Bacteria-Derived Hemolysis-Related Genes Widely Exist in Scuticociliates

Abstract
:1. Introduction
2. Methods
2.1. Ciliate Culture and Identification
2.2. DNA and RNA Sample Preparation and Sequencing
2.3. U. marinum Genome Assembly and Gene Prediction
2.4. Identification of HGT Genes in U. marinum
2.5. Homologs of U. marinum HGT Genes in Other Scuticociliates
3. Results and Discussion
3.1. The U. marinum MAC Genome
3.2. U. marinum Acquired Hemolysis-Related Genes through HGT
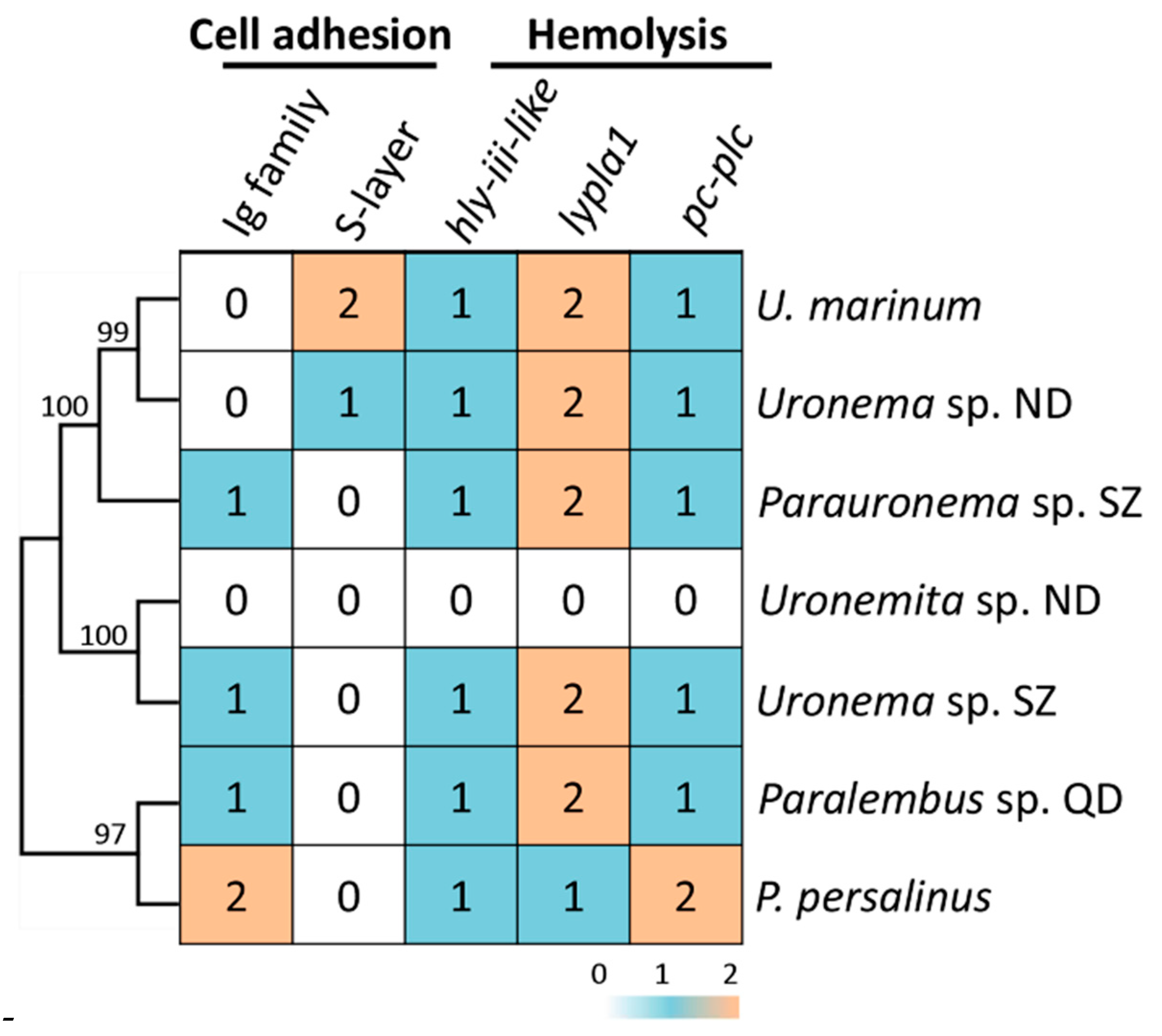
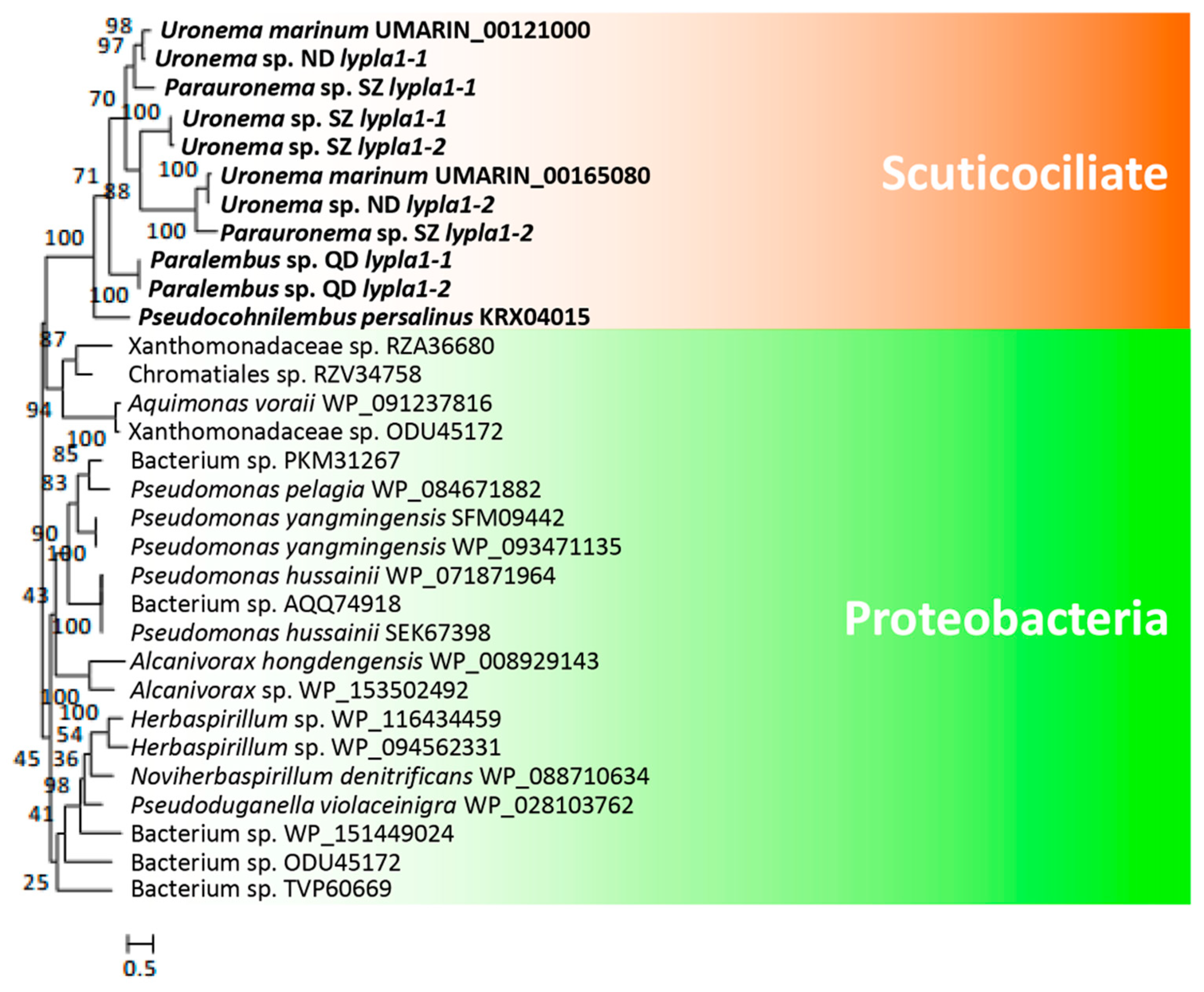
3.3. Hemolysis-Related Genes Are Widely Distributed in Scuticociliates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Iglesias:, R.; Parama, A.; Alvarez, M.F.; Leiro, J.; Fernandez, J.; Sammartin, M.L. Philasterides dicentrarchi (Ciliophora, Scuticociliatida) as the causative agent of scuticociliatosis in farmed turbot Scophthalmus maximus in galicia (nw spain). Dis. Aquat. Organ. 2001, 46, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.M.; Cho, J.B.; Kim, S.K.; Nam, Y.K.; Kim, K.H. Occurrence of scuticociliatosis in olive flounder Paralichthys olivaceus by Phiasterides dicentrarchi (Ciliophora: Scuticociliatida). Dis. Aquat. Organ. 2004, 62, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, I.S.; Al-Marzouk, A.; James, C.M.; Almatar, S.; Al-Gharabally, H. Scuticociliatosis-associated mortalities and histopathology of natural infection in cultured silver pomfret (pampus argenteus euphrasen) in kuwait. Aquaculture 2007, 262, 202–210. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Scuticociliatosis and its recent prophylactic measures in aquaculture with special reference to South Korea taxonomy, diversity and diagnosis of scuticociliatosis: Part i control strategies of scuticociliatosis: Part ii. Fish Shellfish Immun. 2010, 29, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Parama, A.; Iglesias, R.; Alvarez, M.F.; Leiro, J.; Ubeira, F.M.; Sanmartin, M.L. Cysteine proteinase activities in the fish pathogen Philasterides dicentrarchi (Ciliophora: Scuticociliatida). Parasitology 2004, 128, 541–548. [Google Scholar] [CrossRef]
