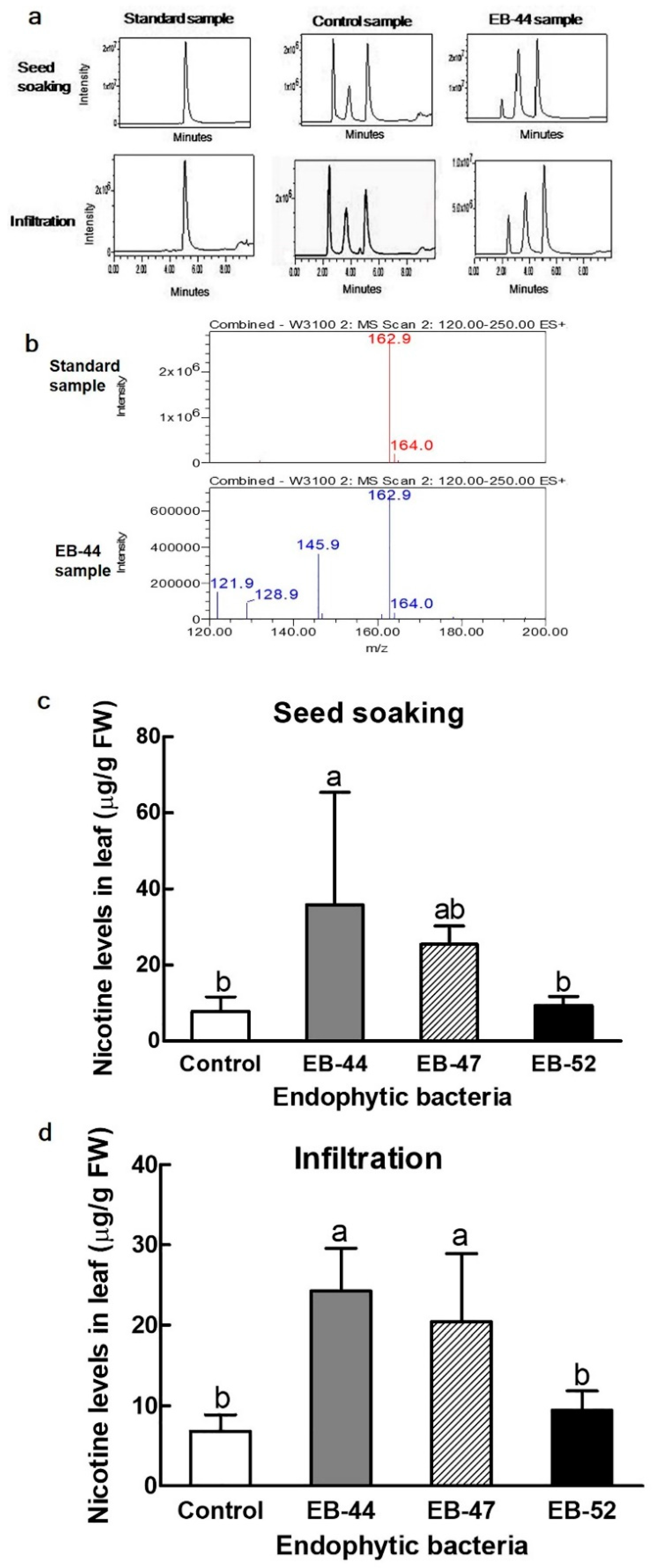
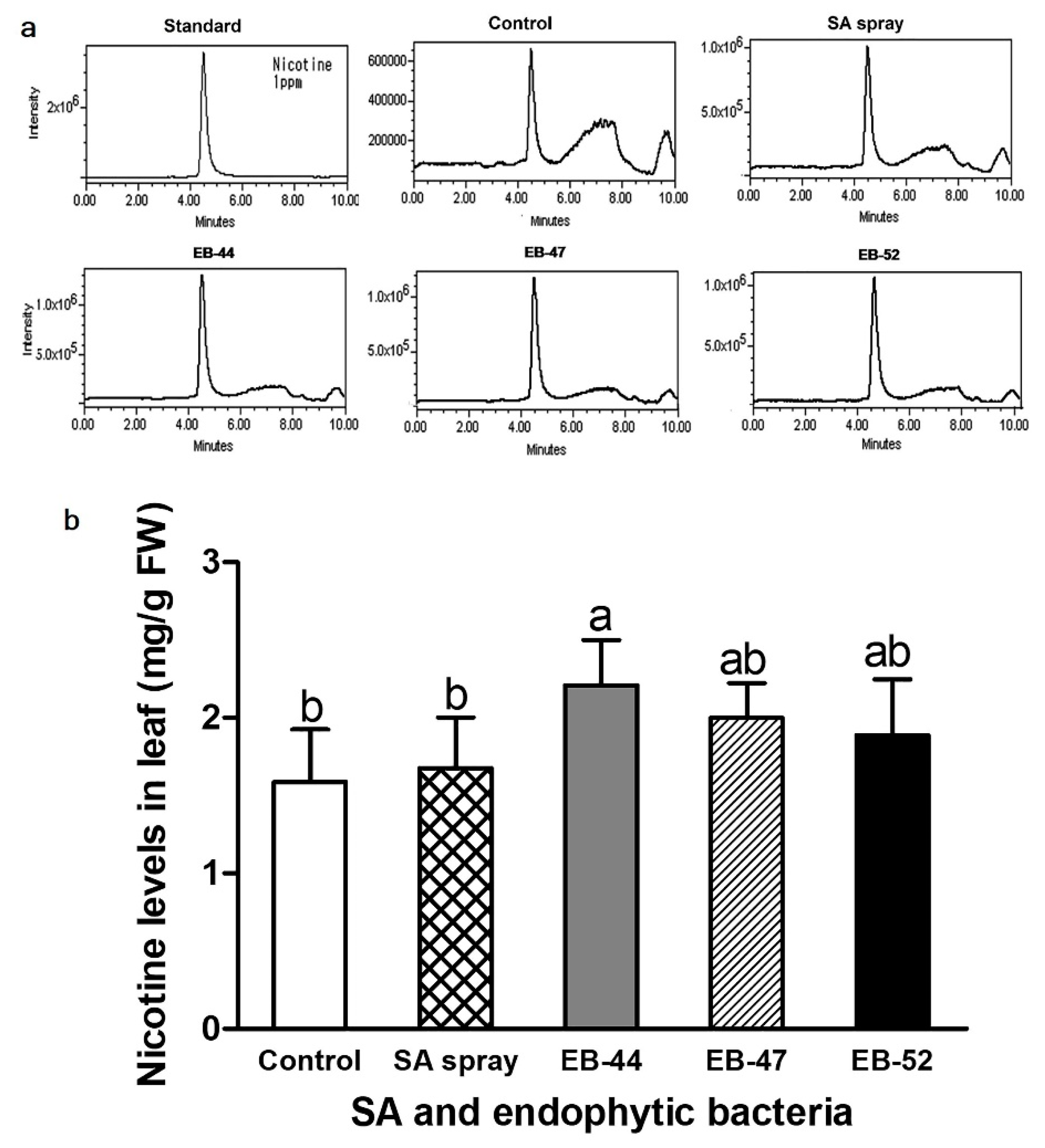
2.7. Quantification of Nicotine production in Tobacco Species Co-Cultivated with EB
2.8. Statistical Analysis
3. Results
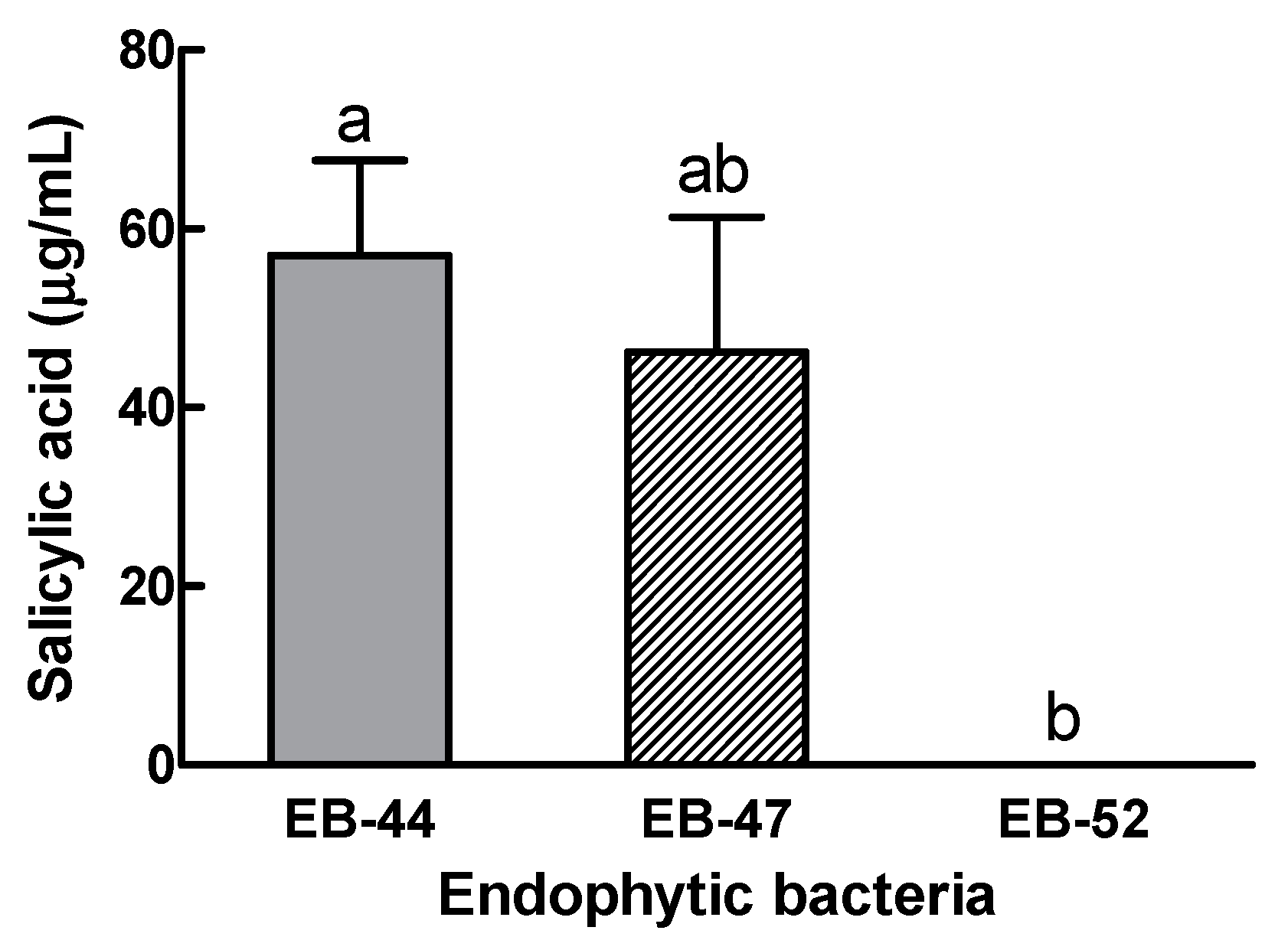
3.1. Isolation of EB and Measurement of SA in EB and the Host Plants
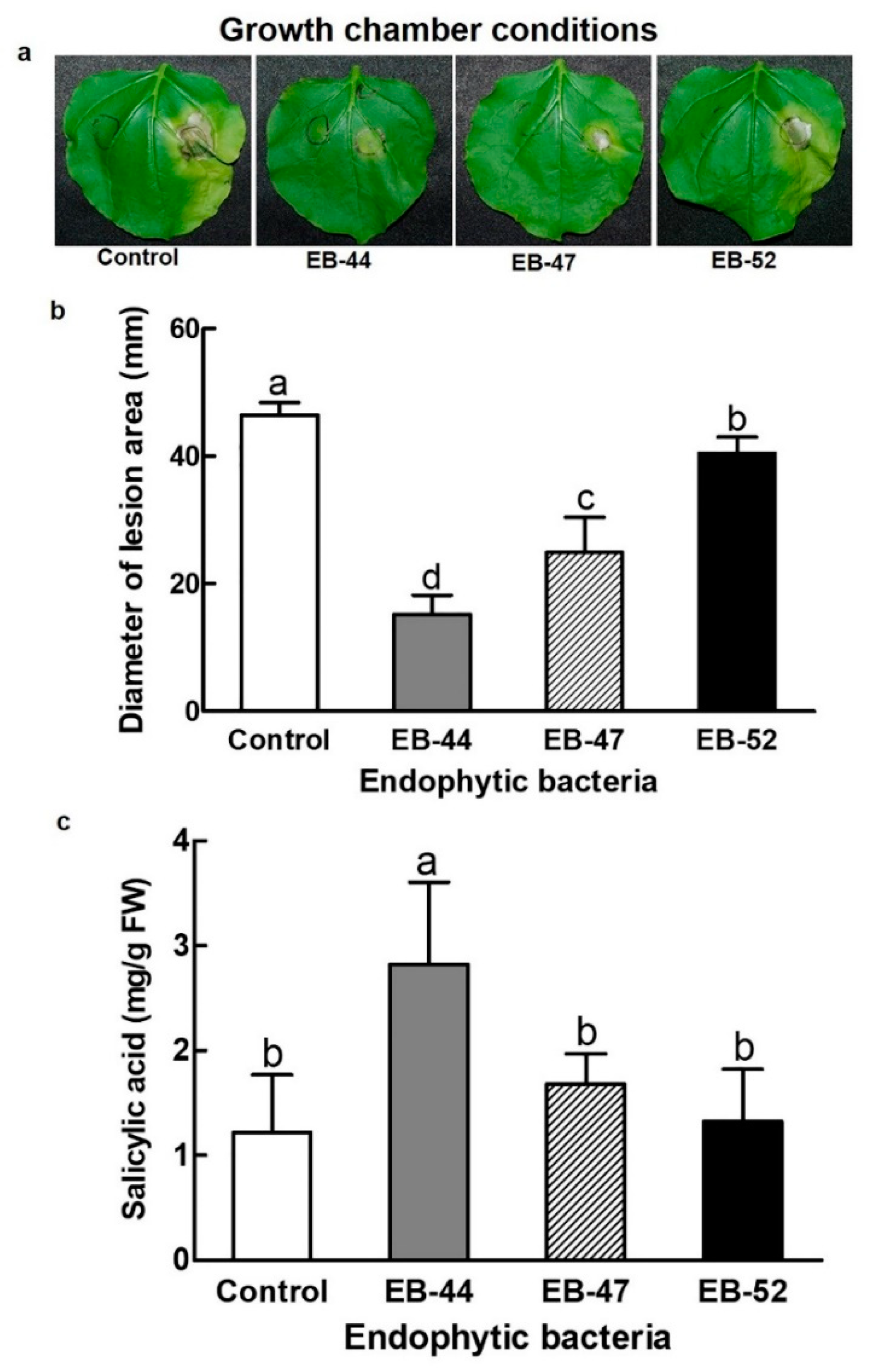
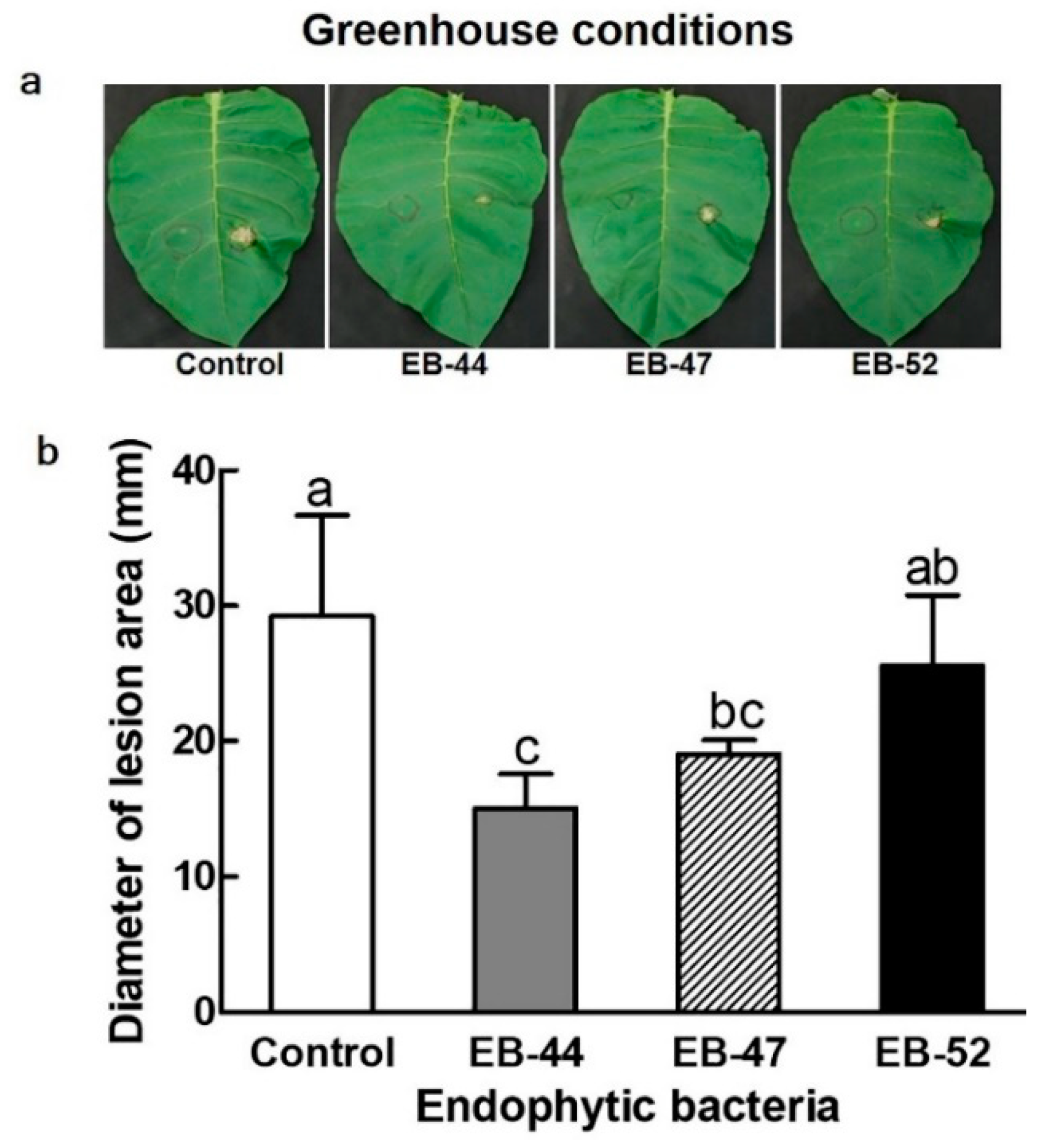
3.2. Disease Resistance in Nicotiana plants Grown from Seeds Treated with SA-Producing EB
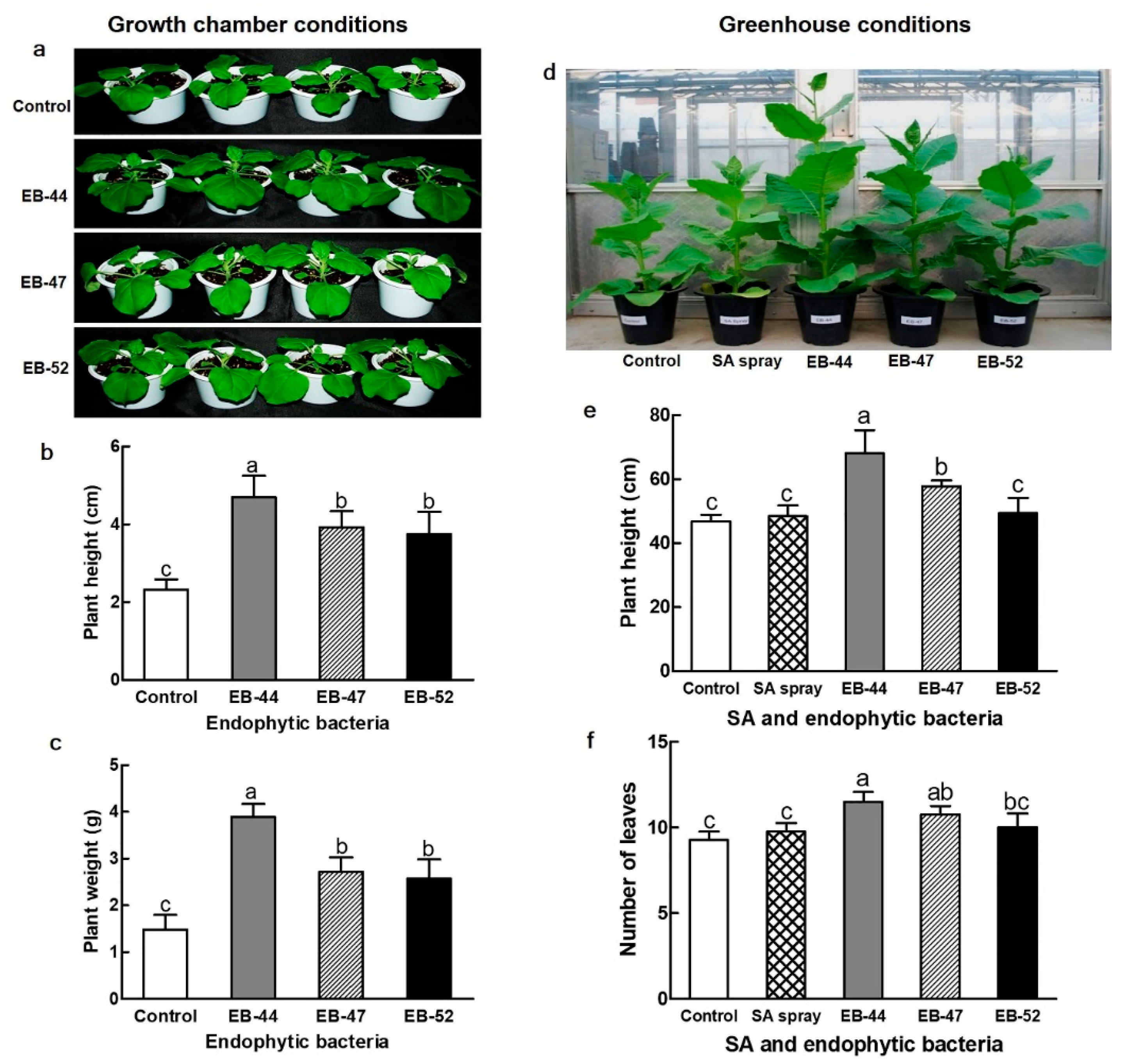
3.3. Growth Promotion of Nicotiana Plants Cultivated from Seeds Treated with SA-Producing EB
3.4. Nicotine Accumulation in Nicotiana Inoculated with SA-Producing EB
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kloepper, J.W.; Beauchamp, C.J. A review of issues related to measuring colonization of plant roots by bacteria. Can. J. Microbiol. 1992, 38, 1219–1232. [Google Scholar] [CrossRef]
- Meguro, A.; Ohmura, Y.; Hasegawa, S.; Shimizu, M.; Nishimura, T.; Kunoh, H. An endophytic actinomycete, Streptomyces sp. MBR-52, that accelerates emergence and elongation of plant adventitious roots. Actinomycetologica 2006, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gibert, A.; Volaire, F.; Barre, P.; Hazard, L. A fungal endophyte reinforces population adaptive differentiation in its host grass species. New Phytol. 2012, 194, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Nakagawa, Y.; Sato, Y.; Furumai, T.; Igarashi, Y.; Onaka, H.; Yoshida, R.; Kunoh, H. Studies on Endophytic Actinomycetes (I) Streptomyces sp. isolated from Rhododendron and Its antifungal activity. J. Gen. Plant Pathol. 2000, 66, 360–366. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Conn, V.M.; Walker, A.R.; Franco, C.M.M. Endophytic actinobacteria induce defense pathway in Arab. thaliana. Mol. Plant Microbe Interact. 2008, 21, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Khalaf, E.M.; Raizada, M.N. Bacterial seed endophytes of domesticated cucurbits antagonize fungal and oomycete pathogens including powdery mildew. Front. Microbiol. 2018, 9, 1–18. [Google Scholar]
