Analysis of Complete Genome Sequence of Acinetobacter baumannii Strain ATCC 19606 Reveals Novel Mobile Genetic Elements and Novel Prophage

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibiotic Resistance Profile and Resistance to Mercury
2.2. Static Biofilm Assay and Quantification of Biofilm Production Using Fluorescence Microscopy
2.3. Genome Sequencing
2.4. Phylogenetics and Sequence Analysis
2.5. Data Availability
2.6. Phage Induction and Isolation
2.7. Phage Concentration and Preparation for Transmission Electron Microscopy (TEM)
2.8. Phage Host Range
3. Result
3.1. Antibiotic Resistance Profile
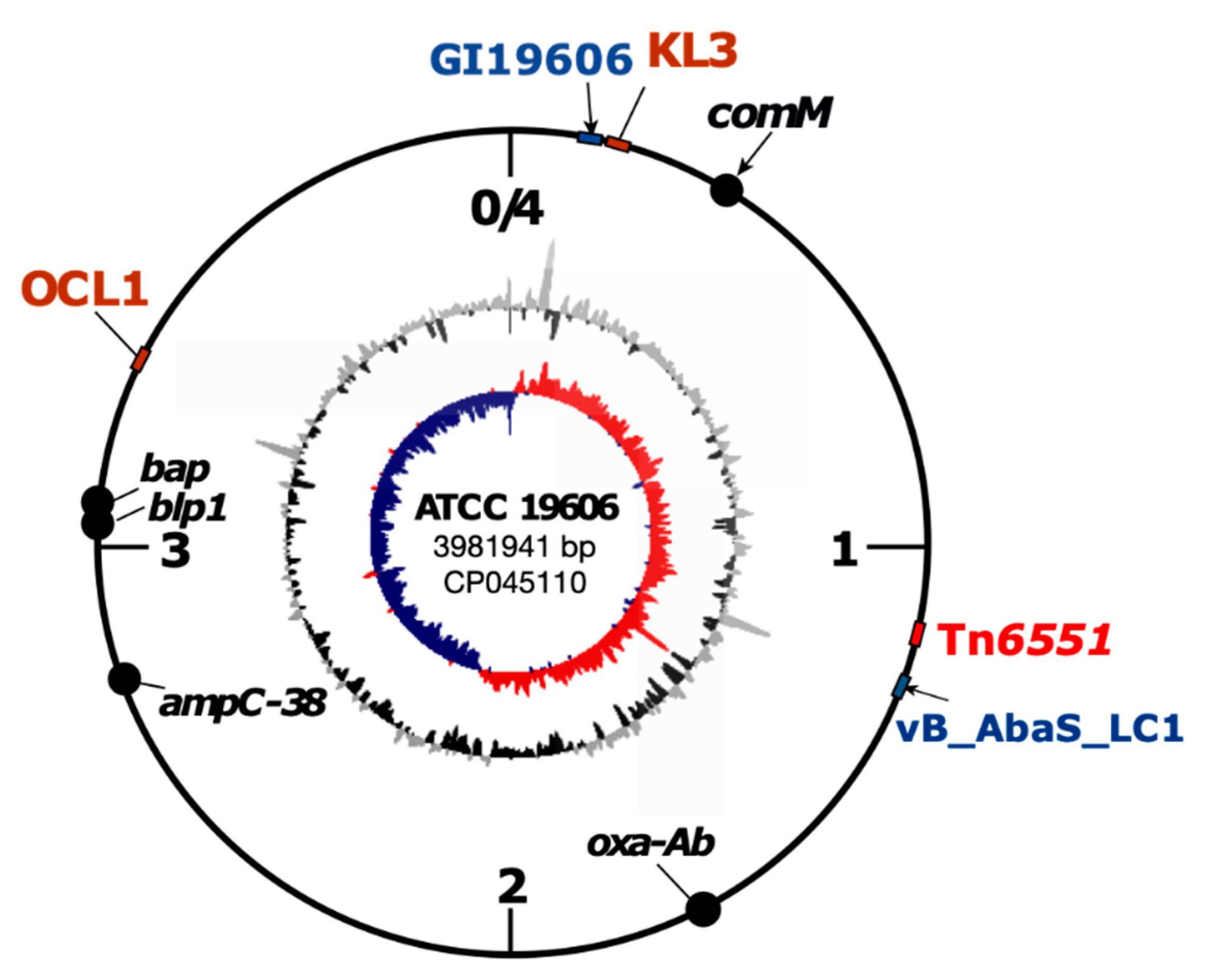
3.2. Complete Genome Sequence of ATCC 19606, Antibiotic Resistance Gene and Its Genomic Context
3.3. Tn6551, a Novel Mercury/Cadmium Transposon Found in ATCC 19606
3.4. ATCC 19606 Carries Two Cryptic Plasmids
3.5. ATCC 19606 Carries a Novel Prophage, vB_AbaS_LC1
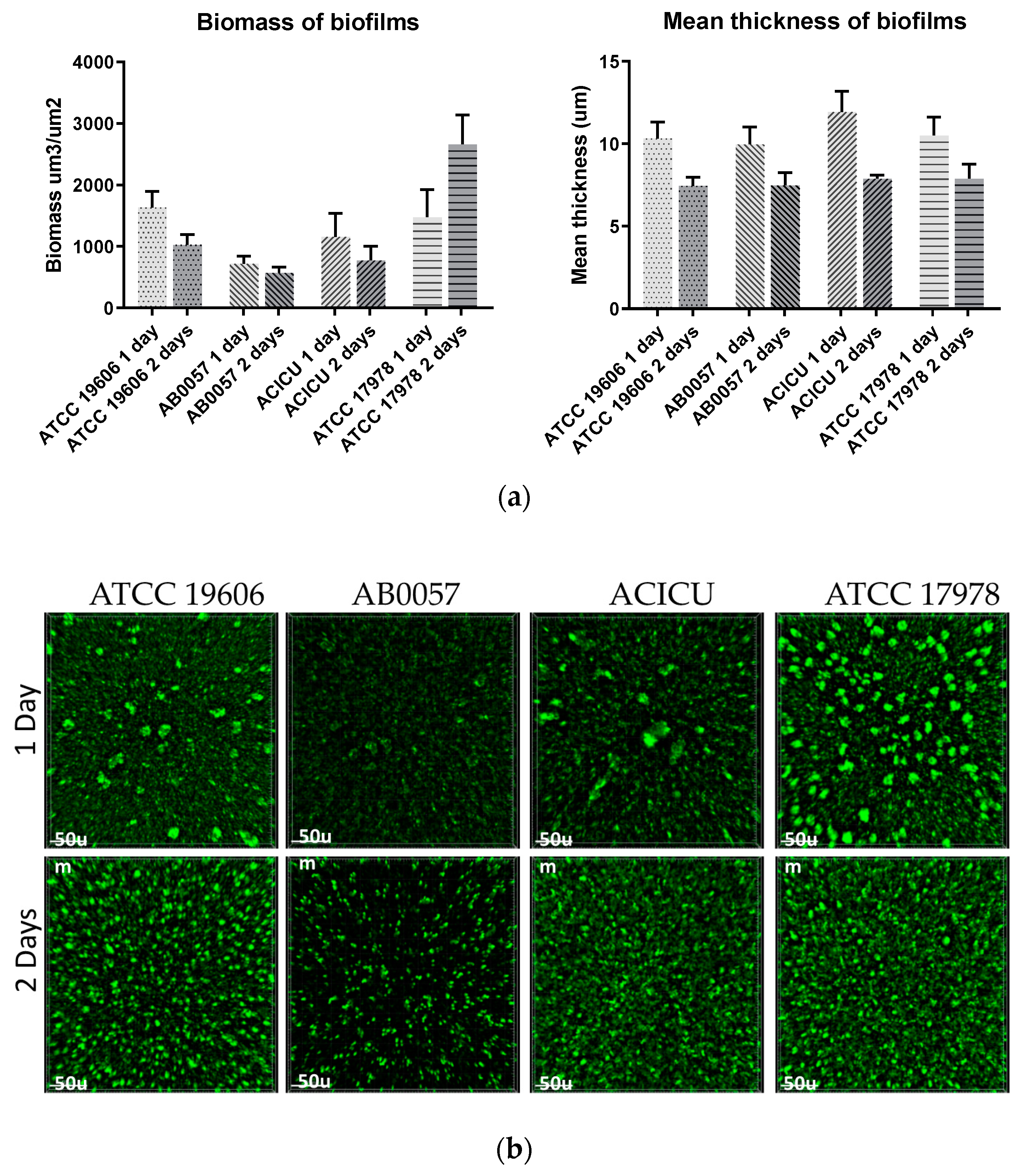
3.6. Biofilm Formation
3.7. Surface Polysaccharide Loci Types
3.8. Comparative Analysis of ST52 Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Hamidian, M.; Hall, R.M. The AbaR antibiotic resistance islands found in Acinetobacter baumannii global clone 1—Structure, origin and evolution. Drug Resist. Updates Rev. Comment. Antimicrob. Anticancer Chemother. 2018, 41, 26–39. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, E.; Del Franco, M.; Martinucci, M.; Roscetto, E.; Zarrilli, R.; Di Nocera, P.P. Biofilm-associated proteins: News from Acinetobacter. BMC Genom. 2015, 16, 933. [Google Scholar] [CrossRef] [Green Version]
- Hugh, R.; Reese, R. A comparison of 120 strains of Bacterium anitratum Schaub and Hauber with the type strain of this species. Int. J. Syst. Evol. Microbiol. 1968, 18, 207–229. [Google Scholar] [CrossRef] [Green Version]
- Harris, G.; Lee, R.K.; Lam, C.K.; Kanzaki, G.; Patel, G.B.; Xu, H.H.; Chen, W. A mouse model of Acinetobacter baumannii-associated pneumonia using a clinically isolated hypervirulent strain. Antimicrob. Agents Chemother. 2013, 57, 3601–3613. [Google Scholar] [CrossRef] [Green Version]
- Pachón-Ibáñez, M.E.; Docobo-Pérez, F.; López-Rojas, R.; Domínguez-Herrera, J.; Jiménez-Mejias, M.E.; García-Curiel, A.; Pichardo, C.; Jiménez, L.; Pachón, J. Efficacy of rifampin and its combinations with imipenem, sulbactam, and colistin in experimental models of infection caused by imipenem-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 1165–1172. [Google Scholar] [CrossRef] [Green Version]