- Lee, E.H.; Kim, C.S.; Cho, J.B.; Ahn, K.J.; Kim, K.H. Measurement of protease activity of live Uronema marinum (Ciliata: Scuticociliatida) by fluorescence polarization. Dis. Aquat. Organ. 2003, 54, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Wang, G.Y.; Cheng, J.; Tian, M.; Pan, X.M.; Warren, A.; Jiang, C.; Yuan, D.; Miao, W. Genome of the facultative scuticociliatosis pathogen Pseudocohnilembus persalinus provides insight into its virulence through horizontal gene transfer. Sci. Rep. 2015, 5, 15470. [Google Scholar] [CrossRef] [Green Version]
- Cheung, P.J.; Nigrelli, R.F.; Ruggieri, G.D. Studies on the morphology of Uronema-marinum Dujardin (Ciliatea, Uronematidae) with a description of the histopathology of the infection in marine fishes. J. Fish Dis. 1980, 3, 295–303. [Google Scholar] [CrossRef]
- Zheng, W.B.; Gao, F.; Warren, A. High-density cultivation of the marine ciliate Uronema marinum (Ciliophora, Oligohymenophorea) in axenic medium. Acta Protozool. 2015, 54, 325–330. [Google Scholar]
- Wang, C.N.; Bourland, W.A.; Mu, W.J.; Pan, X.M. Transcriptome analysis on chlorpyrifos detoxification in Uronema marinum (Ciliophora, Oligohymenophorea). Environ. Sci. Pollut. Res. 2018, 25, 33402–33414. [Google Scholar] [CrossRef]
- Fernandezgaliano, D. The ammoniacal silver carbonate method as a general procedure in the study of protozoa from sewage (and other) waters. Water Res. 1994, 28, 495–496. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16s-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Dykova, I.; Tyml, T.; Kostka, M.; Peckova, H. Strains of Uronema marinum (Scuticociliatia) co-isolated with amoebae of the genus Neoparamoeba. Dis. Aquat. Organ. 2010, 89, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Lu, Y.M.; Feng, J.M.; Yuan, D.X.; Tian, M.; Chang, Y.; Fu, C.J.; Wang, G.Y.; Zeng, H.H.; Miao, W. Tetrahymena functional genomics database (TetraFGD): An integrated resource for Tetrahymena functional genomics. Database-Oxf. 2013, 2013, bat008. [Google Scholar] [CrossRef]
- Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From fastq data to high confidence variant calls: The genome analysis toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. Tophat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Wang, G.Y.; Xiong, J.; Jiang, C.Q.; Miao, W. Exploration of genetic variations through single-cell whole-genome sequencing in the model ciliate Tetrahymena thermophila. J. Eukaryot. Microbiol. 