- Terhonen, E.; Blumenstein, K.; Kovalchuk, A.; Asiegbu, F.O. Forest tree microbiomes and associated fungal endophytes: Functional roles and impact on forest health. Forests 2019, 10, 1–33. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, G.; Höfte, M. Salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 induces resistance to leaf infection by Botrytis cinerea on bean. Phytopathology 1997, 87, 588–593. [Google Scholar] [CrossRef] [Green Version]
- Press, C.M.; Wilson, M.; Tuzun, S.; Kloepper, J.W. Salicylic acid produced by Serratia marcescens 90-166 is not the primary determinant of induced systemic resistance in Cucumber or Tobacco. Mol. Plant Microbe Interact. 1997, 10, 761–768. [Google Scholar] [CrossRef] [Green Version]
- An, C.; Mou, Z. Salicylic acid and its function in plant immunity. J. Integr. Plant Biol. 2011, 53, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Uppalapati, S.R.; Ryu, C.M.; Allen, S.N.; Kang, L.; Tang, Y.; Mysore, K.S. Salicylic acid and systemic acquired resistance play a role in attenuating crown gall disease caused by Agrobacterium tumefaciens. Plant Physiol. 2008, 146, 703–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denancé, N.; Sánchez-Vallet, A.; Goffner, D.; Molina, A. Disease resistance or growth: The role of plant hormones in balancing immune responses and fitness costs. Front. Plant Sci. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.L.; Yang, Y.; He, Z. Roles of plant hormones and their interplay in rice immunity. Mol. Plant 2013, 6, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.G.; Debbage, J.M. Tabtoxin-induced symptoms are associated with the accumulation of ammonia formed during photorespiration. Physiol. Plant Pathol. 1982, 20, 223–233. [Google Scholar] [CrossRef]
