Tn6603, a Carrier of Tn5053 Family Transposons, Occurs in the Chromosome and in a Genomic Island of Pseudomonas aeruginosa Clinical Strains




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Growth Media
2.2. Transposons and Plasmid Constructions
2.3. Conjugation, Transposition Assays, and Analysis of Transconjugants
2.4. DNA Techniques, Nextgen Sequencing, and Analysis
2.5. Nucleotide Sequence Accession Numbers
3. Results
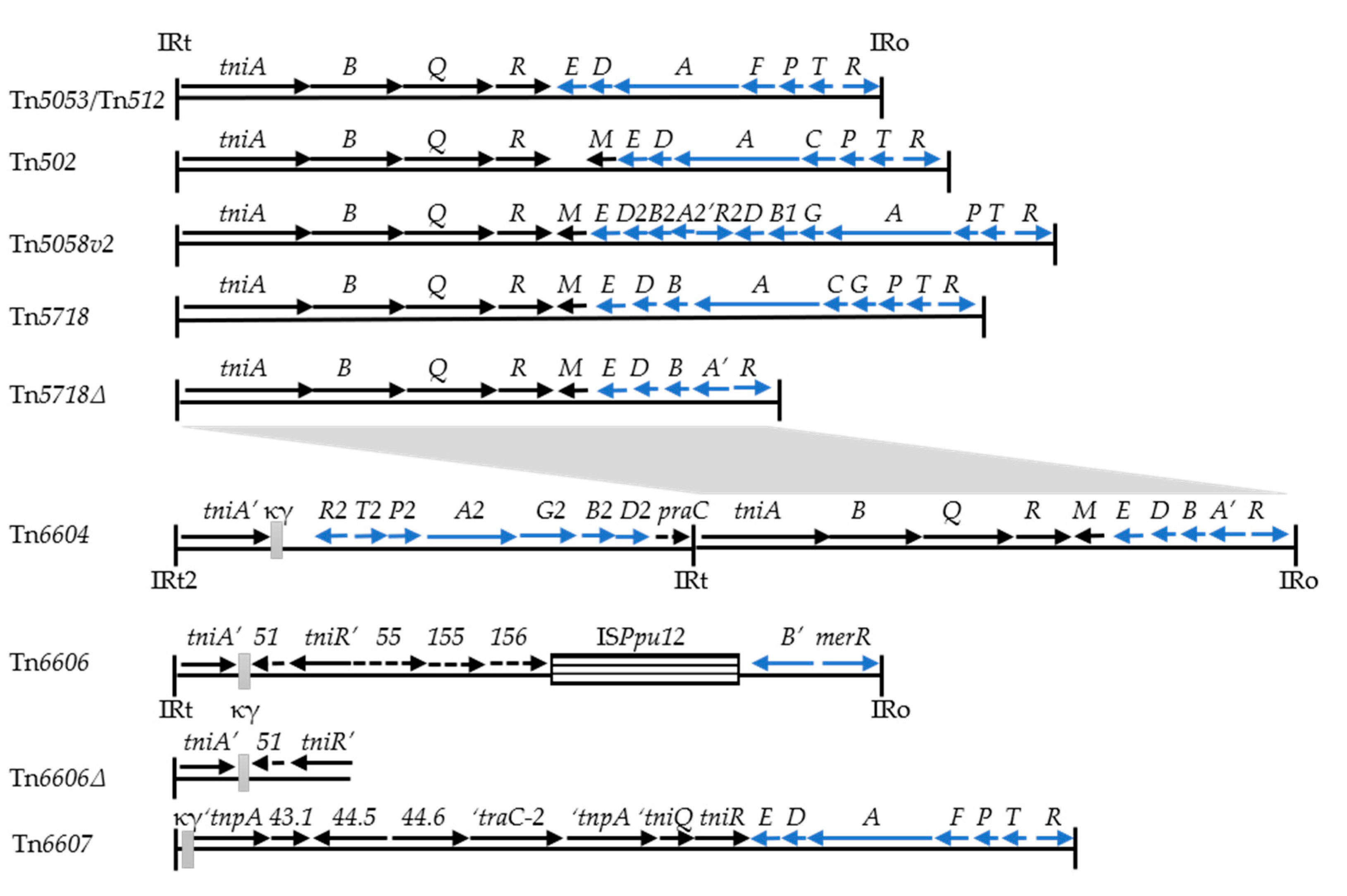
3.1. Isolation of Res Site Hunter Transposons, Including the New Element Tn6604, from Clinical Strains of P. aeruginosa
3.2. Isolation of Res Site Hunter Transposons Nested with a Carrier Transposon
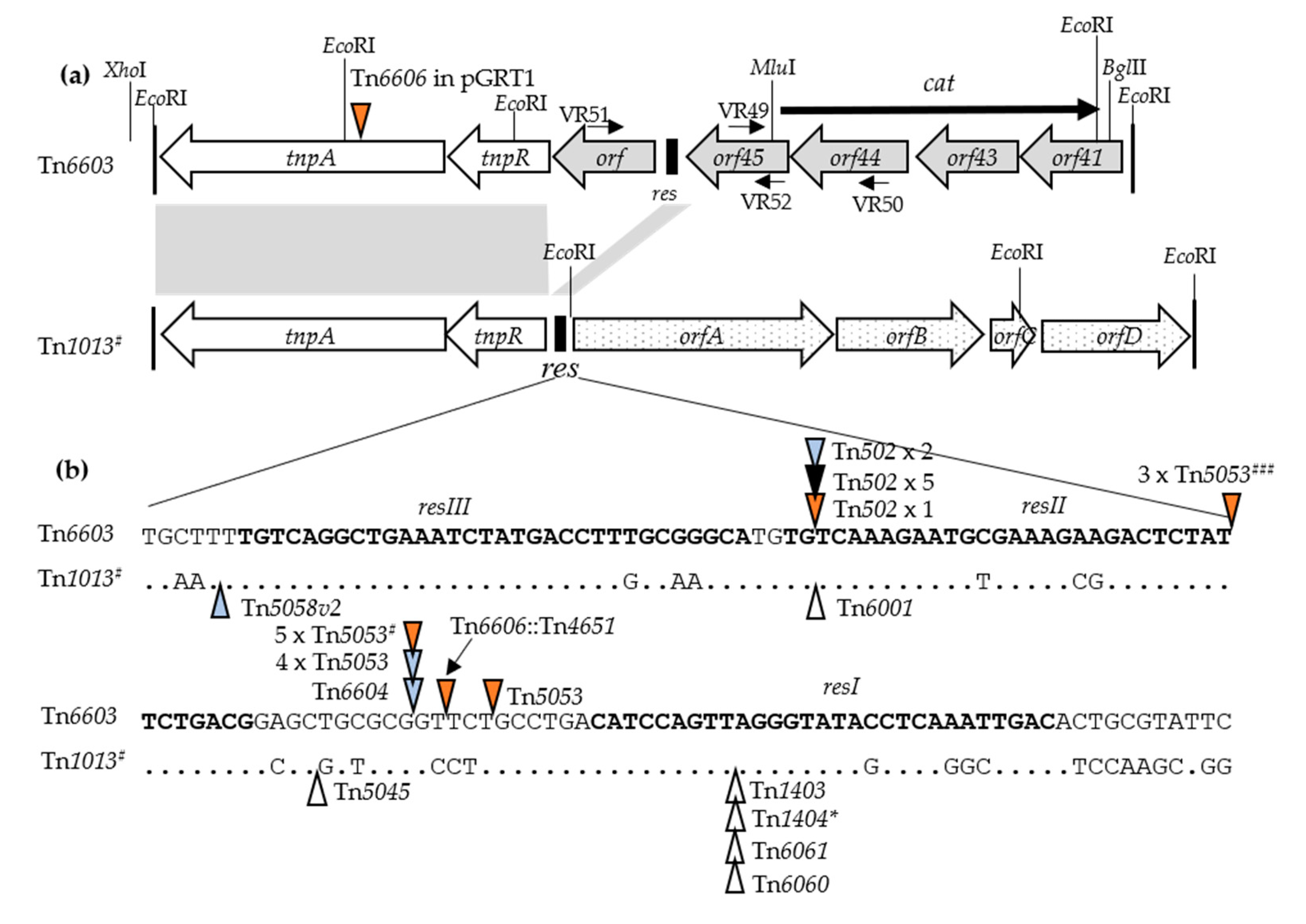
3.3. Isolation and Characterization of the Carrier Transposon Tn6603
3.4. Distribution of Tn6603 and Tn6603-Nested Transposons and Their Association with Genomic Island PAGI-5v
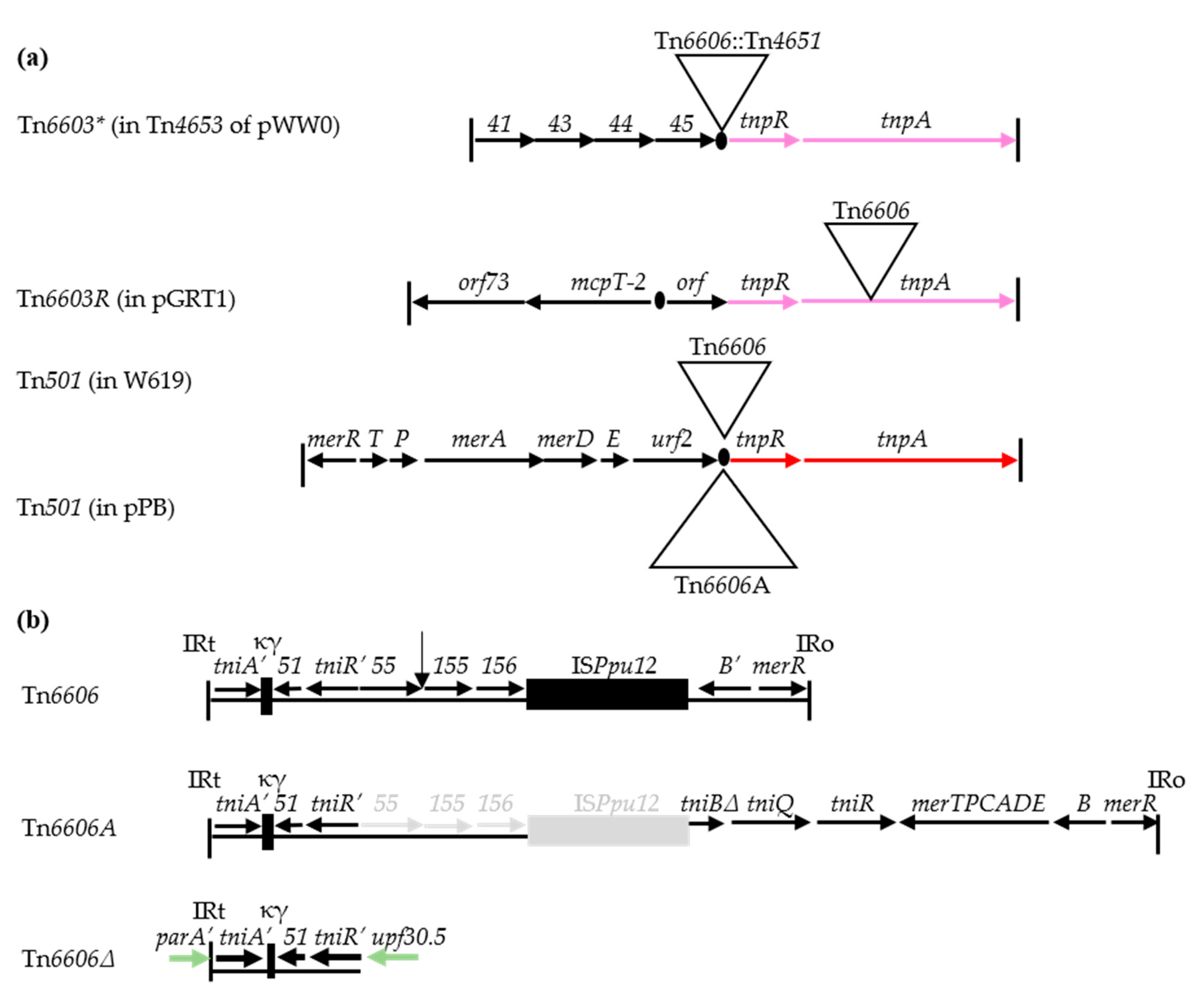
3.5. Detection of Tn6606 and Related Elements: Defective Transposons of the Tn5053 Family
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic resistance in Pseudomonas aeruginosa–Mechanisms, epidemiology and evolution. Drug Resist. Update 2019, 44, 100640. [Google Scholar] [CrossRef] [PubMed]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, V.L.; Ozer, E.A.; Hauser, A.R. The accessory genome of Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 2010, 74, 621–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinsted, J.; De La Cruz, F.; Schmitt, R. The Tn21 subgroup of bacterial transposable elements. Plasmid 1990, 24, 163–189. [Google Scholar] [CrossRef]
- Liebert, C.A.; Hall, R.M.; Summers, A.O. Transposon Tn21, flagship of the floating genome. Microbiol. Mol. Biol. Rev. 1999, 63, 507–522. [Google Scholar] [CrossRef] [Green Version]
- Battle, S.E.; Meyer, F.; Rello, J.; Kung, V.L.; Hauser, A.R. Hybrid pathogenicity island PAGI-5 contributes to the highly virulent phenotype of a Pseudomonas aeruginosa isolate in mammals. J. Bacteriol. 2008, 190, 7130–7140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.S.; Yoon, E.-J.; Lee, H.; Jeong, S.H.; Lee, K.-W. Clonal dissemination of Pseudomonas aeruginosa ST235 carrying blaIMP-6 and emergence of blaGES-24 and blaIMP-10 on novel genomic islands PAGI-15 and -16 in Korea. Antimicrob. Agents Chemother. 2016, 60. [Google Scholar] [CrossRef] [Green Version]
- Kawalek, A.; Kotecka, K.; Modrzejewska, M.; Gawor, J.; Jagura-Burdzy, G.; Bartosik, A.A. Genome sequence of Pseudomonas aeruginosa PAO1161, a PAO1 derivative with the ICEPae1161 integrative and conjugative element. BMC Genom. 2020, 21, 14. [Google Scholar] [CrossRef]
