Renin Angiotensin System, COVID-19 and Male Fertility: Any Risk for Conceiving?

 , , ,
, , ,
Abstract
:1. Introduction: SARS-CoV-2 and COVID-19 Pandemic
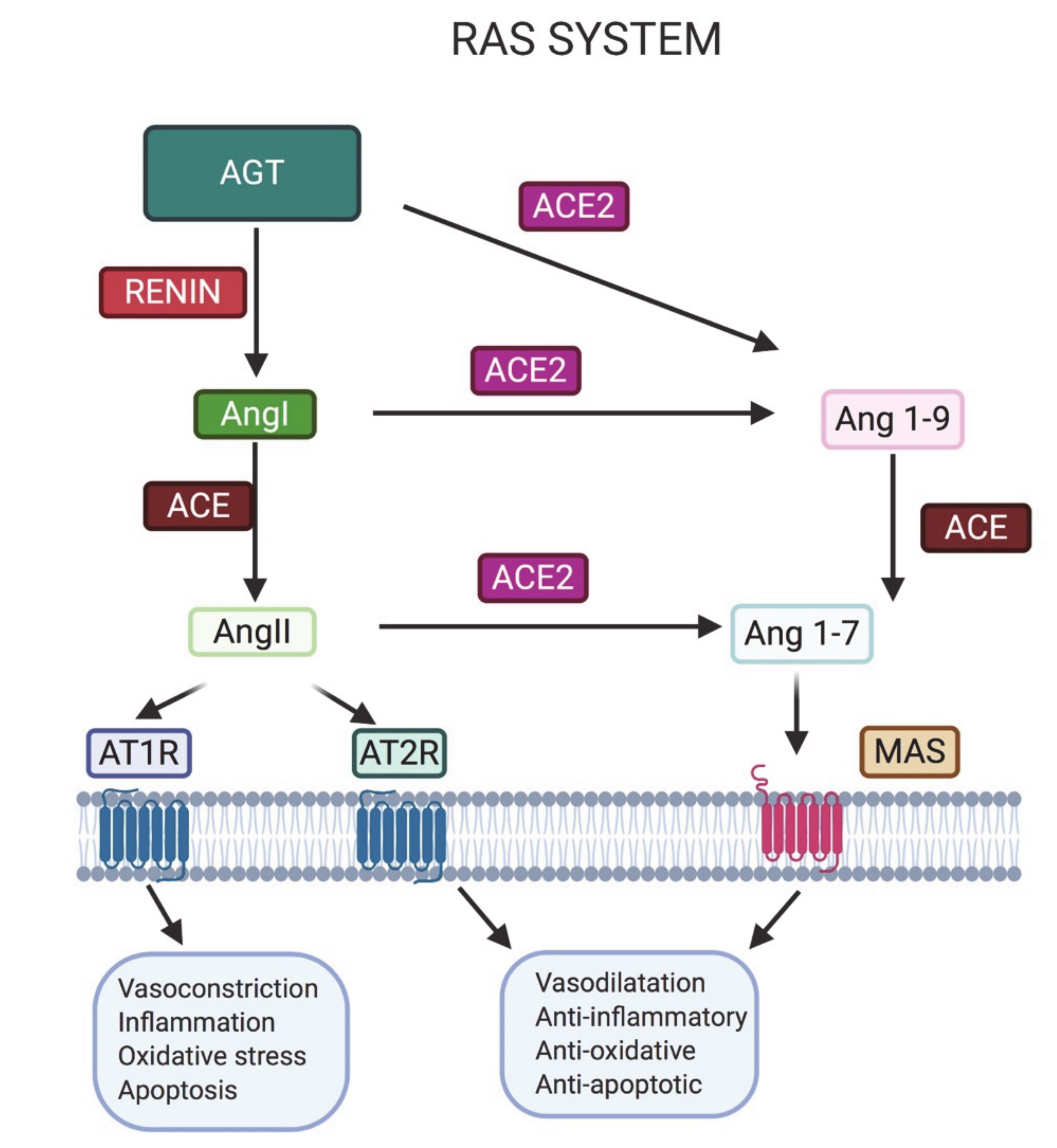
2. Importance of the Renin–Angiotensin System (RAS) in Male Fertility
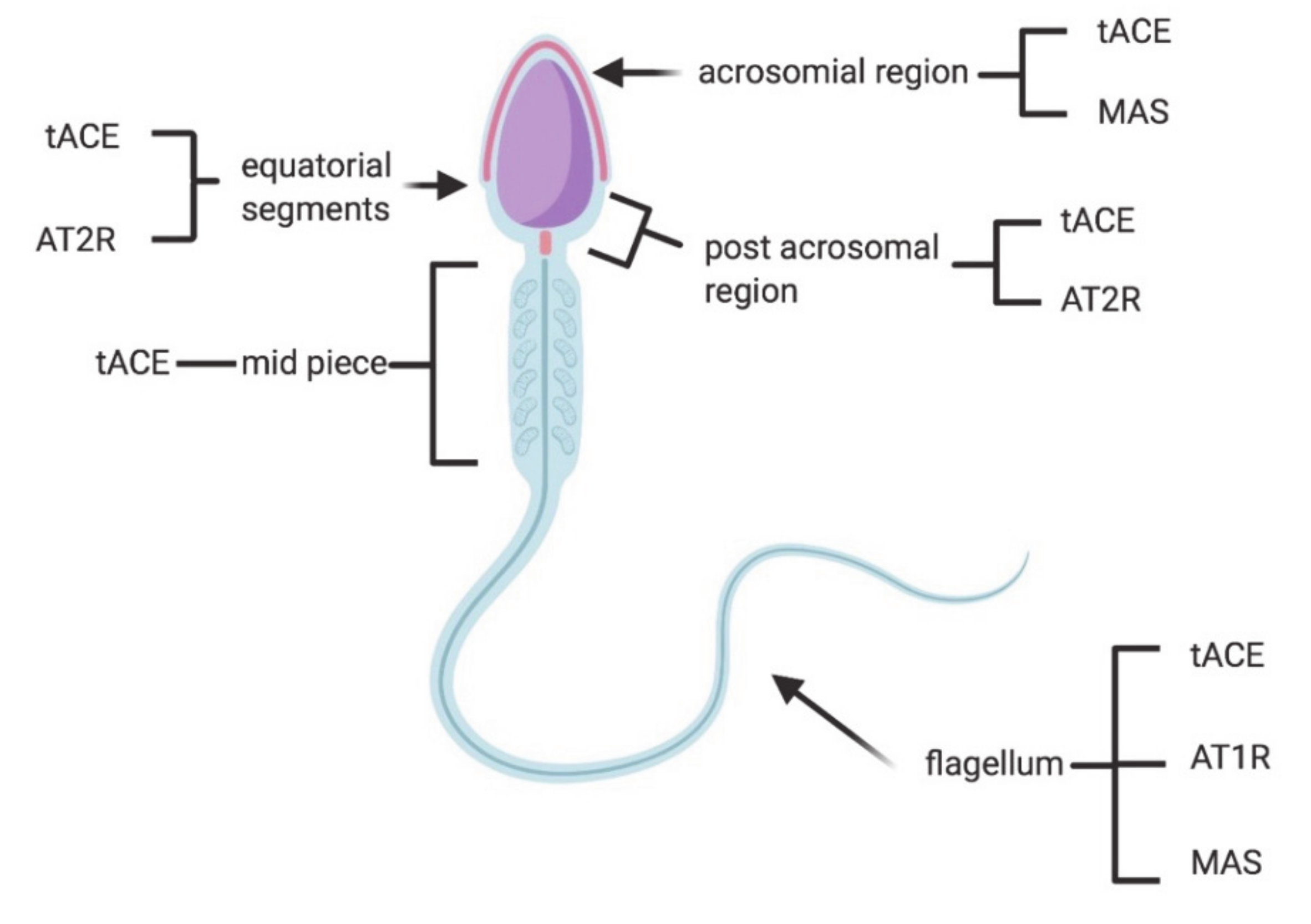
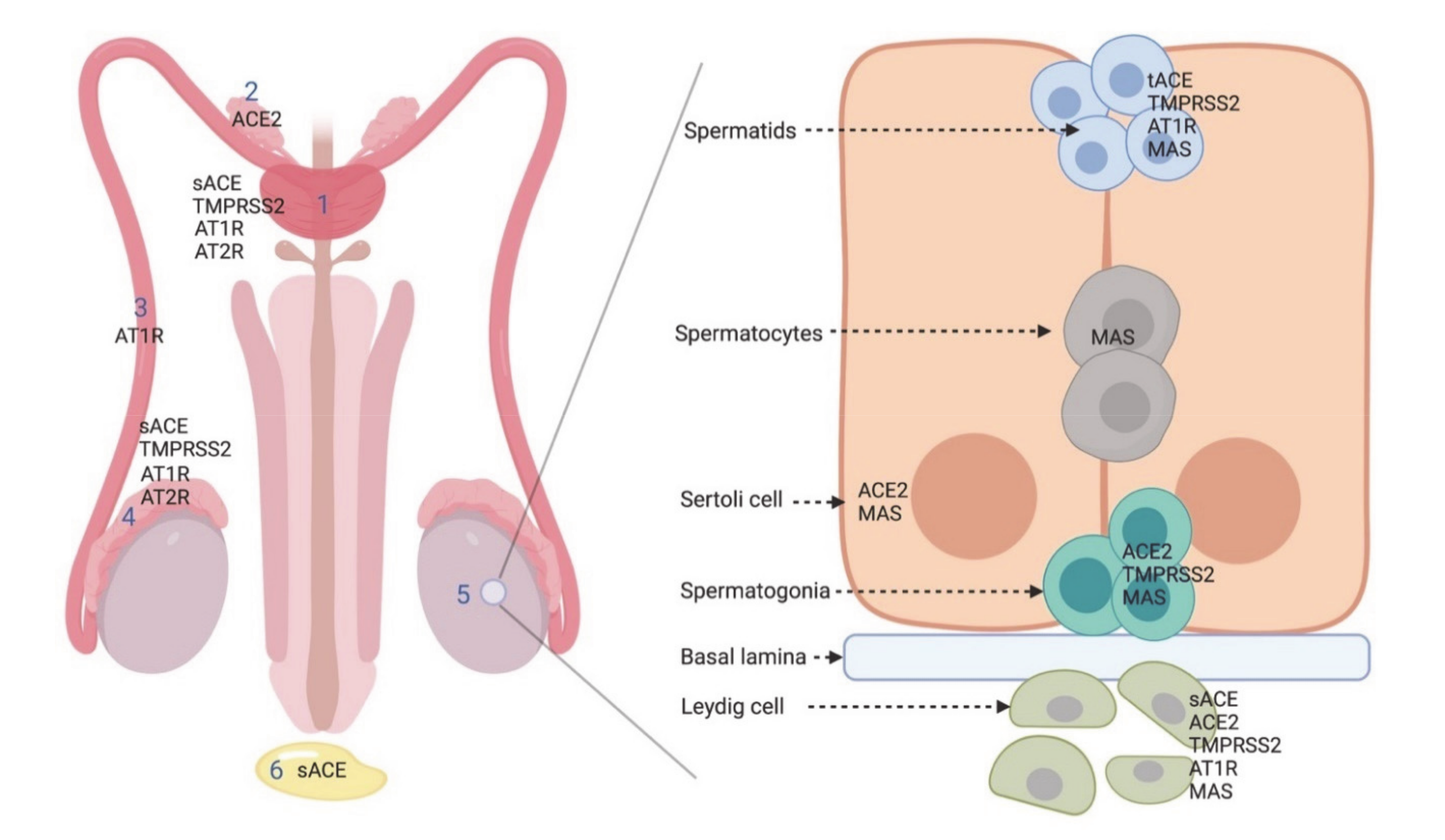
2.1. ACE2 in the Male Reproductive System
2.2. ACE
2.3. AT1R and AT2R
2.4. MAS
2.5. Drugs Acting on RAS System
3. TMPRSS2 in the Male Reproductive System: Another Actor in SARS-CoV-2 Infection
4. SARS-CoV-2 Detection in the Reproductive System and Effects on Fertility
5. Safety of Samples in PMA Procedures
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, S.; Lin, Q.; Ran, J.; Musa, S.S.; Yang, G.; Wang, W.; Lou, Y.; Gao, D.; Yang, L.; He, D.; et al. Preliminary estimation of the basic reproduction number of novel coronavirus (2019-nCoV) in China, from 2019 to 2020: A data-driven analysis in the early phase of the outbreak. Int. J. Infect. Dis. 2020, 92, 214–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO World Health Organization. Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 26 August 2020).