- Studholme, D.J.; Ibanez, S.G.; MacLean, D.; Dangl, J.L.; Chang, J.H.; Rathjen, J.P. A draft genome sequence and functional screen reveals the repertoire of type III secreted proteins of Pseudomonas syringae pathovar tabaci 11528. BMC Genom. 2009, 10, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.D.; Zhang, Z.S.; Sun, X.B.; Zhao, M.; Sun, G.Y.; Chow, W.S. Photoinhibition and photoinhibition-like damage to the photosynthetic apparatus in tobacco leaves induced by pseudomonas syringae pv. tabaci under light and dark conditions. BMC Plant Biol. 2016, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, P.B. Wildfire and Angular Leaf Spot. In Compendium of Tobacco Diseases; Shew, H.D., Lucas, G.B., Eds.; APS Press: Saint Paul, MN, USA, 1991; pp. 30–32. [Google Scholar]
- Valleau, W.D. Breeding tobacco for disease resistance. Econ. Bot. 1952, 6, 69–102. [Google Scholar] [CrossRef]
- McManus, P.S.; Stockwell, V.O.; Sundin, G.W.; Jones, A.L. Antibiotic use in plant agriculture. Annu. Rev. Phytopathol. 2002, 40, 443–465. [Google Scholar] [CrossRef]
- Carter, A.P.; Clemons, W.M.; Brodersen, D.E.; Morgan-Warren, R.J.; Wimberly, B.T.; Ramakrishnan, V. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature 2000, 407, 340–348. [Google Scholar] [CrossRef]
- Hyun, J.W.; Kim, H.J.; Yi, P.H.; Hwang, R.Y.; Park, E.W. Mode of action of streptomycin resistance in the citrus canker pathogen (Xanthomonas smithii subsp. citri) in Jeju island. Plant Pathol. J. 2012, 28, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Tao, J.; Liu, T.; Liu, Y.; Xiao, N.; Li, T.; Gu, Y.; Yin, H.; Meng, D. Responses of phyllosphere microbiota and plant health to application of two different biocontrol agents. AMB Express 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doolittle, D.J.; Winegar, R.; Lee, C.K.; Caldwell, W.S.; Hayes, A.W.; De Bethizy, J.D. The genotoxic potential of nicotine and its major metabolites. Mutat. Res. 1995, 344, 95–102. [Google Scholar] [CrossRef]
- Bush, L.P.; Fannin, R.L.; Chelvarajan, H.R.B. Biosynthesis and metabolism of nicotine and related alkaloids. In Nicotine and Related Alkaloids: Absorbtion, Distribution, Metabolism and Excretion; Garrod, J.W., Wahren, J., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 1–30. [Google Scholar]