- Jacoby, G.A. Resistance plasmids of Pseudomonas. In The Biology of Pseudomonas; Elsevier BV: Amsterdam, The Netherlands, 1986; pp. 265–293. [Google Scholar]
- Boronin, A.M. Diversity of Pseudomonas plasmids: To what extent? FEMS Microbiol. Lett. 1992, 100, 461–467. [Google Scholar] [CrossRef]
- Schlüter, A.; Szczepanowski, R.; Pühler, A.; Top, E.M. Genomics of IncP-1 antibiotic resistance plasmids isolated from wastewater treatment plants provides evidence for a widely accessible drug resistance gene pool. FEMS Microbiol. Rev. 2007, 31, 449–477. [Google Scholar] [CrossRef]
- Cazares, A.; Moore, M.P.; Hall, J.P.J.; Wright, L.L.; Grimes, M.; Emond-Rhéault, J.-G.; Pongchaikul, P.; Santanirand, P.; Lévesque, R.; Fothergill, J.L.; et al. A megaplasmid family driving dissemination of multidrug resistance in Pseudomonas. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, S.R.; Recchia, G.D.; Stokes, H.W.; Hall, R.M.; Gong, Y.-F.; Robinson, B.S.; Rose, R.E.; Deminie, C.; Spicer, T.P.; Stock, D.; et al. Family of class 1 integrons Related to In4 from Tn1696. Antimicrob. Agents Chemother. 2001, 45, 2319–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrovski, S.; Stanisich, V.A. Embedded elements in the IncPβ plasmids R772 and R906 can be mobilized and can serve as a source of diverse and novel elements. Microbiology 2011, 157, 1714–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues, S.; Da Silva, G.J.; Nielsen, K.M. Integrons. Mob. Genet. Elem. 2012, 2, 211–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.M. Integrons and gene cassettes: Hotspots of diversity in bacterial genomes. Ann. N. Y. Acad. Sci. 2012, 1267, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, 31. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R.; Tsafnat, G.; Coiera, E.; Iredell, J.R. Gene cassettes and cassette arrays in mobile resistance integrons. FEMS Microbiol. Rev. 2009, 33, 757–784. [Google Scholar] [CrossRef] [Green Version]
- Minakhina, S.; Kholodii, G.; Mindlin, S.; Yurieva, O.; Nikiforov, V. Tn5053 family transposons are res site hunters sensing plasmidal res sites occupied by cognate resolvases. Mol. Microbiol. 1999, 33, 1059–1068. [Google Scholar] [CrossRef]
- Rådström, P.; Sköld, O.; Swedberg, G.; Flensburg, J.; Roy, P.H.; Sundström, L. Transposon Tn5090 of plasmid R751, which carries an integron, is related to Tn7, Mu, and the retroelements. J. Bacteriol. 1994, 176, 3257–3268. [Google Scholar] [CrossRef] [Green Version]