- Coleman, C.M.; Frieman, M.B. Coronaviruses: Important Emerging Human Pathogens. J. Virol. 2014, 88, 5209–5212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathewson, A.C.; Bishop, A.; Yao, Y.; Kemp, F.; Ren, J.; Chen, H.; Xu, X.; Berkhout, B.; van der Hoek, L.; Jones, I.M. Interaction of severe acute respiratory syndrome-coronavirus and NL63 coronavirus spike proteins with angiotensin converting enzyme-2. J. Gen. Virol. 2008, 89, 2741–2745. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pascolo, L.; Zupin, L.; Melato, M.; Tricarico, P.M.; Crovella, S. TMPRSS2 and ACE2 Coexpression in SARS-CoV-2 Salivary Glands Infection. J. Dent. Res. 2020, 99, 1120–1121. [Google Scholar] [CrossRef] [PubMed]
- Malle, L. A map of SARS-CoV-2 and host cell interactions. Nat. Rev. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef]
- Zupin, L.; Pascolo, L.; Crovella, S. Is FURIN gene expression in salivary glands related to SARS-CoV-2 infectivity through saliva? J. Clin. Pathol. 2020. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Krüger, N.; Müller, M.; Drosten, C.; Pöhlmann, S. The novel coronavirus 2019 (2019-nCoV) uses the SARS-coronavirus receptor ACE2 and the cellular protease TMPRSS2 for entry into target cells. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Roche, J.A.; Roche, R. A hypothesized role for dysregulated bradykinin signaling in COVID-19 respiratory complications. FASEB J. 2020, 34, 7265–7269. [Google Scholar] [CrossRef] [PubMed]
- Kai, H.; Kai, M. Interactions of coronaviruses with ACE2, angiotensin II, and RAS inhibitors—lessons from available evidence and insights into COVID-19. Hypertens. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Huang, D.; Yu, H.; Zhu, Z.; Xia, Z.; Su, Y.; Li, Z.; Zhou, G.; Gou, J.; Qu, J.; et al. COVID-19: Abnormal liver function tests. J. Hepatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Durvasula, R.; Wellington, T.; McNamara, E.; Watnick, S. COVID-19 and Kidney Failure in the Acute Care Setting: Our Experience From Seattle. Am. J. Kidney Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Long, B.; Brady, W.J.; Koyfman, A.; Gottlieb, M. Cardiovascular complications in COVID-19. Am. J. Emerg. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Neto, A.N.; Monteiro, R.A.A.; Silva, L.F.F.; Malheiros, D.M.A.C.; Oliveira, E.P.; Theodoro-Filho, J.; Pinho, J.R.R.; Gomes-Gouvêa, M.S.; Salles, A.P.M.; Oliveira, I.R.S.; et al. Pulmonary and systemic involvement in COVID-19 patients assessed with ultrasound-guided minimally invasive autopsy. Histopathology 2020, 77, 186–197. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Dolhnikoff, M.; Duarte-Neto, A.N.; Almeida Monteiro, R.A.; Silva, L.F.F.; Oliveira, E.P.; Saldiva, P.H.N.; Mauad, T.; Negri, E.M. Pathological evidence of pulmonary thrombotic phenomena in severe COVID-19. J. Thromb. Haemost. 2020, 18, 1517–1519. [Google Scholar] [CrossRef] [Green Version]
- van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 2020, 323, 1843–1844. [Google Scholar] [CrossRef] [Green Version]
- Oran, D.P.; Topol, E.J. Prevalence of Asymptomatic SARS-CoV-2 Infection: A Narrative Review. Ann. Intern. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsson, J.F. Systematic review of COVID-19 in children shows milder cases and a better prognosis than adults. Acta Paediatr. 2020, 109, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-M.; Bai, P.; He, W.; Wu, F.; Liu, X.-F.; Han, D.-M.; Liu, S.; Yang, J.-K. Gender Differences in Patients With COVID-19: Focus on Severity and Mortality. Front. Public Health 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, T.; Wang, Y.; Wang, Z.; Liang, Y.; Huang, T.; Zhang, H.; Sun, W.; Wang, Y. COVID-19 patients’ clinical characteristics, discharge rate, and fatality rate of meta-analysis. J. Med. Virol. 2020, 92, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ji, H.; Zheng, W.; Wu, X.; Zhu, J.J.; Arnold, A.P.; Sandberg, K. Sex differences in renal angiotensin converting enzyme 2 (ACE2) activity are 17β-oestradiol-dependent and sex chromosome-independent. Biol. Sex Differ. 2010, 1, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, R.; Mao, S.-Q.; Gu, T.-L.; Zhong, F.-D.; Gong, M.-L.; Hao, L.-M.; Yin, F.-Y.; Dong, C.-Z.; Zhang, L.-N. Preliminary analysis of the association between methylation of the ACE2 promoter and essential hypertension. Mol. Med. Rep. 2017, 15, 3905–3911. [Google Scholar] [CrossRef] [Green Version]
- Carrel, L.; Willard, H.F. X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature 2005, 434, 400–404. [Google Scholar] [CrossRef]
- Lucas, J.M.; Heinlein, C.; Kim, T.; Hernandez, S.A.; Malik, M.S.; True, L.D.; Morrissey, C.; Corey, E.; Montgomery, B.; Mostaghel, E.; et al. The Androgen-Regulated Protease TMPRSS2 Activates a Proteolytic Cascade Involving Components of the Tumor Microenvironment and Promotes Prostate Cancer Metastasis. Cancer Discov. 2014, 4, 1310–1325. [Google Scholar] [CrossRef] [Green Version]
- Bukowska, A.; Spiller, L.; Wolke, C.; Lendeckel, U.; Weinert, S.; Hoffmann, J.; Bornfleth, P.; Kutschka, I.; Gardemann, A.; Isermann, B.; et al. Protective regulation of the ACE2/ACE gene expression by estrogen in human atrial tissue from elderly men. Exp. Biol. Med. 2017, 242, 1412–1423. [Google Scholar] [CrossRef]
- Gagliardi, M.C.; Tieri, P.; Ortona, E.; Ruggieri, A. ACE2 expression and sex disparity in COVID-19. Cell Death Discov. 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Seddighzadeh, B.; Cooperberg, M.R.; Huang, F.W. Expression of ACE2, the SARS-CoV-2 Receptor, and TMPRSS2 in Prostate Epithelial Cells. Eur. Urol. 2020, 78, 296–298. [Google Scholar] [CrossRef] [PubMed]
- Zupin, L.; Pascolo, L.; Zito, G.; Ricci, G.; Crovella, S. SARS-CoV-2 and the next generations: Which impact on reproductive tissues? J. Assist. Reprod. Genet. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, L.; Feng, Z.; Wan, S.; Huang, P.; Sun, X.; Wen, F.; Huang, X.; Ning, G.; Wang, W. Comparative genetic analysis of the novel coronavirus (2019-nCoV/SARS-CoV-2) receptor ACE2 in different populations. Cell Discov. 2020, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Jabeen, N.; Raza, F.; Shabbir, S.; Baig, A.A.; Amanullah, A.; Aziz, B. Structural variations in human ACE2 may influence its binding with SARS-CoV-2 spike protein. J. Med Virol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darbani, B. The Expression and Polymorphism of Entry Machinery for COVID-19 in Human: Juxtaposing Population Groups, Gender, and Different Tissues. Int. J. Environ. Res. Public Health 2020, 17, 3433. [Google Scholar] [CrossRef] [PubMed]
- Asselta, R.; Paraboschi, E.M.; Mantovani, A.; Duga, S. ACE2 and TMPRSS2 variants and expression as candidates to sex and country differences in COVID-19 severity in Italy. Aging 2020, 12, 10087–10098. [Google Scholar] [CrossRef]