- Wink, M.W. Modes of action of alkaloids. In Alkaloids: Biochemistry, Ecology and Medicinal Applications; Robers, M.F., Wink, M., Eds.; Plenum Press: New York, NY, USA, 1998; pp. 301–3325. [Google Scholar]
- Ziebell, H.; Murphy, A.M.; Groen, S.C.; Tungadi, T.; Westwood, J.H.; Lewsey, M.G.; Moulin, M.; Leczkowski, A.; Smith, A.G.; Stevens, M.; et al. Cucumber mosaic virus and its 2b RNA silencing suppressor modify plant-aphid interactions in tobacco. Sci. Rep. 2011, 1, 1–7. [Google Scholar] [CrossRef]
- Zhang, L.; Oh, Y.; Li, H.; Baldwin, I.T.; Galis, I. Alternative oxidase in resistance to biotic stresses: Nicotiana attenuata AOX contributes to resistance to a pathogen and a piercing-sucking insect but not manduca sexta larvae. Plant Physiol. 2012, 160, 1453–1467. [Google Scholar] [CrossRef] [Green Version]
- Vogel-Adghough, D.; Stahl, E.; Návarová, H.; Zeier, J. Pipecolic acid enhances resistance to bacterial infection and primes salicylic acid and nicotine accumulation in tobacco. Plant Signal. Behav. 2013, 8, e26366. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Teng, W.; Shi, Q.; Zhang, F. Multiple signals regulate nicotine synthesis in tobacco plant. Plant Signal. Behav. 2007, 2, 280–281. [Google Scholar] [CrossRef] [Green Version]
- Ruan, A.; Min, H.; Peng, X.; Huang, Z. Isolation and characterization of Pseudomonas sp. strain HF-1, capable of degrading nicotine. Res. Microbiol. 2005, 156, 700–706. [Google Scholar] [CrossRef]
- Chen, C.; Li, X.; Yang, J.; Gong, X.; Li, B.; Zhang, K.Q. Isolation of nicotine-degrading bacterium Pseudomonas sp. Nic22, and its potential application in tobacco processing. Int. Biodeterior. Biodegrad. 2008, 62, 226–231. [Google Scholar] [CrossRef]
- Yuan, Y.J.; Lu, Z.X.; Wu, N.; Huang, L.J.; Lü, F.X.; Bie, X.M. Isolation and preliminary characterization of a novel nicotine-degrading bacterium, Ochrobactrum intermedium DN2. Int. Biodeterior. Biodegrad. 2005, 56, 45–50. [Google Scholar] [CrossRef]