- Kholodii, G.; Mindlin, S.Z.; Bass, I.; Yurieva, O.; Minakhina, S.; Nikiforov, V. Four genes, two ends, and a res region are involved in transposition of Tn5053: A paradigm for a novel family of transposons carrying either a mer operon or an integron. Mol. Microbiol. 1995, 17, 1189–1200. [Google Scholar] [CrossRef]
- Petrovski, S.; Blackmore, D.W.; Jackson, K.L.; Stanisich, V.A. Mercury(II)-resistance transposons Tn502 and Tn512, from Pseudomonas clinical strains, are structurally different members of the Tn5053 family. Plasmid 2011, 65, 58–64. [Google Scholar] [CrossRef]
- Mindlin, S.; Kholodii, G.; Gorlenko, Z.; Minakhina, S.; Minakhin, L.; Kalyaeva, E.; Kopteva, A.; Petrova, M.; Yurieva, O.; Nikiforov, V. Mercury resistance transposons of Gram-negative environmental bacteria and their classification. Res. Microbiol. 2001, 152, 811–822. [Google Scholar] [CrossRef]
- Haines, A.S.; Akhtar, P.; Stephens, E.R.; Jones, K.; Thomas, C.M.; Perkins, C.D.; Williams, J.R.; Day, M.J.; Fry, J.C. Plasmids from freshwater environments capable of IncQ retrotransfer are diverse and include pQKH54, a new IncP-1 subgroup archetype. Microbiology 2006, 152, 2689–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurieva, O.; Kholodii, G.; Minakhin, L.; Gorlenko, Z.; Kalyaeva, E.; Mindlin, S.; Nikiforov, V. Intercontinental spread of promiscuous mercury-resistance transposons in environmental bacteria. Mol. Microbiol. 1997, 24, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Mindlin, S.; Minakhin, L.; Petrova, M.; Kholodii, G.; Minakhina, S.; Gorlenko, Z.; Nikiforov, V. Present-day mercury resistance transposons are common in bacteria preserved in permafrost grounds since the Upper Pleistocene. Res. Microbiol. 2005, 156, 994–1004. [Google Scholar] [CrossRef]
- Mindlin, S.; Petrova, M. Mercury resistance transposons. In Bacterial Integrative Mobile Genetic Elements; Roberts, A.P., Mullany, P., Eds.; Landes Biosciences: Austin, TX, USA, 2013; pp. 33–52. [Google Scholar]
- Kos, V.N.; Déraspe, M.; McLaughlin, R.E.; Whiteaker, J.D.; Roy, P.H.; Alm, R.A.; Corbeil, J.; Gardner, H. The resistome of Pseudomonas aeruginosa in relationship to phenotypic susceptibility. Antimicrob. Agents Chemother. 2015, 59, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Brown, H.J.; Stokes, H.W.; Hall, R.M. The integrons In0, In2, and In5 are defective transposon derivatives. J. Bacteriol. 1996, 178, 4429–4437. [Google Scholar] [CrossRef] [Green Version]
- Gillings, M.R. Integrons: Past, present, and future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [Green Version]