- Davenport, K.W.; Daligault, H.E.; Minogue, T.D.; Bruce, D.C.; Chain, P.S.; Coyne, S.R.; Jaissle, J.G.; Koroleva, G.I.; Ladner, J.T.; Li, P.E.; et al. Draft Genome Assembly of Acinetobacter baumannii ATCC 19606. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Lu, J.; Zhao, J.; Zhang, X.; Yu, H.H.; Velkov, T.; Li, J. Complete genome sequence and genome-scale metabolic modelling of Acinetobacter baumannii type strain ATCC 19606. Int. J. Med. Microbiol. 2020, 310, 151412. [Google Scholar] [CrossRef]
- Tsubouchi, T.; Suzuki, M.; Niki, M.; Oinuma, K.I.; Niki, M.; Kakeya, H.; Kaneko, Y. Complete Genome Sequence of Acinetobacter baumannii ATCC 19606(T), a Model Strain of Pathogenic Bacteria Causing Nosocomial Infection. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; 22nd Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Hamidian, M.; Hall, R.M. Genetic structure of four plasmids found in Acinetobacter baumannii isolate D36 belonging to lineage 2 of global clone 1. PLoS ONE 2018, 13, e0204357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47. [Google Scholar] [CrossRef] [PubMed]
- Gaio, D.; To, J.; Liu, M.; Monahan, L.; Anantanawat, K.; Darling, A.E. Hackflex: Low cost Illumina sequencing library construction for high sample counts. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Carver, T.J.; Rutherford, K.M.; Berriman, M.; Rajandream, M.A.; Barrell, B.G.; Parkhill, J. ACT: The Artemis Comparison Tool. Bioinformatics 2005, 21, 3422–3423. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M.; Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of bacteriophages by double agar overlay plaque assay. Methods Mol. Biol. 2009, 501, 69–76. [Google Scholar] [CrossRef]
- Hamidian, M.; Hall, R.M. Acinetobacter baumannii ATCC 19606 Carries GIsul2 in a Genomic Island Located in the Chromosome. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Hawkey, J.; Wick, R.; Holt, K.E.; Hall, R.M. Evolution of a clade of Acinetobacter baumannii global clone 1, lineage 1 via acquisition of carbapenem- and aminoglycoside-resistance genes and dispersion of ISAba1. Microb. Genom. 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Hamidian, M.; Hall, R.M. Resistance to third-generation cephalosporins in Acinetobacter baumannii due to horizontal transfer of a chromosomal segment containing ISAba1-ampC. J. Antimicrob. Chemother. 2014, 69, 2865–2866. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, G.A.; Hall, R.M. The tet39 Determinant and the msrE-mphE Genes in Acinetobacter Plasmids Are Each Part of Discrete Modules Flanked by Inversely Oriented pdif (XerC-XerD) Sites. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Andrea, M.M.; Giani, T.; D’Arezzo, S.; Capone, A.; Petrosillo, N.; Visca, P.; Luzzaro, F.; Rossolini, G.M. Characterization of pABVA01, a plasmid encoding the OXA-24 carbapenemase from Italian isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3528–3533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merino, M.; Acosta, J.; Poza, M.; Sanz, F.; Beceiro, A.; Chaves, F.; Bou, G. OXA-24 carbapenemase gene flanked by XerC/XerD-like recombination sites in different plasmids from different Acinetobacter species isolated during a nosocomial outbreak. Antimicrob. Agents Chemother. 