- Gong, X.-W.; Yang, J.-K.; Duan, Y.-Q.; Dong, J.-Y.; Zhe, W.; Wang, L.; Li, Q.-H.; Zhang, K.-Q. Isolation and characterization of Rhodococcus sp. Y22 and its potential application to tobacco processing. Res. Microbiol. 2009, 160, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.N.; Liu, Z.; Xu, P. Biodegradation of nicotine by a newly isolated Agrobacterium sp. strain S33. J. Appl. Microbiol. 2009, 107, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.E.D.O.; de Queiroz, M.V.; Borges, A.C.; de Moraes, C.A.; de Araújo, E.F. Isolation and characterization of endophytic bacteria isolated from the leaves of the common bean (Phaseolus vulgaris). Braz. J. Microbiol. 2012, 43, 1562–1575. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susilowati, R.; Sabdono, A.; Widowati, I. Isolation and characterization of bacteria associated with brown algae Sargassum spp. from Panjang island and their antibacterial activities. Procedia Environ. Sci. 2015, 23, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, R.; Park, E.; Lee, S.W.; Baek, K.H. Soybean (Glycine max L. Merr.) sprouts germinated under red light irradiation induce disease resistance against bacterial rotting disease. PLoS ONE 2015, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadt-Hallmann, A.; Kloepper, J.W.; Benhamou, N. Bacterial endophytes in cotton: Mechanisms of entering the plant. Can. J. Microbiol. 1997, 43, 577–582. [Google Scholar] [CrossRef]
- Hallmann, J.; Quadt-Hallmann, A.; Miller, W.G.; Sikora, R.A.; Lindow, S.E. Endophytic colonization of plants by the biocontrol agent Rhizobium etli G12 in relation to Meloidogyne incognita infection. Phytopathology 2002, 91, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Sturz, A.V.; Nowak, J. Endophytic communities of rhizobacteria and the strategies required to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Serino, L.; Reimmann, C.; Baur, H.; Beyeler, M.; Visca, P.; Haas, D. Structural genes for salicylate biosynthesis from chorismate in Pseudomonas aeruginosa. Mol. Gen. Genet. 1995, 249, 217–228. [Google Scholar] [CrossRef]