- Ghaly, T.M.; Chow, L.; Asher, A.J.; Waldron, L.S.; Gillings, M.R. Evolution of class 1 integrons: Mobilization and dispersal via food-borne bacteria. PLoS ONE 2017, 12, e0179169. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Król, J.E.; Suzuki, H.; Foster, B.; Van Houdt, R.; Brown, C.J.; Mergeay, M.; Top, E.M. Plasmids captured in C. metallidurans CH34: Defining the PromA family of broad-host-range plasmids. Antonie Van Leeuwenhoek 2009, 96, 193–204. [Google Scholar] [CrossRef]
- Zhang, M.; Brons, J.K.; van Elsas, J.D. The complete sequences and ecological roles of two IncP-1β plasmids, pHB44 and pBS64, isolated from the mycosphere of Laccaria proxima. Front. Microbiol. 2016, 7, 909. [Google Scholar] [CrossRef]
- Mindlin, S.Z.; Minakhina, S.V.; Kholodiĭ, G.; Kopteva, A.V.; Nikiforov, V.G. Incorporation of Tn5053 and Tn402 into various plasmids. Genetika 1996, 32, 1426–1430. [Google Scholar]
- Kamali-Moghaddam, M.; Sundstrom, L. Transposon targeting determined by resolvase. FEMS Microbiol. Lett. 2000, 186, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Kamali-Moghaddam, M. Arrayed transposase-binding sequences on the ends of transposon Tn5090/Tn402. Nucl. Acids Res. 2001, 29, 1005–1011. [Google Scholar] [CrossRef] [Green Version]
- Petrovski, S.; Stanisich, V.A. Tn502 and Tn512 Are res site hunters that provide evidence of resolvase-independent transposition to random sites. J. Bacteriol. 2010, 192, 1865–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, P.R.; Scott, M.; Worden, P.; Huntington, P.; Hudson, B.; Karagiannis, T.; Charles, I.G.; Djordjevic, S.P. Genomic islands 1 and 2 play key roles in the evolution of extensively drug-resistant ST235 isolates of Pseudomonas aeruginosa. Open Biol. 2016, 6. [Google Scholar] [CrossRef]
- Sinclair, M.; Holloway, B.W. A chromosomally located transposon in Pseudomonas aeruginosa. J. Bacteriol. 1982, 151, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R.; Brown, H.J.; Hall, R.M. Characterization and movement of the class 1 integron known as Tn2521 and Tn1405. Antimicrob. Agents Chemother. 2002, 46, 1288–1294. [Google Scholar] [CrossRef] [Green Version]
- Petrova, M.; Gorlenko, Z.; Mindlin, S.Z. Tn5045, a novel integron-containing antibiotic and chromate resistance transposon isolated from a permafrost bacterium. Res. Microbiol. 2011, 162, 337–345. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Palombo, E.A.; Yusoff, K.; Stanisich, V.A.; Krishnapillai, V.; Willetts, N. Cloning and genetic analysis of tra cistrons of the Tra 2/Tra 3 region of plasmid RP1. Plasmid 1989, 22, 59–69. [Google Scholar] [CrossRef]