2010, 54, 2724–2727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorsey, C.W.; Tomaras, A.P.; Actis, L.A. Sequence and organization of pMAC, an Acinetobacter baumannii plasmid harboring genes involved in organic peroxide resistance. Plasmid 2006, 56, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, A.P.; Dorsey, C.W.; Edelmann, R.E.; Actis, L.A. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: Involvement of a novel chaperone-usher pili assembly system. Microbiology 2003, 149, 3473–3484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef]
- Kenyon, J.J.; Hall, R.M. Variation in the complex carbohydrate biosynthesis loci of Acinetobacter baumannii genomes. PLoS ONE 2013, 8, e62160. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Nigro, S.J.; Hall, R.M. Variants of the gentamicin and tobramycin resistance plasmid pRAY are widely distributed in Acinetobacter. J. Antimicrob. Chemother. 2012, 67, 2833–2836. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Hall, R.M. pACICU2 is a conjugative plasmid of Acinetobacter carrying the aminoglycoside resistance transposon TnaphA6. J. Antimicrob. Chemother. 2014, 69, 1146–1148. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Holt, K.E.; Pickard, D.; Dougan, G.; Hall, R.M. A GC1 Acinetobacter baumannii isolate carrying AbaR3 and the aminoglycoside resistance transposon TnaphA6 in a conjugative plasmid. J. Antimicrob. Chemother. 2014, 69, 955–958. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Kenyon, J.J.; Holt, K.E.; Pickard, D.; Hall, R.M. A conjugative plasmid carrying the carbapenem resistance gene blaOXA-23 in AbaR4 in an extensively resistant GC1 Acinetobacter baumannii isolate. J. Antimicrob. Chemother. 2014, 69, 2625–2628. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, L.A.; Ramage, E.; Weiss, E.J.; Radey, M.; Hayden, H.S.; Held, K.G.; Huse, H.K.; Zurawski, D.V.; Brittnacher, M.J.; Manoil, C. Resources for Genetic and Genomic Analysis of Emerging Pathogen Acinetobacter baumannii. J. Bacteriol. 2015, 197, 2027–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, K.; Kenyon, J.J.; Hamidian, M.; Schultz, M.B.; Pickard, D.J.; Dougan, G.; Hall, R. Five decades of genome evolution in the globally distributed, extensively antibiotic-resistant Acinetobacter baumannii global clone 1. Microb. Genom. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Breisch, J.; Waclawska, I.; Averhoff, B. Identification and characterization of a carnitine transporter in Acinetobacter baumannii. MicrobiologyOpen 2019, 8, e00752. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; St Michael, F.; Cox, A.D.; et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Unno, Y.; Kawakami, S.; Ubagai, T.; Ono, Y. Virulence characteristics of Acinetobacter baumannii clinical isolates vary with the expression levels of omps. J. Med. Microbiol 2017, 66, 203–212. [Google Scholar] [CrossRef]
- Zeidler, S.; Müller, V. The role of compatible solutes in desiccation resistance of Acinetobacter baumannii. MicrobiologyOpen 2019, 8, e00740. [Google Scholar] [CrossRef] [Green Version]
- Zimbler, D.L.; Arivett, B.A.; Beckett, A.C.; Menke, S.M.; Actis, L.A. Functional features of TonB energy transduction systems of Acinetobacter baumannii. Infect. Immun. 2013, 81, 3382–3394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, G.A.; Hall, R.M. Mobilisation of a small Acinetobacter plasmid carrying an oriT transfer origin by conjugative RepAci6 plasmids. Plasmid 2019, 103, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Hamidian, M.; Ambrose, S.J.; Hall, R.M. A large conjugative Acinetobacter baumannii plasmid carrying the sul2 sulphonamide and strAB streptomycin resistance genes. Plasmid 2016, 87–88, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Leungtongkam, U.; Thummeepak, R.; Tasanapak, K.; Sitthisak, S. Acquisition and transfer of antibiotic resistance genes in association with conjugative plasmid or class 1 integrons of Acinetobacter baumannii. PLoS ONE 2018, 13, e0208468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, F.; Shen, C.; Zheng, X.; Liu, Y.; Chen, H.; Zhong, L.; Liang, Y.; Liao, K.; Xia, Y.; Tian, G.B.; et al. Identification of a Novel Plasmid Carrying mcr-4.3 in an Acinetobacter baumannii Strain in China. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Douraghi, M.; Kenyon, J.J.; Aris, P.; Asadian, M.; Ghourchian, S.; Hamidian, M. Accumulation of Antibiotic Resistance Genes in Carbapenem-Resistant Acinetobacter baumannii Isolates Belonging to Lineage 2, Global Clone 1, from Outbreaks in 2012-2013 at a Tehran Burns Hospital. mSphere 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- García-Quintanilla, M.; Pulido, M.R.; López-Rojas, R.; Pachón, J.; McConnell, M.J. Emerging therapies for multidrug resistant Acinetobacter baumannii. Trends Microbiol. 2013, 21, 157–163. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Family | Antibiotic | Growth Inhibition Zone (Diameter in mm) 1 |
|---|---|---|
| ß-lactams | Ampicillin | 0 |
| Cefotaxime | 15 | |
| Ceftazidime | 18 | |
| Ceftriaxone | 17 | |
| Imipenem | 19 | |
| Meropenem | 21 | |
| Ampicillin/Sulbactam | 19 | |