2019, 66, 954–965. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with burrows-wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.D.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with Tophat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Delcher, A.L.; Mount, S.M.; Wortman, J.R.; Smith, R.K.; Hannick, L.I.; Maiti, R.; Ronning, C.M.; Rusch, D.B.; Town, C.D.; et al. Improving the Arabidopsis genome annotation using maximal transcript alignment assemblies. Nucleic Acids Res. 2003, 31, 5654–5666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majoros, W.H.; Pertea, M.; Salzberg, S.L. Tigrscan and Glimmerhmm: Two open source ab initio eukaryotic gene-finders. Bioinformatics 2004, 20, 2878–2879. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Salzberg, S.L.; Zhu, W.; Pertea, M.; Allen, J.E.; Orvis, J.; White, O.; Buell, C.R.; Wortman, J.R. Automated eukaryotic gene structure annotation using evidencemodeler and the program to assemble spliced alignments. Genome Biol. 2008, 9, R7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. Interproscan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Stoeckert, C.J.; Roos, D.S. Orthomcl: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. Blast plus: Architecture and applications. BMC Bioinform. 2009, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. Fasttree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Li, D.H.; Liu, C.M.; Luo, R.B.; Sadakane, K.; Lam, T.W. Megahit: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Diekhans, M.; Baertsch, R.; Haussler, D. Using native and syntenically mapped cDNA alignments to improve de novo gene finding. Bioinformatics 2008, 24, 637–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birney, E.; Clamp, M.; Durbin, R. Genewise and genomewise. Genome Res. 2004, 14, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Stanke, M.; Morgenstern, B. Augustus: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aury, J.M.; Jaillon, O.; Duret, L.; Noel, B.; Jubin, C.; Porcel, B.M.; Segurens, B.; Daubin, V.; Anthouard, V.; Aiach, N.; et al. Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia. Nature 2006, 444, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Doerder, F.P.; Deak, J.C.; Lief, J.H. Rate of phenotypic assortment in Tetrahymena thermophila. Dev. Genet. 