- Stanisich, V.A.; Bennett, P.M.; Oritz, J.M. A molecular analysis of transductional marker rescue involving P-group plasmids in Pseudomonas aeruginosa. Mol. Gen. Genet. MGG 1976, 143, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Lyras, D.; Palombo, E.A.; Stanisich, V.A. Characterization of a Tra 2 function of RP1 that affects growth of Pseudomonas aeruginosa PAO and surface exclusion in Escherichia coli K12. Plasmid 1992, 27, 105–118. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, P.M.; Grinsted, J.; Richmond, M.H. Transposition of TnA does not generate deletions. Mol. Genet. Genom. 1977, 154, 205–211. [Google Scholar] [CrossRef]
- Wallace, L.J.; Ward, J.M.; Richmond, M.H. The tnpR gene product of TnA is required for transposition immunity. Mol. Genet. Genom. 1981, 184, 87–91. [Google Scholar] [CrossRef]
- Rose, R.E. The nucleotide sequence of pACYC184. Nucl. Acids Res. 1988, 16, 355. [Google Scholar] [CrossRef]
- Bolivar, F. Construction and characterization of new cloning vehicles III. Derivatives of plasmid pBR322 carrying unique Eco RI sites for selection of Eco RI generated recombinant DNA molecules. Gene 1978, 4, 121–136. [Google Scholar] [CrossRef]
- Yusoff, K.; Stanisich, V.A. Location of a function on RP1 that fertility inhibits IncW plasmids. Plasmid 1984, 11, 178–181. [Google Scholar] [CrossRef]
- Stanisich, V.A.; Arwas, R.; Bennett, P.M.; De La Cruz, F. Characterization of Pseudomonas mercury-resistance transposon Tn502, which has a preferred insertion site in RP1. Microbiology 1989, 135, 2909–2915. [Google Scholar] [CrossRef] [Green Version]
- Haines, A.S.; Jones, K.; Batt, S.M.; Kosheleva, I.A.; Thomas, C.M. Sequence of plasmid pBS228 and reconstruction of the IncP-1α phylogeny. Plasmid 2007, 58, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Kholodii, G.; Gorlenko, Z.; Mindlin, S.; Hobman, J.; Nikiforov, V. Tn5041-like transposons: Molecular diversity, evolutionary relationships and distribution of distinct variants in environmental bacteria. Microbiology 2002, 148, 3569–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szuplewska, M.; Ludwiczak, M.; Lyzwa, K.; Czarnecki, J.; Bartosik, D. Mobility and generation of mosaic non-autonomous transposons by Tn3-derived Inverted-Repeat Miniature Elements (TIMEs). PLoS ONE 2014, 9, e105010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneiker, S.; Keller, M.; Dröge, M.; Lanka, E.; Pühler, A.; Selbitschka, W. The genetic organization and evolution of the broad host range mercury resistance plasmid pSB102 isolated from a microbial population residing in the rhizosphere of alfalfa. Nucl. Acids Res. 2001, 29, 5169–5181. [Google Scholar] [CrossRef]