| Aminoglycosides | Gentamicin | 14 |
| Kanamycin | 15 | |
| Neomycin | 14 | |
| Tobramycin | 13 | |
| Netilmicin | 16 | |
| Amikacin | 15 | |
| Streptomycin | 7 | |
| Spectinomycin | 0 | |
| Fluoroquinolones | Nalidixic acid | 13 |
| Ciprofloxacin | 16 | |
| Tetracyclines | Tetracycline | 15 |
| Chloramphenicol | Chloramphenicol | 0 |
| RNA Synthesis inhibitor | Rifampicin | 14 |
| Folic acid Synthesis Inhibitors | Trimethoprim | 12 |
| Sulfamethoxazole | 0 |
| ATCC 19606 Genomes | Length | GC% | Reading Frames | Sequencing Technology | Assembly Program | GenBank no. | Ref. |
|---|---|---|---|---|---|---|---|
| Chromosome | 3,981,941 | 39.1 | 3727 | Illumina MiSeq & MinION | Unicycler | CP045110 | This study |
| p1ATCC 19606 | 7655 | 33.3 | 14 | CP045108 | |||
| p2ATCC19606 (pMAC) | 9540 | 34.6 | 14 | CP045109 | |||
| Chromosome | 3,980,848 | 39.1 | 3709 | PacBio | SMRT | CP046654 | [9] |
| pMAC | 9540 | 34.6 | 14 | plasmidSPAdes | CP046655 | ||
| Chromosome | 3,978,812 | Illumina MiSeq & MinION | Unicycler | AP022836 | [10] | ||
| pATCC 19606-1 | 9408 | 34.6 | 14 | AP022837 | |||
| pATCC 19606-2 | 7631 | 33.3 | 12 | AP022838 |
| Genus/Species | Strain | Date | Country | Source | ST (IP) | Tn | TSD 5′-3′ 1 | GenBank no. |
|---|---|---|---|---|---|---|---|---|
| A. baumannii | ATCC 19606 | <1948 | USA | Urine | 52 | 6551 | ATTTTTTT | CP045110 |
| A. baumannii | ab736 | 2015 | USA | Blood | 52 | 6551 | ATTTTTTT | CP015121 |
| A. baumannii | AB031 | 2010 | Canada | Blood | 638 | 6551 | ATTTTTTT | CP009256 |
| A. pittii | BEC1-S18-ESBL-01 | 2018 | Japan | Water 2 | 457 | 6551 | ATTTTTTT | AP022302 |
| A. baumannii | NCTC7364 | 2014 | UK | nk 3 | 494 | 6551-v14 | ATTTTTTT | LT605059 |
| A. indicus | Yy_1 | 2019 | China | Soil | - | 6551-v1 | ATTTTTTT | CP039031 |
| A. junii | WCHAJ59 | 2015 | China | Sewage | - | 6551-v1 | ATTTTTTT | CP028800 |
| A. nosocomialis | AC1530 | 2015 | Malaysia | Blood | - | 6551-v1 | ATAATTAA | CP045560 |
| Neisseria brasiliensis | N.177.16 | 2016 | Brazil | Ulcer exudate | - | 6551-v1 | ATGTATTT | CP046027 |
| Predicted Function | Biological Function | Protein Id 1 | E-Value | Tool |
|---|---|---|---|---|
| Hypothetical protein | Unknown function | QFQ04715.1 | 4.00 × 10−177 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04716.1 | 5.00 × 10−114 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04717.1 | 4.00 × 10−53 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04718.1 | 0 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04719.1 | 1.00 × 10−42 | BLASTp |
| RecA protein; Recombination, Radio-resistance, DNA repair, ATPase, DNA binding; HET: AGS; 2.5A {Deinococcus radiodurans} SCOP: d.48.1.1, c.37.1.11 | Recombination | QFQ04720.1 | 1.30 × 10−10 | HHpred |