1992, 13, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Doak, T.G.; Lipps, H.J.; Wang, J.M.; Swart, E.C.; Chang, W.J. Copy number variations of 11 macronuclear chromosomes and their gene expression in Oxytricha trifallax. Gene 2012, 505, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Eisen, J.A.; Coyne, R.S.; Wu, M.; Wu, D.; Thiagarajan, M.; Wortman, J.R.; Badger, J.H.; Ren, Q.; Amedeo, P.; Jones, K.M.; et al. Macronuclear genome sequence of the ciliate Tetrahymena thermophila, a model eukaryote. PLoS Biol. 2006, 4, e286. [Google Scholar] [CrossRef]
- Coyne, R.; Hannick, L.; Shanmugam, D.; Hostetler, J.; Brami, D.; Joardar, V.; Johnson, J.; Radune, D.; Singh, I.; Badger, J.; et al. Comparative genomics of the pathogenic ciliate Ichthyophthirius multifiliis, its free-living relatives and a host species provide insights into adoption of a parasitic lifestyle and prospects for disease control. Genome Biol. 2011, 12, R100. [Google Scholar] [CrossRef] [Green Version]
- McGrath, C.L.; Gout, J.F.; Doak, T.G.; Yanagi, A.; Lynch, M. Insights into three whole-genome duplications gleaned from the Paramecium caudatum genome sequence. Genetics 2014, 197, 1417–1482. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Katz, L.A.; Song, W.B. Insights into the phylogenetic and taxonomy of philasterid ciliates (Protozoa, Ciliophora, Scuticociliatia) based on analyses of multiple molecular markers. Mol. Phylogenet. Evol. 2012, 64, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.M.; Jiang, C.Q.; Warren, A.; Tian, M.; Cheng, J.; Liu, G.L.; Xiong, J.; Miao, W. Phylogenomic analyses reveal subclass Scuticociliatia as the sister group of subclass Hymenostomatia within class Oligohymenophorea. Mol. Phylogenet. Evol. 2015, 90, 104–111. [Google Scholar] [CrossRef]
- Stow, J.L.; Condon, N.D. The cell surface environment for pathogen recognition and entry. Clin. Transl. Immunol. 2016, 5, e71. [Google Scholar] [CrossRef] [PubMed]