- Lopera Maya, E.A.; van der Graaf, A.; Lanting, P.; van der Geest, M.; Fu, J.; Swertz, M.; Franke, L.; Wijmenga, C.; Deelen, P.; Zhernakova, A.; et al. Lack of Association Between Genetic Variants at ACE2 and TMPRSS2 Genes Involved in SARS-CoV-2 Infection and Human Quantitative Phenotypes. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Atlas, S.A. The Renin-Angiotensin Aldosterone System: Pathophysiological Role and Pharmacologic Inhibition. J. Manag. Care Pharm. 2007, 13, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Fountain, J.H.; Lappin, S.L. Physiology, Renin Angiotensin System. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Rüster, C.; Wolf, G. Renin-angiotensin-aldosterone system and progression of renal disease. J. Am. Soc. Nephrol. 2006, 17, 2985–2991. [Google Scholar] [CrossRef]
- Esther, C.R.; Howard, T.E.; Marino, E.M.; Goddard, J.M.; Capecchi, M.R.; Bernstein, K.E. Mice lacking angiotensin-converting enzyme have low blood pressure, renal pathology, and reduced male fertility. Lab. Invest. 1996, 74, 953–965. [Google Scholar] [PubMed]
- Paul, M.; Poyan Mehr, A.; Kreutz, R. Physiology of Local Renin-Angiotensin Systems. Physiological Reviews 2006, 86, 747–803. [Google Scholar] [CrossRef] [PubMed]
- Ramaraj, P.; Kessler, S.P.; Colmenares, C.; Sen, G.C. Selective restoration of male fertility in mice lacking angiotensin-converting enzymes by sperm-specific expression of the testicular isozyme. J. Clin. Investig. 1998, 102, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrario, C.M.; Ahmad, S.; Nagata, S.; Simington, S.W.; Varagic, J.; Kon, N.; Dell’italia, L.J. An evolving story of angiotensin-II-forming pathways in rodents and humans. Clin. Sci. 2014, 126, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Castilho, A.C.S.; Fontes, P.K.; Franchi, F.F.; Santos, P.H.; Razza, E.M. Renin-Angiotensin System on Reproductive Biology. In Renin-Angiotensin System-Past, Present and Future; Tolekova, A.N., Ed.; InTech: London, UK, 2017; ISBN 978-953-51-3351-3. [Google Scholar]
- Pandey, K.N.; Maki, M.; Inagami, T. Detection of renin mRNA in mouse testis by hybridization with renin cDNA probe. Biochem. Biophys. Res. Commun. 1984, 125, 662–667. [Google Scholar] [CrossRef]
- Deschepper, C.F.; Mellon, S.H.; Cumin, F.; Baxter, J.D.; Ganong, W.F. Analysis by immunocytochemistry and in situ hybridization of renin and its mRNA in kidney, testis, adrenal, and pituitary of the rat. Proc. Natl. Acad. Sci. USA 1986, 83, 7552–7556. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.N.; Inagami, T. Regulation of renin angiotensins by gonadotropic hormones in cultured murine Leydig tumor cells. Release of angiotensin but not renin. J. Biol. Chem. 1986, 261, 3934–3938. [Google Scholar]
- Herr, D.; Bekes, I.; Wulff, C. Local Renin-Angiotensin System in the Reproductive System. Front. Endorcrinol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Passos-Silva, D.G.; Verano-Braga, T.; Santos, R.A.S. Angiotensin-(1–7): Beyond the cardio-renal actions. Clin. Sci. 2013, 124, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.B.; Araújo, F.C.; Pereira, V.M.; Dos Reis, A.M.; Santos, R.A.; Reis, F.M. Angiotensin (1-7) and its receptor Mas are expressed in the human testis: Implications for male infertility. J. Mol. Histol. 2010, 41, 75–80. [Google Scholar] [CrossRef]
- Pan, P.-P.; Zhan, Q.-T.; Le, F.; Zheng, Y.-M.; Jin, F. Angiotensin-Converting Enzymes Play a Dominant Role in Fertility. Int. J. Mol. Sci. 2013, 14, 21071–21086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, P.; Sernia, C. The renin-angiotensin system and male reproduction: New functions for old hormones. J. Mol. Endocrinol. 2003, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinson, G.P.; Mehta, J.; Evans, S.; Matthews, S.; Puddefoot, J.R.; Saridogan, E.; Holt, W.V.; Djahanbakhch, O. Angiotensin II stimulates sperm motility. Regul. Pept. 1996, 67, 131–135. [Google Scholar] [CrossRef]
- Pauls, K.; Metzger, R.; Steger, K.; Klonisch, T.; Danilov, S.; Franke, F.E. Isoforms of angiotensin I-converting enzyme in the development and differentiation of human testis and epididymis. Andrologia 2003, 35, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Sibony, M.; Gasc, J.M.; Soubrier, F.; Alhenc-Gelas, F.; Corvol, P. Gene expression and tissue localization of the two isoforms of angiotensin I converting enzyme. Hypertension 1993, 21, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Li, K.; Ding, Y.; Lu, W.L.; Wang, J. ACE2 Expression in Kidney and Testis May Cause Kidney and Testis Damage After 2019-nCoV Infection. MedRxiv 2020. [Google Scholar] [CrossRef]
- Sabeur, K.; Vo, A.T.; Ball, B.A. Effects of angiotensin II on the acrosome reaction in equine spermatozoa. J. Reprod. Fertil. 2000, 120, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Dinh, D.T.; Frauman, A.G.; Sourial, M.; Casley, D.J.; Johnston, C.I.; Fabiani, M.E. Identification, distribution, and expression of angiotensin II receptors in the normal human prostate and benign prostatic hyperplasia. Endorcrinology 2001, 142, 1349–1356. [Google Scholar] [CrossRef]
- Gianzo, M.; Muñoa-Hoyos, I.; Urizar-Arenaza, I.; Larreategui, Z.; Quintana, F.; Garrido, N.; Subirán, N.; Irazusta, J. Angiotensin II type 2 receptor is expressed in human sperm cells and is involved in sperm motility. Fertil. Steril. 2016, 105, 608–616. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, A.; Cortés, L.; Beitia, M.; Totorikaguena, L.; Agirregoitia, N.; Corcostegui, B.; Casis, L.; Matorras, R.; Irazusta, J.; Agirregoitia, E. Role of Angiotensin-(1-7) via MAS receptor in human sperm motility and acrosome reaction. Reproduction 2020, 159, 241–249. [Google Scholar] [CrossRef]
- Vaarala, M.H.; Porvari, K.S.; Kellokumpu, S.; Kyllönen, A.P.; Vihko, P.T. Expression of transmembrane serine protease TMPRSS2 in mouse and human tissues. J. Pathol. 2001, 193, 134–140. [Google Scholar] [CrossRef]
- Lucas, J.; True, L.; Hawley, S.; Matsumura, M.; Morrissey, C.; Vessella, R.; Nelson, P. The androgen-regulated type II serine protease TMPRSS2 is differentially expressed and mislocalized in prostate adenocarcinoma. J. Pathol. 2008, 215, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-W.; Lee, M.-S.; Lucht, A.; Chou, F.-P.; Huang, W.; Havighurst, T.C.; Kim, K.; Wang, J.-K.; Antalis, T.M.; Johnson, M.D.; et al. TMPRSS2, a Serine Protease Expressed in the Prostate on the Apical Surface of Luminal Epithelial Cells and Released into Semen in Prostasomes, Is Misregulated in Prostate Cancer Cells. Am. J. Pathol. 2010, 176, 2986–2996. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, X. scRNA-seq Profiling of Human Testes Reveals the Presence of the ACE2 Receptor, A Target for SARS-CoV-2 Infection in Spermatogonia, Leydig and Sertoli Cells. Cells 2020, 9, 920. [Google Scholar] [CrossRef] [Green Version]