| Hypothetical protein | Unknown function | QFQ04721.1 | 1.00 × 10−73 | BLASTp |
| 50S ribosomal protein L2, 50S; Ribosome, bacterial ribosome, proline-rich antimicrobial; | Protein synthesis | QFQ04722.1 | 0.3 | HHpred |
| Hypothetical protein | Unknown function | QFQ04723.1 | 1.00 × 10−102 | BLASTp |
| Helix-turn-helix domain-containing protein | Lysogeny | QFQ04724.1 | 0 | BLASTp |
| Cro/Cl family transcriptional regulator | Lysogeny | QFQ04725.1 | 7.00 × 10−54 | BLASTp |
| XRE family transcriptional regulator | Lysogeny | QFQ04726.1 | 4.00 × 10−117 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04727.1 | 7.00 × 10−64 | BLASTp |
| DNA cytosine methyltransferase | Defence | QFQ04728.1 | 0 | BLASTp |
| YdaU family protein | Unknown function | QFQ04729.1 | 0 | BLASTp |
| Replicative DNA helicase (E.C.3.6.4.12), Bacteriophage; Helicase-loader, Helicase, DNA replication; HET: ADP; 4.1A {Escherichia coli O111:NM}; Related PDB entries: 6BBM_W 6BBM_V 6BBM_X 6BBM_Y | Replication | QFQ04730.1 | 1.20 × 10−13 | HHpred |
| Hypothetical protein | Unknown function | QFQ04731.1 | 6.00 × 10−80 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04732.1 | 6.00 × 10−109 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04733.1 | 2.00 × 10−75 | BLASTp |
| DUF1064 domain-containing protein | Unknown function | QFQ04734.1 | 4.00 × 10−94 | BLASTp |
| Antitermination protein | Replication | QFQ04735.1 | 4.00 × 10−97 | BLASTp |
| Hypothetical protein | Replication | QFQ04736.1 | 2.00 × 10−32 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04737.1 | 6.00 × 10−164 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04738.1 | 1.00 × 10−60 | BLASTp |
| DNA mismatch endonuclease (E.C.3.1.-.-)/DNA complex; PROTEIN-DNA complex, mismatch, intercalation, zinc; 2.3A {Escherichia coli} SCOP: c.52.1.15 | DNA repair | QFQ04739.1 | 9.50 × 10−9 | HHpred |
| Hypothetical protein | Unknown function | QFQ04740.1 | 5.00 × 10−159 | BLASTp |
| DUF2280 domain-containing protein | Unknown function | QFQ04741.1 | 6.00 × 10−112 | BLASTp |
| Phage terminase large subunit | Replication | QFQ04742.1 | 0 | BLASTp |
| DUF1073 domain-containing protein | Unknown function | QFQ04743.1 | 0 | BLASTp |
| Phage head morphogenesis protein | Structural | QFQ04744.1 | 0 | BLASTp |
| Possible nuclease of RNase H fold, RuvC/YqgF family | Replication | QFQ04745.1 | 1.00 × 10−129 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04746.1 | 5.00 × 10−66 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04747.1 | 1.00 × 10−38 | BLASTp |
| Prohead core protein protease (E.C.3.4.99.-); protease pentamer, phage T4, prohead; 1.943A {Enterobacteria phage T4}; Related PDB entries: 5JBL_E 5JBL_B 5JBL_C 5JBL_A | Structural | QFQ04748.1 | 3.7 | HHpred |