- Partridge, S.R. Analysis of antibiotic resistance regions in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 820–855. [Google Scholar] [CrossRef] [Green Version]
- Stokes, H.W.; Elbourne, L.D.H.; Hall, R.M. Tn1403, a multiple-antibiotic resistance transposon made up of three distinct transposons. Antimicrob. Agents Chemother. 2007, 51, 1827–1829. [Google Scholar] [CrossRef] [Green Version]
- Tseng, S.-P.; Tsai, J.-C.; Teng, L.-J.; Hsueh, P.-R. Dissemination of transposon Tn6001 in carbapenem-non-susceptible and extensively drug-resistant Pseudomonas aeruginosa in Taiwan. J. Antimicrob. Chemother. 2009, 64, 1170–1174. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, P.R.; Merlino, J.; Labbate, M.; Cheong, E.Y.-L.; Gottlieb, T.; Stokes, H.W. Tn6060, a transposon from a genomic island in a Pseudomonas aeruginosa clinical isolate that includes two class 1 integrons. Antimicrob. Agents Chemother. 2009, 53, 5294–5296. [Google Scholar] [CrossRef] [Green Version]
- Coyne, S.; Courvalin, P.; Galimand, M. Acquisition of multidrug resistance transposon Tn6061 and IS6100-mediated large chromosomal inversions in Pseudomonas aeruginosa clinical isolates. Microbiology 2010, 156, 1448–1458. [Google Scholar] [CrossRef] [Green Version]
- Greated, A.; Lambertsen, L.; Williams, P.A.; Thomas, C.M. Complete sequence of the IncP-9 TOL plasmid pWW0 from Pseudomonas putida. Environ. Microbiol. 2002, 4, 856–871. [Google Scholar] [CrossRef]
- Botelho, J.; Roberts, A.P.; León-Sampedro, R.; Grosso, F.; Peixe, L. Carbapenemases on the move: It’s good to be on ICEs. Mob. DNA 2018, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M.; Farinha, M.A.; Jansons, I. Nucleotide sequence of the Pseudomonas aeruginosa insertion Sequence IS222: Another Member of the IS3 Family. Plasmid 1994, 31, 222–228. [Google Scholar] [CrossRef]
- Kiewitz, C.; Larbig, K.; Klockgether, J.; Weinel, C.; Tümmler, B. Monitoring genome evolution ex vivo: Reversible chromosomal integration of a 106 kb plasmid at two tRNA(Lys) gene loci in sequential Pseudomonas aeruginosa airway isolates. Microbiology 2000, 146, 2365–2373. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Gurkar, A.U.; Lory, S. Interstrain transfer of the large pathogenicity island (PAPI-1) of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2006, 103, 19830–19835. [Google Scholar] [CrossRef] [Green Version]
- Botelho, J.; Mourão, J.; Roberts, A.P.; Peixe, L. Comprehensive genome data analysis establishes a triple whammy of carbapenemases, ICEs and multiple clinically relevant bacteria. Microb. Genom. 2020, 6. [Google Scholar] [CrossRef]