- Leung, P.S.; Wong, T.P.; Chung, Y.W.; Chan, H.C. Androgen dependent expression of AT1 receptor and its regulation of anion secretion in rat epididymis. Cell Biol. Int. 2002, 26, 117–122. [Google Scholar] [CrossRef]
- Hamming, I.; Cooper, M.E.; Haagmans, B.L.; Hooper, N.M.; Korstanje, R.; Osterhaus, A.D.M.E.; Timens, W.; Turner, A.J.; Navis, G.; van Goor, H. The emerging role of ACE2 in physiology and disease. J. Pathol. 2007, 212, 1–11. [Google Scholar] [CrossRef]
- Hikmet, F.; Méar, L.; Edvinsson, Å.; Micke, P.; Uhlén, M.; Lindskog, C. The protein expression profile of ACE2 in human tissues. Mol. Syst. Biol. 2020, 16, e9610. [Google Scholar] [CrossRef]
- Kuba, K.; Imai, Y.; Ohto-Nakanishi, T.; Penninger, J.M. Trilogy of ACE2: A peptidase in the renin-angiotensin system, a SARS receptor, and a partner for amino acid transporters. Pharmacol. Ther. 2010, 128, 119–128. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, W.; Yang, L.; You, R. Physiological and pathological regulation of ACE2, the SARS-CoV-2 receptor. Pharmacol. Res. 2020, 157, 104833. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, Y.; Wang, R.; Lu, K.; Tu, M.; Guo, H.; Xie, W.; Qin, Z.; Li, S.; Zhu, P.; et al. Bioinformatic Analysis Reveals That the Reproductive System is Potentially at Risk from SARS-CoV-2. Preprints 2020. [Google Scholar] [CrossRef]
- Valdés, G.; Neves, L.a.A.; Anton, L.; Corthorn, J.; Chacón, C.; Germain, A.M.; Merrill, D.C.; Ferrario, C.M.; Sarao, R.; Penninger, J.; et al. Distribution of angiotensin-(1-7) and ACE2 in human placentas of normal and pathological pregnancies. Placenta 2006, 27, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Lavrentyev, E.N.; Malik, K.U. High glucose-induced Nox1-derived superoxides downregulate PKC-betaII, which subsequently decreases ACE2 expression and ANG(1-7) formation in rat VSMCs. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H106–H118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derby, C.A.; Zilber, S.; Brambilla, D.; Morales, K.H.; McKinlay, J.B. Body mass index, waist circumference and waist to hip ratio and change in sex steroid hormones: The Massachusetts Male Ageing Study. Clin. Endocrinol. 2006, 65, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Oakes, J.M.; Fuchs, R.M.; Gardner, J.D.; Lazartigues, E.; Yue, X. Nicotine and the renin-angiotensin system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R895–R906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.-B.; Wang, Z.-X.; Qiao, Z.-D. The hazardous effects of tobacco smoking on male fertility. Asian J. Androl. 2015, 17, 954–960. [Google Scholar] [CrossRef]
- Köhn, F.M.; Dammshäuser, I.; Neukamm, C.; Renneberg, H.; Siems, W.E.; Schill, W.B.; Aumüller, G. Ultrastructural localization of angiotensin-converting enzyme in ejaculated human spermatozoa. Hum. Reprod. 1998, 13, 604–610. [Google Scholar] [CrossRef]
- Ehlers, M.R.; Riordan, J.F. Angiotensin-converting enzyme: New concepts concerning its biological role. Biochemistry 1989, 28, 5311–5318. [Google Scholar] [CrossRef]
- Miska, W.; Schill, W.B. Enhancement of sperm motility by bradykinin and kinin analogs. Arch. Androl. 1990, 25, 63–67. [Google Scholar] [CrossRef]
- Li, L.-J.; Zhang, F.-B.; Liu, S.-Y.; Tian, Y.-H.; Le, F.; Wang, L.-Y.; Lou, H.-Y.; Xu, X.-R.; Huang, H.-F.; Jin, F. Human sperm devoid of germinal angiotensin-converting enzyme is responsible for total fertilization failure and lower fertilization rates by conventional in vitro fertilization. Biol. Reprod. 2014, 90, 125. [Google Scholar] [CrossRef]
- Tebbs, C.; Pratten, M.K.; Broughton Pipkin, F. Angiotensin II is a growth factor in the peri-implantation rat embryo. J. Anat. 1999, 195, 75–86. [Google Scholar] [CrossRef]
- Gianzo, M.; Urizar-Arenaza, I.; Muñoa-Hoyos, I.; Larreategui, Z.; Garrido, N.; Casis, L.; Irazusta, J.; Subirán, N. Human sperm testicular angiotensin-converting enzyme helps determine human embryo quality. Asian J. Androl. 2018, 20, 498. [Google Scholar] [CrossRef] [PubMed]
- George, A.J.; Thomas, W.G.; Hannan, R.D. The renin-angiotensin system and cancer: Old dog, new tricks. Nat. Rev. Cancer 2010, 10, 745–759. [Google Scholar] [CrossRef]
- Volpe, M.; Musumeci, B.; De Paolis, P.; Savoia, C.; Morganti, A. Angiotensin II AT2 receptor subtype: An uprising frontier in cardiovascular disease? J. Hypertens. 2003, 21, 1429–1443. [Google Scholar] [CrossRef] [PubMed]
- Vinson, G.P.; Puddefoot, J.R.; Ho, M.M.; Barker, S.; Mehta, J.; Saridogan, E.; Djahanbakhch, O. Type 1 angiotensin II receptors in rat and human sperm. J. Endocrinol. 1995, 144, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.S.; Chan, H.C.; Fu, L.X.; Leung, P.Y.; Chew, S.B.; Wong, P.Y. Angiotensin II receptors: Localization of type I and type II in rat epididymides of different developmental stages. J. Membr. Biol. 1997, 157, 97–103. [Google Scholar] [CrossRef]
- Gur, Y.; Breitbart, H.; Lax, Y.; Rubinstein, S.; Zamir, N. Angiotensin II induces acrosomal exocytosis in bovine spermatozoa. Am. J. Physiol. 1998, 275, E87–E93. [Google Scholar] [CrossRef]
- Wennemuth, G.; Babcock, D.F.; Hille, B. Distribution and function of angiotensin II receptors in mouse spermatozoa. Andrologia 1999, 31, 323–325. [Google Scholar] [PubMed]
- Magnan, J.; Regoli, D. Characterization of receptors for angiotensin in the rat vas deferens. Can. J. Physiol. Pharmacol. 1979, 57, 417–423. [Google Scholar] [CrossRef]
- Millan, M.A.; Aguilera, G. Angiotensin II receptors in testes. Endocrinology 1988, 122, 1984–1990. [Google Scholar] [CrossRef]
- Grove, K.L.; Speth, R.C. Rat epididymis contains functional angiotensin II receptors. Endocrinology 1989, 125, 223–230. [Google Scholar] [CrossRef]
- Kitami, Y.; Okura, T.; Marumoto, K.; Wakamiya, R.; Hiwada, K. Differential gene expression and regulation of type-1 angiotensin II receptor subtypes in the rat. Biochem. Biophys. Res. Commun. 1992, 188, 446–452. [Google Scholar] [CrossRef]
- Sum, C.S.; Cheung, W.T. Characterization of contractile response to angiotensin in epididymal rat vas deferens. Pharmacology 1995, 51, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.S.; Chan, H.C.; Fu, L.X.; Zhou, W.L.; Wong, P.Y. Angiotensin II receptors, AT1 and AT2 in the rat epididymis. Immunocytochemical and electrophysiological studies. Biochim. Biophys. Acta 1997, 1357, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Leung, P.S.; Yao, X.Q.; Chan, H.C.; Fu, L.X.; Wong, P.Y. Differential gene expression of angiotensin II receptor subtypes in the epididymides of mature and immature rats. Life Sci. 1998, 62, 461–468. [Google Scholar] [CrossRef]