| Capsid fibre protein; bacteriophage, phi29, prohead, VIRUS; HET: SO4; 1.8A {Bacillus phage phi29}; Related PDB entries: 6QYY_E 6QYY_A 6QYY_D 6QYY_C 6QYY_F | Structural | QFQ04749.1 | 0.0013 | HHpred |
| Capsid Stabilising Protein, Major Capsid; Major Capsid Protein, Capsid Stabilising; 3.6A {Pseudoalteromonas phage TW1}; Related PDB entries: 5WK1_A 5WK1_D 5WK1_C 5WK1_F 5WK1_E 5WK1_G | Structural | QFQ04750.1 | 1.30 × 10−33 | HHpred |
| Mu-like prophage FluMu protein gp35; structural genomics, Haemophilus influenzae, hypothetical; NMR {Haemophilus influenzae} SCOP: l.1.1.1, d.344.1.1, a.140.3.2 | Structural | QFQ04751.1 | 0.005 | HHpred |
| PORTAL PROTEIN, 15 PROTEIN, HEAD; VIRAL PROTEIN, VIRAL INFECTION, TAILED; 7.2A {BACILLUS PHAGE SPP1}; Related PDB entries: 5A20_D 5A20_C 5A21_D 2KBZ_A | Structural | QFQ04752.1 | 0.15 | HHpred |
| Hypothetical protein | Unknown function | QFQ04753.1 | 1.00 × 10−38 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04754.1 | 2.00 × 10−111 | BLASTp |
| Minor tail protein U; Mixed Alpha-Beta fold, VIRAL PROTEIN; HET: MSE, SO4; 2.7A {Enterobacteria phage lambda} SCOP: d.323.1.1; Related PDB entries: 3FZ2_L 3FZ2_K 3FZ2_F 3FZB_E 3FZB_B 3FZB_A 3FZB_I 3FZB_D 3FZB_J 3FZB_H 3FZB_C 3FZ2_I 3FZB_F 3FZ2_A 3FZB_G 3FZ2_D 3FZ2_J 3FZ2_H 3FZ2_E 3FZ2_B 3FZ2_G 1Z1Z_A | Structural | QFQ04755.1 | 3.70 × 10−17 | HHpred |
| Hypothetical protein | Unknown function | QFQ04756.1 | 0 | BLASTp |
| Tail assembly chaperone; Bacteriophage HK97, morphogenesis, tail assembly; HET: MSE; 2.3A {Enterobacteria phage HK97}; Related PDB entries: 2OB9_B | Structural | QFQ04757.1 | 6.5 | HHpred |
| Hypothetical protein | Unknown function | QFQ04758.1 | 2.00 × 10−48 | BLASTp |
| Zinc ribbon domain-containing protein | Unknown function | QFQ04759.1 | 2.00 × 10−117 | BLASTp |
| Phage tail protein | Structural | QFQ04760.1 | 0 | BLASTp |
| DUF2460 | Unknown function | QFQ04761.1 | 1.00 × 10−161 | BLASTp |
| DUF2163 domain-containing protein | Unknown function | QFQ04762.1 | 0 | BLASTp |
| SGNH/GDSL hydrolase family protein (sialate O-acetylesterase;cellulosome enzyme) | Lysis | QFQ04763.1 | 0 | BLASTp |
| Putative Exo-alpha-sialidase; Carbohydrate-Binding Module, Bacterial Pathogen, Sialic; HET: SIA; 2.2A {CLOSTRIDIUM PERFRINGENS}; Related PDB entries: 2V73_A | Lysis | QFQ04764.1 | 0.0032 | HHpred |
| C40 family peptidase | Lysis | QFQ04765.1 | 2.00 × 10−98 | BLASTp |
| Putative phage tail protein | Structural | QFQ04766.1 | 0 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04767.1 | 4.00 × 10−87 | BLASTp |
| Glycosyl hydrolase 108 | Lysis | QFQ04768.1 | 2.00 × 10−141 | BLASTp |
| Anaerobic dehydrogenase | Lysis | QFQ04769.1 | 7.00 × 10−73 | BLASTp |