- Yalpani, N.; Silverman, P.; Wilson, T.M.A.; Kleier, D.A.; Raskin, L. Salicylic acid is a systemic signal and an inducer of pathogenesis-related proteins in virus-infected tobacco. Plant Cell 1991, 3, 809–818. [Google Scholar]
- Yalpani, N.; Shulaev, V.; Raskin, I. Endogenous salicylic acid levels correlate with accumulation of pathogenesis-related proteins and virus resistance in tobacco. Phytopathology 1993, 83, 702–708. [Google Scholar] [CrossRef]
- Coquoz, J.-L.; Buchala, A.; Métraux, J.-P. The biosynthesis of salicylic acid in potato plants. Plant Physiol. 1998, 117, 1095–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarre, D.A.; Mayo, D. Differential characteristics of salicylic acid-mediated signaling in potato. Physiol. Mol. Plant Pathol. 2004, 64, 179–188. [Google Scholar] [CrossRef]
- Delaney, T.P.; Uknes, S.; Vernooij, B.; Friedrich, L.; Weymann, K.; Negrotto, D.; Gaffney, T.; Gut-Rella, M.; Kessmann, H.; Ward, E.; et al. A central role of salicylic acid in plant disease resistance. Science 1994, 266, 1247–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Maurhofer, M.; Hase, C.; Meuwly, P.; Metraux, J.P.; Defago, G. Induction of systemic resistance of tobacco to tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens strain CHA0: Influence of the gacA gene and of pyoverdine production. Phytopathology 1994, 84, 139–146. [Google Scholar] [CrossRef]
- Chasan, R. SA: Source or signal for SAR? Plant Cell 1995, 7, 1519–1521. [Google Scholar] [CrossRef] [Green Version]
- Shulaev, V.; Silverman, P.; Raskin, I. Airborne signalling by methyl salicylate in plant pathogen resistance. Nature 1997, 385, 718–721. [Google Scholar] [CrossRef]
- Park, S.W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Santner, A.; Calderon-Villalobos, L.I.A.; Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef]