- Young, D.; Waitches, G.; Birchmeier, C.; Fasano, O.; Wigler, M. Isolation and characterization of a new cellular oncogene encoding a protein with multiple potential transmembrane domains. Cell 1986, 45, 711–719. [Google Scholar] [CrossRef]
- Santos, R.A.S.; Simoes e Silva, A.C.; Maric, C.; Silva, D.M.R.; Machado, R.P.; de Buhr, I.; Heringer-Walther, S.; Pinheiro, S.V.B.; Lopes, M.T.; Bader, M.; et al. Angiotensin-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc. Natl. Acad. Sci. USA 2003, 100, 8258–8263. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.A.S.; Ferreira, A.J. Angiotensin-(1-7) and the renin-angiotensin system. Curr. Opin. Nephrol. Hypertens. 2007, 16, 122–128. [Google Scholar] [CrossRef]
- Walther, T.; Balschun, D.; Voigt, J.P.; Fink, H.; Zuschratter, W.; Birchmeier, C.; Ganten, D.; Bader, M. Sustained long term potentiation and anxiety in mice lacking the Mas protooncogene. J. Biol. Chem. 1998, 273, 11867–11873. [Google Scholar] [CrossRef] [Green Version]
- Leal, M.C.; Pinheiro, S.V.B.; Ferreira, A.J.; Santos, R.A.S.; Bordoni, L.S.; Alenina, N.; Bader, M.; França, L.R. The role of angiotensin-(1-7) receptor Mas in spermatogenesis in mice and rats. J. Anat. 2009, 214, 736–743. [Google Scholar] [CrossRef]
- Dworakowska, D.; Grossman, A.B. Renin-angiotensin system inhibitors in management of hypertension during the COVID-19 pandemic. J. Physiol. Pharmacol. 2020, 71. [Google Scholar] [CrossRef]
- Siems, W.-E.; Maul, B.; Wiesner, B.; Becker, M.; Walther, T.; Rothe, L.; Winkler, A. Effects of kinins on mammalian spermatozoa and the impact of peptidolytic enzymes. Andrologia 2003, 35, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Okeahialam, B.N.; Amadi, K.; Ameh, A.S. Effect of lisnopril, an angiotensin converting enzyme (ACE) inhibitor on spermatogenesis in rats. Arch. Androl. 2006, 52, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Monsees, T.K.; Miska, W.; Schill, W.B. Characterization of kininases in testicular cells. Immunopharmacology 1996, 32, 169–171. [Google Scholar] [CrossRef]
- Saha, L.; Garg, S.K.; Bhargava, V.K.; Mazumdar, S. Role of angiotensin-converting enzyme inhibitor, lisinopril, on spermatozoal functions in rats. Methods Find Exp. Clin. Pharmacol. 2000, 22, 159–162. [Google Scholar]
- Rodríguez-Puertas, R. ACE2 Activators for the Treatment of Covid 19 Patients. J. Med Virol. 2020. [Google Scholar] [CrossRef]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, T.; Katoh, H.; Kato, F.; et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef] [Green Version]
- Antalis, T.M.; Bugge, T.H.; Wu, Q. Membrane-Anchored Serine Proteases in Health and Dsease. In Proteases in Health and Disease-Google Libri; Academic Press: Cambridge, MA, USA, 2011; pp. 1–50. [Google Scholar]
- Choi, S.-Y.; Bertram, S.; Glowacka, I.; Park, Y.W.; Pöhlmann, S. Type II transmembrane serine proteases in cancer and viral infections. Trends Mol. Med. 2009, 15, 303–312. [Google Scholar] [CrossRef]
- Böttcher-Friebertshäuser, E.; Freuer, C.; Sielaff, F.; Schmidt, S.; Eickmann, M.; Uhlendorff, J.; Steinmetzer, T.; Klenk, H.-D.; Garten, W. Cleavage of Influenza Virus Hemagglutinin by Airway Proteases TMPRSS2 and HAT Differs in Subcellular Localization and Susceptibility to Protease Inhibitors. J. Virol. 2010, 84, 5605–5614. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, S.; Nagata, N.; Shirato, K.; Kawase, M.; Takeda, M.; Taguchi, F. Efficient Activation of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein by the Transmembrane Protease TMPRSS2. J. Virol. 2010, 84, 12658–12664. [Google Scholar] [CrossRef] [Green Version]
- Shulla, A.; Heald-Sargent, T.; Subramanya, G.; Zhao, J.; Perlman, S.; Gallagher, T. A Transmembrane Serine Protease Is Linked to the Severe Acute Respiratory Syndrome Coronavirus Receptor and Activates Virus Entry. J. Virol. 2011, 85, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Simmons, G.; Zmora, P.; Gierer, S.; Heurich, A.; Pöhlmann, S. Proteolytic activation of the SARS-coronavirus spike protein: Cutting enzymes at the cutting edge of antiviral research. Antivir. Res. 2013, 100, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Paoloni-Giacobino, A.; Chen, H.; Peitsch, M.C.; Rossier, C.; Antonarakis, S.E. Cloning of the TMPRSS2 Gene, Which Encodes a Novel Serine Protease with Transmembrane, LDLRA, and SRCR Domains and Maps to 21q22.3. Genomics 1997, 44, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Afar, D.E.; Vivanco, I.; Hubert, R.S.; Kuo, J.; Chen, E.; Saffran, D.C.; Raitano, A.B.; Jakobovits, A. Catalytic cleavage of the androgen-regulated TMPRSS2 protease results in its secretion by prostate and prostate cancer epithelia. Cancer Res. 2001, 61, 1686–1692. [Google Scholar]
- Lizio, M.; Abugessaisa, I.; Noguchi, S.; Kondo, A.; Hasegawa, A.; Hon, C.C.; de Hoon, M.; Severin, J.; Oki, S.; Hayashizaki, Y.; et al. Update of the FANTOM web resource: Expansion to provide additional transcriptome atlases. Nucleic Acids Res. 2019, 47, D752–D758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, S.H.; Hirsh, A.; Li, D.C.; Holloway, G.; Chao, J.; Boucher, R.C.; Gabriel, S.E. Regulation of the Epithelial Sodium Channel by Serine Proteases in Human Airways. J. Biol. Chem. 2002, 277, 8338–8345. [Google Scholar] [CrossRef] [Green Version]
- Lin, B.; Ferguson, C.; White, J.T.; Wang, S.; Vessella, R.; True, L.D.; Hood, L.; Nelson, P.S. Prostate-localized and androgen-regulated expression of the membrane-bound serine protease TMPRSS2. Cancer Res. 1999, 59, 4180–4184. [Google Scholar] [PubMed]
- Kim, T.S.; Heinlein, C.; Hackman, R.C.; Nelson, P.S. Phenotypic Analysis of Mice Lacking the Tmprss2-Encoded Protease. Mol. Cell. Biol. 2006, 26, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Ko, C.-J.; Huang, C.-C.; Lin, H.-Y.; Juan, C.-P.; Lan, S.-W.; Shyu, H.-Y.; Wu, S.-R.; Hsiao, P.-W.; Huang, H.-P.; Shun, C.-T.; et al. Androgen-Induced TMPRSS2 Activates Matriptase and Promotes Extracellular Matrix Degradation, Prostate Cancer Cell Invasion, Tumor Growth, and Metastasis. Cancer Res. 2015, 75, 2949–2960. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhou, G.; Sun, Y.; Wang, S.; Yang, J.; Meng, E.; Pan, D.; Li, W.; Zhou, X.; Wang, Y.; et al. Clinical pathology and pathogenesis of severe acute respiratory syndrome. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 2003, 17, 217–221. [Google Scholar]