| Hypothetical protein | Unknown function | QFQ04770.1 | 4.00 × 10−168 | BLASTp |
| Y-family DNA polymerase | Defence | QFQ04771.1 | 0 | BLASTp |
| Trans-lesion error-prone DNA polymerase V autoproteolytic subunit | Defence | QFQ04772.1 | 6.00 × 10−115 | BLASTp |
| Site-specific integrase | Lysogeny | QFQ04773.1 | 0 | BLASTp |
| Strain | Country | Date | Isolation Source | KL | OC | GI19606 | Additional Resistance Genes 2 | Tn6551 | Φ 3 | p1/p2 4 | GenBank no. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ATCC 19606 | US | <1948 | Urine | 3 | 1 | Y | − | + | + | +/+ | CP045110 |
| MSP4-16 | India | 2010 | Mangrove soil | 3 | 1 | Y | − | + | + | +/+ | AODW01 |
| ab736 | US | 2015 | Blood | 3 | 1 | Y | − | + | + | −/+ | CP015121 |
| NBRC 110495 | Japan | 2008 | Human abscess | 3 | 1 | − | aadB5 | − | − | −/+ 6 | BBOR01 |
| NBRC 110494 | Japan | 2008 | Burned skin | 3 | 1 | − | aadB5 | − | − | −/+ 6 | BBTE01 |
| NBRC 110493 | Japan | 2008 | Burned skin | 3 | 1 | − | aadB5 | − | − | −/+ 6 | BBOQ01 |
| GTC 03329 | Japan | 2008 | Human abscess | 3 | 1 | − | aadB5 | − | − | −/+ 6 | BBNJ01 |
| GTC 03325 | Japan | 2008 | Burned skin | 3 | 1 | − | aadB5 | − | − | −/+ 6 | BBSP01 |
| GTC 03324 | Japan | 2008 | Burned skin | 3 | 1 | − | aadB5 | − | − | −/+ 6 | BBNH01 |
| AB_095 | Pakistan | 2016 | ICU washroom sink | 3 | 1 | − | tet(B), sul1, sul2, blaGES-11,dfrA7, aacA4, oxa23, aphA6 | − | − | −/− | RHZR01 |
| AB_165 | Pakistan | 2016 | Alcohol foam dispenser in ICU | 3 | 1 | − | tet(B), sul1, sul2, blaGES-11,dfrA7, aacA4, oxa23, aphA6 | − | − | −/− | RHZA01 |
| 4300STDY7045730 | Thailand | 2016 | na 7 | 57 | 1 | − | tet39 | − | − | −/− | UFJF01 |
| WE2714 | China | na | na | 32 | 6 | − | − | − | − | −/− | QKVH01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamidian, M.; Blasco, L.; Tillman, L.N.; To, J.; Tomas, M.; Myers, G.S.A. Analysis of Complete Genome Sequence of Acinetobacter baumannii Strain ATCC 19606 Reveals Novel Mobile Genetic Elements and Novel Prophage. Microorganisms 2020, 8, 1851. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121851
Hamidian M, Blasco L, Tillman LN, To J, Tomas M, Myers GSA. Analysis of Complete Genome Sequence of Acinetobacter baumannii Strain ATCC 19606 Reveals Novel Mobile Genetic Elements and Novel Prophage. Microorganisms. 2020; 8(12):1851. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121851
Chicago/Turabian StyleHamidian, Mohammad, Lucia Blasco, Lauren N. Tillman, Joyce To, María Tomas, and Garry S. A. Myers. 2020. "Analysis of Complete Genome Sequence of Acinetobacter baumannii Strain ATCC 19606 Reveals Novel Mobile Genetic Elements and Novel Prophage" Microorganisms 8, no. 12: 1851. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121851