- Wolters, H.; Jürgens, G. Survival of the flexible: Hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 2009, 10, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, J.; Grúz, J.; Bačkor, M.; Strnad, M.; Repčák, M. Salicylic acid-induced changes to growth and phenolic metabolism in Matricaria chamomilla plants. Plant Cell Rep. 2009, 28, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, B.; Leitner, E.; Pfannhauser, W. Determination of the nicotine content of various edible nightshades (Solanaceae) and their products and estimation of the associated dietary nicotine intake. J. Agric. Food Chem. 1999, 47, 3113–3120. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Jack, A.M.; Lewis, R.S.; Dewey, R.E.; Bush, L.P. Nicotine biosynthesis, metabolism and translocation in tobacco as determined by nicotine demethylase mutants. Phytochemistry 2013, 95, 188–196. [Google Scholar] [CrossRef]
- Li, X.; Guo, H.; Qi, Y.; Liu, H.; Zhang, X.; Ma, P.; Liang, Z.; Dong, J. Salicylic acid-induced cytosolic acidification increases the accumulation of phenolic acids in Salvia miltiorrhiza cells. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 126, 333–341. [Google Scholar] [CrossRef]
- De Luca, V.; St Pierre, B. The cell and developmental biology of alkaloid biosynthesis. Trends Plant Sci. 2000, 5, 168–173. [Google Scholar] [CrossRef]
- Shoji, T.; Yamada, Y.; Hashimoto, T. Jasmonate induction of putrescine N-methyltransferase genes in the root of Nicotiana sylvestris. Plant Cell Physiol. 2000, 41, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-B.; Bokowiec, M.T.; Rushton, P.J.; Han, S.-C.; Timko, M.P. Tobacco transcription factors NtMYC2a and NtMYC2b form nuclear complexes with the NtJAZ1 repressor and regulate multiple jasmonate-inducible steps in nicotine biosynthesis. Mol. Plant 2012, 5, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Nugroho, L.H.; Peltenburg-Looman, A.M.G.; De Vos, H.; Verberne, M.C.; Verpoorte, R. Nicotine and related alkaloids accumulation in constitutive salicylic acid producing tobacco plants. Plant Sci. 2002, 162, 575–581. [Google Scholar] [CrossRef]