- Hynonen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biot. 2013, 97, 5225–5243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.J.; Boyd, J.M.; Horswill, A.R.; Nauseef, W.M. Phosphatidylinositol-specific phospholipase C contributes to survival of Staphylococcus aureus USA300 in human blood and neutrophils. Infect. Immun. 2014, 82, 1559–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baida, G.E.; Kuzmin, N.P. Mechanism of action of hemolysin III from Bacillus cereus. BBA-Biomembranes 1996, 1284, 122–124. [Google Scholar] [CrossRef] [Green Version]
- Kalliomaki, M.A.; Walker, W.A. Physiologic and pathologic interactions of bacteria with gastrointestinal epithelium. Gastroenterol. Clin. N. 2005, 34, 383–399. [Google Scholar] [CrossRef]
- Ramarao, N.; Sanchis, V. The pore-forming haemolysins of Bacillus cereus: A review. Toxins 2013, 5, 1119–1139. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | U. marinum | P. persalinus | T. thermophila | I. multifiliis | P. tetraurelia |
|---|---|---|---|---|---|
| Subclass | Scuticociliatia | Scuticociliatia | Oligohymenophorea | Oligohymenophorea | Oligohymenophorea |
| Genome size (Mb) | 86.8 | 55.5 | 103.0 | 47.8 | 72.1 |
| N50 (Kb) | 470 | 368 | 521 | 66 | 413 |
| Scaffold number | 403 | 288 | 1148 | 1375 | 697 |
| Longest scaffold (Mb) | 2.48 | 2.0 | 2.2 | 0.4 | 1.0 |
| Sequencing method/platform | Nanopore/MGI | Illumina | Sanger | Sanger/454 | Sanger |
| Average guanine-cytosine (GC) content | 18% | 19% | 22% | 16% | 28% |
| Assembled chromosome number | 218 (54%) | 0 (0%) | 129 (11.2%) | 0 (0%) | 8(1%) |
| Completeness | 84.8% | 79.5% | 85.1% | 67.0% | 85.5% |
| Gene number | 24,582 | 13,186 | 26,460 | 8062 | 39,642 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Chen, K.; Jiang, C.; Yang, W.; Gu, S.; Wang, G.; Lu, Y.; Miao, W.; Xiong, J. Bacteria-Derived Hemolysis-Related Genes Widely Exist in Scuticociliates. Microorganisms 2020, 8, 1838. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8111838
Zhang J, Chen K, Jiang C, Yang W, Gu S, Wang G, Lu Y, Miao W, Xiong J. Bacteria-Derived Hemolysis-Related Genes Widely Exist in Scuticociliates. Microorganisms. 2020; 8(11):1838. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8111838
Chicago/Turabian StyleZhang, Jing, Kai Chen, Chuanqi Jiang, Wentao Yang, Siyu Gu, Guangying Wang, Yishan Lu, Wei Miao, and Jie Xiong. 2020. "Bacteria-Derived Hemolysis-Related Genes Widely Exist in Scuticociliates" Microorganisms 8, no. 11: 1838. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8111838