- Gu, J.; Gong, E.; Zhang, B.; Zheng, J.; Gao, Z.; Zhong, Y.; Zou, W.; Zhan, J.; Wang, S.; Xie, Z.; et al. Multiple organ infection and the pathogenesis of SARS. J. Exp. Med. 2005, 202, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; He, L.; Zhang, Q.; Huang, Z.; Che, X.; Hou, J.; Wang, H.; Shen, H.; Qiu, L.; Li, Z.; et al. Organ distribution of severe acute respiratory syndrome (SARS) associated coronavirus (SARS-CoV) in SARS patients: Implications for pathogenesis and virus transmission pathways. J. Pathol. 2004, 203, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Korteweg, C. Pathology and Pathogenesis of Severe Acute Respiratory Syndrome. Am. J. Pathol. 2007, 170, 1136–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Qi, L.; Chi, X.; Yang, J.; Wei, X.; Gong, E.; Peh, S.; Gu, J. Orchitis: A complication of severe acute respiratory syndrome (SARS). Biol. Reprod. 2006, 74, 410–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, M.; Itoh, M. REVIEW ARTICLE: Patterns of Infiltration of Lymphocytes into the Testis Under Normal and Pathological Conditions in Mice: TESTICULAR INFILTRATION OF LYMPHOCYTES. Am. J. Reprod. Immunol. 2007, 59, 55–61. [Google Scholar] [CrossRef]
- Jacobo, P.; Pérez, C.V.; Theas, M.S.; Guazzone, V.A.; Lustig, L. CD4+ and CD8+ T cells producing Th1 and Th17 cytokines are involved in the pathogenesis of autoimmune orchitis. Reproduction 2011, 141, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.; Schuppe, H.-C. Influence of genital heat stress on semen quality in humans. Andrologia 2007, 39, 203–215. [Google Scholar] [CrossRef]
- Yin, Y.; Wunderink, R.G. MERS, SARS and other coronaviruses as causes of pneumonia: MERS, SARS and coronaviruses. Respirology 2018, 23, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.W.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.A.; Zaki, A.; Fouchier, R.A.M.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Abobaker, A.; Raba, A.A. Does COVID-19 affect male fertility? World J. Urol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Xiao, X.; Aierken, A.; Liao, M.; Hua, J. The ACE2 Expression in Sertoli cells and Germ cells may cause male reproductive disorder after SARS-CoV-2 Infection. J. Cell. Mol. Med. 2020, 24, 9472–9477. [Google Scholar] [CrossRef] [PubMed]
- Stanley, K.E.; Thomas, E.; Leaver, M.; Wells, D. Coronavirus disease (COVID-19) and fertility: Viral host entry protein expression in male and female reproductive tissues. Fertil. Steril. 2020. [Google Scholar] [CrossRef] [PubMed]
- Paoli, D.; Pallotti, F.; Colangelo, S.; Basilico, F.; Mazzuti, L.; Turriziani, O.; Antonelli, G.; Lenzi, A.; Lombardo, F. Study of SARS-CoV-2 in semen and urine samples of a volunteer with positive naso-pharyngeal swab. J. Endocrinol. Investig. 2020. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Wang, Y.; Li, W.; Hu, B.; Chen, G.; Xia, P.; Wang, W.; Li, C.; Diao, F.; Hu, Z.; et al. Absence of 2019 novel coronavirus in semen and testes of COVID-19 patients†. Biol. Reprod. 2020. [Google Scholar] [CrossRef]
- Kayaaslan, B.; Korukluoglu, G.; Hasanoglu, I.; Kalem, A.K.; Eser, F.; Akinci, E.; Guner, R. Investigation of SARS-CoV-2 in Semen of Patients in the Acute Stage of COVID-19 Infection. Urol. Int. 2020, 1–6. [Google Scholar] [CrossRef]
- Pavone, C.; Giammanco, G.M.; Baiamonte, D.; Pinelli, M.; Bonura, C.; Montalbano, M.; Profeta, G.; Curcurù, L.; Bonura, F. Italian males recovering from mild COVID-19 show no evidence of SARS-CoV-2 in semen despite prolonged nasopharyngeal swab positivity. Int. J. Impot. Res. 2020. [Google Scholar] [CrossRef]
- Guo, L.; Zhao, S.; Li, W.; Wang, Y.; Li, L.; Jiang, S.; Ren, W.; Yuan, Q.; Zhang, F.; Kong, F.; et al. Absence of SARS-CoV-2 in Semen of a COVID-19 Patient Cohort. Andrology 2020. [Google Scholar] [CrossRef]
- Li, D.; Jin, M.; Bao, P.; Zhao, W.; Zhang, S. Clinical Characteristics and Results of Semen Tests Among Men With Coronavirus Disease 2019. JAMA Netw. Open 2020, 3, e208292. [Google Scholar] [CrossRef]
- Pan, F.; Xiao, X.; Guo, J.; Song, Y.; Li, H.; Patel, D.P.; Spivak, A.M.; Alukal, J.P.; Zhang, X.; Xiong, C.; et al. No evidence of severe acute respiratory syndrome-coronavirus 2 in semen of males recovering from coronavirus disease 2019. Fertil. Steril. 2020, 113, 1135–1139. [Google Scholar] [CrossRef]
- Yang, M.; Chen, S.; Huang, B.; Zhong, J.-M.; Su, H.; Chen, Y.-J.; Cao, Q.; Ma, L.; He, J.; Li, X.-F.; et al. Pathological Findings in the Testes of COVID-19 Patients: Clinical Implications. Eur. Urol. Focus 2020, 6, 1124–1129. [Google Scholar] [CrossRef]
- La Marca, A.; Busani, S.; Donno, V.; Guaraldi, G.; Ligabue, G.; Girardis, M. Testicular pain as an unusual presentation of COVID-19: A brief review of SARS-CoV-2 and the testis. Reprod. BioMedicine Online 2020. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Thomsen, T.; Sell, N.; Goldsmith, A.J. Abdominal and testicular pain: An atypical presentation of COVID-19. Am. J. Emerg. Med. 2020, 38, 1542.e1–1542.e3. [Google Scholar] [CrossRef] [PubMed]
- Holtmann, N.; Edimiris, P.; Andree, M.; Doehmen, C.; Baston-Buest, D.; Adams, O.; Kruessel, J.-S.; Bielfeld, A.P. Assessment of SARS-CoV-2 in human semen—a cohort study. Fertil. Steril. 2020, 114, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xie, W.; Li, D.; Shi, L.; Ye, G.; Mao, Y.; Xiong, Y.; Sun, H.; Zheng, F.; Chen, Z.; et al. Evaluation of sex-related hormones and semen characteristics in reproductive-aged male COVID-19 patients. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Loveland, K.L.; Klein, B.; Pueschl, D.; Indumathy, S.; Bergmann, M.; Loveland, B.E.; Hedger, M.P.; Schuppe, H.-C. Cytokines in Male Fertility and Reproductive Pathologies: Immunoregulation and Beyond. Front. Endorcrinol. 2017, 8. [Google Scholar] [CrossRef]
- Verma, S.; Saksena, S.; Sadri-Ardekani, H. ACE2 receptor expression in testes: Implications in coronavirus disease 2019 pathogenesis†. Biol. Reprod. 2020, 103, 449–451. [Google Scholar] [CrossRef]
- Corona, G.; Baldi, E.; Isidori, A.M.; Paoli, D.; Pallotti, F.; De Santis, L.; Francavilla, F.; La Vignera, S.; Selice, R.; Caponecchia, L.; et al. SARS-CoV-2 infection, male fertility and sperm cryopreservation: A position statement of the Italian Society of Andrology and Sexual Medicine (SIAMS) (Società Italiana di Andrologia e Medicina della Sessualità). J. Endocrinol. Investig. 2020, 43, 1153–1157. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, H.; Shen, H.; Li, Z.; Geng, J.; Han, H.; Cai, J.; Li, X.; Kang, W.; Weng, D.; et al. The clinical pathology of severe acute respiratory syndrome (SARS): A report from China. J. Pathol. 2003, 200, 282–289. [Google Scholar] [CrossRef]
- Zhou, W.; De Iuliis, G.N.; Dun, M.D.; Nixon, B. Characteristics of the Epididymal Luminal Environment Responsible for Sperm Maturation and Storage. Front. Endorcrinol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- ESHRE Coronavirus Covid-19: ESHRE statement on pregnancy and conception. Available online: https://www.eshre.eu/Press-Room/ESHRE-News (accessed on 20 May 2020).