- Guglielmini, J.; Quintais, L.; Garcillán-Barcia, M.P.; de la Cruz, F.; Rocha, E.P. The repertoire of ICE in prokaryotes underscores the unity, diversity, and ubiquity of conjugation. PLoS Genet. 2011, 7, e1002222. [Google Scholar] [CrossRef]
- Carter, M.Q.; Chen, J.; Lory, S. The Pseudomonas aeruginosa pathogenicity island PAPI-1 is transferred via a novel type IV pilus. J. Bacteriol. 2010, 192, 3249–3258. [Google Scholar] [CrossRef] [Green Version]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guédon, G. Conjugative and mobilizable genomic islands in bacteria: Evolution and diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [Google Scholar] [CrossRef] [Green Version]
- Toleman, M.A.; Walsh, T.R. Combinatorial events of insertion sequences and ICE in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 912–935. [Google Scholar] [CrossRef] [Green Version]
- Molina, L.; Duque, E.; Gómez, M.J.; Krell, T.; Lacal, J.; García-Puente, A.; García, V.; Matilla, M.A.; Ramos, J.L.; Segura, A. The pGRT1 plasmid of Pseudomonas putida DOT-T1E encodes functions relevant for survival under harsh conditions in the environment. Environ. Microbiol. 2011, 13, 2315–2327. [Google Scholar] [CrossRef]
- Nakazawa, T. Travels of a Pseudomonas, from Japan around the world. Environ. Microbiol. 2002, 4, 782–786. [Google Scholar] [CrossRef]
- Wu, X.; Monchy, S.; Taghavi, S.; Zhu, W.; Ramos, J.; van der Lelie, D. Comparative genomics and functional analysis of niche-specific adaptation in Pseudomonas putida. FEMS Microbiol. Rev. 2011, 35, 299–323. [Google Scholar] [CrossRef] [Green Version]
- Baggi, G.; Barbieri, P.; Galli, E.; Tollari, S. Isolation of a Pseudomonas stutzeri strain that degrades o-xylene. Appl. Environ. Microbiol. 1987, 53, 2129–2132. [Google Scholar] [CrossRef] [Green Version]
- Coetzee, J.N. Mobilization of the Proteus mirabilis Chromosome by R plasmid R772. J. Gen. Microbiol. 1978, 108, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.A.; Jones, R.M.; Shaw, L.E. A Third transposable element, ISPpu12, from the toluene-xylene catabolic plasmid pWW0 of Pseudomonas putida mt-2. J. Bacteriol. 2002, 184, 6572–6580. [Google Scholar] [CrossRef] [Green Version]