- Preston, C.A.; Lewandowski, C.; Enyedi, A.J.; Baldwin, I.T. Tobacco mosaic virus inoculation inhibits wound-induced jasmonic acid-mediated responses within but not between plants. Planta 1999, 209, 87–95. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Total no. of Isolates | Tissue | No. of Isolates | CFUs/g of Fresh Tissue | No. of EB Isolates | |
|---|---|---|---|---|---|---|
| SA | No SA | |||||
| Metasequoia glyptostroboides | 8 | Leaves | 4 | 3.3 × 104 | 0 | 4 |
| Cones | 4 | 1.2 × 103 | 2 | 2 | ||
| Ginkgo biloba | 50 | Leaves | 11 | 1.8 × 103 | 1 | 10 |
| Fruits | 39 | 6.0 × 104 | 1 | 38 | ||
| Taxus brevifolia | Leaves | 23 | 8.6 × 104 | 2 | 21 | |
| 41 | Seeds | 8 | 7.2 × 103 | 0 | 8 | |
| Fruits | 10 | 1.8 × 102 | 1 | 9 | ||
| Pinus densiflora | 2 | Leaves | 2 | 9.6 × 103 | 0 | 2 |
| Salix babylonica | 24 | Leaves | 24 | 1.5 × 105 | 20 | 4 |
| Salix chaenomeloides | 9 | Leaves | 9 | 5.9 × 104 | 0 | 9 |
| Total | 134 | 134 | - | 27 | 107 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.N.; Ali, M.S.; Choi, S.-J.; Park, Y.-I.; Baek, K.-H. Salicylic Acid-Producing Endophytic Bacteria Increase Nicotine Accumulation and Resistance against Wildfire Disease in Tobacco Plants. Microorganisms 2020, 8, 31. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010031
Islam MN, Ali MS, Choi S-J, Park Y-I, Baek K-H. Salicylic Acid-Producing Endophytic Bacteria Increase Nicotine Accumulation and Resistance against Wildfire Disease in Tobacco Plants. Microorganisms. 2020; 8(1):31. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010031
Chicago/Turabian StyleIslam, Md. Nurul, Md. Sarafat Ali, Seong-Jin Choi, Youn-Il Park, and Kwang-Hyun Baek. 2020. "Salicylic Acid-Producing Endophytic Bacteria Increase Nicotine Accumulation and Resistance against Wildfire Disease in Tobacco Plants" Microorganisms 8, no. 1: 31. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010031