- ASRM Patient Management and Clinical Recommendations During The Coronavirus (COVID-19) Pandemic. Available online: https://www.asrm.org/news-and-publications/covid-19/statements/patient-management-and-clinical-recommendations-during-the-coronavirus-covid-19-pandemic/ (accessed on 20 May 2020).
- ESHRE ESHRE guidance on recommencing ART treatments. Available online: https://www.eshre.eu/Press-Room/ESHRE-News (accessed on 20 May 2020).
- The ESHRE COVID-19 Working Group; Vermeulen, N.; Ata, B.; Gianaroli, L.; Lundin, K.; Mocanu, E.; Rautakallio-Hokkanen, S.; Tapanainen, J.S.; Veiga, A. A picture of medically assisted reproduction activities during the COVID-19 pandemic in Europe. Hum. Reprod. Open 2020, 2020. [Google Scholar] [CrossRef]
- Vaiarelli, A.; Bulletti, C.; Cimadomo, D.; Borini, A.; Alviggi, C.; Ajossa, S.; Anserini, P.; Gennarelli, G.; Guido, M.; Levi-Setti, P.E.; et al. COVID-19 and ART: The view of the Italian Society of Fertility and Sterility and Reproductive Medicine. Reprod. BioMedicine Online 2020. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Liu, X.; Xiao, M.; Xie, J.; Cao, W.; Liu, Z.; Morse, A.; Xie, Y.; Li, T.; Zhu, L. SARS-CoV-2 is not detectable in the vaginal fluid of women with severe COVID-19 infection. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SART SART Embryology Laboratory Suggestions For COVID-19. Available online: https://www.sart.org/professionals-and-providers/covid-19-resources/embryology-laboratory-suggestions-for-covid-19/ (accessed on 20 May 2020).
- Parmegiani, L.; Accorsi, A.; Bernardi, S.; Arnone, A.; Cognigni, G.E.; Filicori, M. A reliable procedure for decontamination before thawing of human specimens cryostored in liquid nitrogen: Three washes with sterile liquid nitrogen (SLN2). Fertil. Steril. 2012, 98, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Jindal, S.K.; Rawlins, R.G.; Muller, C.H.; Drobnis, E.Z. Guidelines for risk reduction when handling gametes from infectious patients seeking assisted reproductive technologies. Reprod. Biomed. Online 2016, 33, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitken, R.J. COVID-19 and human spermatozoa—Potential risks for infertility and sexual transmission? Andrology 2020. [Google Scholar] [CrossRef] [PubMed]
- Millsop, J.W.; Heller, M.M.; Eliason, M.J.; Murase, J.E. Dermatological medication effects on male fertility: Medication effects on male fertility. Dermatol. Ther. 2013, 26, 337–346. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| LOCATION | ACE [44,57,58] | ACE2 [21,59] | AT1R [55,60,61] | AT2R [55,61,62] | MAS [53,63] | TMPRSS2 [64,65,66,67] | REFERENCES | ||
|---|---|---|---|---|---|---|---|---|---|
| sACE | tACE | ||||||||
| PROSTATE | X | X | X | X | [44,55,57,58,60,61,62,64,65,66,67,68] | ||||
| VAS DEFERENS | X | [55,60,61] | |||||||
| EPIDIDYMIS | X | X | X | X | [44,55,57,58,60,61,62,64,65,66,67,68] | ||||
| SERTOLI CELLS | X | X | [53,59,63,67] | ||||||
| LEYDIG CELLS | X | X | X | X | X | [44,53,55,57,58,59,60,61,63,64,65,66,67] | |||
| SEMINAL PLASMA | X | X | [44,57,58,64,65,66,67] | ||||||
| SPERMATOGONIA | X | X | X | [53,59,63,64,65,66,67] | |||||
| SPERMATOCYTES | X | [53,63] | |||||||
| SPERMATIDS | X | X | X | X | [44,53,55,57,58,60,61,63,64,65,66,67] | ||||
| S P E R M A T O Z O A | ACROSOMAL REGION | X | X | [44,53,57,58,63] | |||||
| EQUATORIAL SEGMENT | X | X | [44,55,57,58,61,62] | ||||||
| POST ACROSOMAL REGION | X | X | [44,55,57,58,61,62] | ||||||
| MID PIECE | X | [44,57,58] | |||||||
| FLAGELLUM | X | X | [44,53,57,58,63] | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascolo, L.; Zito, G.; Zupin, L.; Luppi, S.; Giolo, E.; Martinelli, M.; De Rocco, D.; Crovella, S.; Ricci, G. Renin Angiotensin System, COVID-19 and Male Fertility: Any Risk for Conceiving? Microorganisms 2020, 8, 1492. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101492
Pascolo L, Zito G, Zupin L, Luppi S, Giolo E, Martinelli M, De Rocco D, Crovella S, Ricci G. Renin Angiotensin System, COVID-19 and Male Fertility: Any Risk for Conceiving? Microorganisms. 2020; 8(10):1492. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101492
Chicago/Turabian StylePascolo, Lorella, Gabriella Zito, Luisa Zupin, Stefania Luppi, Elena Giolo, Monica Martinelli, Daniela De Rocco, Sergio Crovella, and Giuseppe Ricci. 2020. "Renin Angiotensin System, COVID-19 and Male Fertility: Any Risk for Conceiving?" Microorganisms 8, no. 10: 1492. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101492