- Kamachi, K.; Sota, M.; Tamai, Y.; Nagata, N.; Konda, T.; Inoue, T.; Top, E.M.; Arakawa, Y. Plasmid pBP136 from Bordetella pertussis represents an ancestral form of IncP-1β plasmids without accessory mobile elements. Microbiology 2006, 152, 3477–3484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smalla, K.; Haines, A.S.; Jones, K.; Krögerrecklenfort, E.; Heuer, H.; Schloter, M.; Thomas, C.M. Increased Abundance of IncP-1β plasmids and mercury resistance genes in mercury-polluted river sediments: First discovery of IncP-1β plasmids with a complex mer transposon as the sole accessory element. Appl. Environ. Microbiol. 2006, 72, 7253–7259. [Google Scholar] [CrossRef] [Green Version]
- Dealtry, S.; Ding, G.-C.; Weichelt, V.; Dunon, V.; Schlüter, A.; Martini, M.C.; Del Papa, M.F.; Lagares, A.; Amos, G.C.A.; Wellington, E.M.H.; et al. Cultivation-independent screening revealed hot spots of IncP-1, IncP-7 and IncP-9 plasmid occurrence in different environmental habitats. PLoS ONE 2014, 9, e89922. [Google Scholar] [CrossRef]
- Klockgether, J.; Cramer, N.; Wiehlmann, L.; Davenport, C.F.; Tümmler, B. Pseudomonas aeruginosa genomic structure and diversity. Front. Microbiol. 2011, 2, 150. [Google Scholar] [CrossRef] [Green Version]
- Sevastsyanovich, Y.R.; Krasowiak, R.; Bingle, L.E.H.; Haines, A.S.; Sokolov, S.; Kosheleva, I.A.; Leuchuk, A.A.; Titok, M.; Smalla, K.; Thomas, C.M. Diversity of IncP-9 plasmids of Pseudomonas. Microbiology 2008, 154, 2929–2941. [Google Scholar] [CrossRef] [Green Version]
- Benson, S.; Shapiro, J. TOL is a broad-host-range plasmid. J. Bacteriol. 1978, 135, 278–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klockgether, J.; Würdemann, D.; Reva, O.; Wiehlmann, L.; Tümmler, B. Diversity of the abundant pKLC102/PAGI-2 family of genomic islands in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 2443–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, P.R.; Scott, M.J.; Djordjevic, S.P. Genomic islands 1 and 2 carry multiple antibiotic resistance genes in Pseudomonas aeruginosa ST235, ST253, ST111 and ST175 and are globally dispersed. J. Antimicrob. Chemother. 2016, 72, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Klockgether, J.; Reva, O.; Larbig, K.; Tümmler, B. Sequence analysis of the mobile genome island pKLC102 of Pseudomonas aeruginosa C. J. Bacteriol. 2004, 186, 518–534. [Google Scholar] [CrossRef] [Green Version]
- Szczepanowski, R.; Eikmeyer, F.; Harfmann, J.; Blom, J.; Rogers, L.M.; Top, E.M.; Schlüter, A. Sequencing and comparative analysis of IncP-1α antibiotic resistance plasmids reveal a highly conserved backbone and differences within accessory regions. J. Biotechnol. 2011, 155, 95–103. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid a,b | Relevant Features or Derivations | Reference/Source |
|---|---|---|
| Conjugative Plasmids | ||
| pUB307 | Aps deletant of RP1; has parA-resIncP region; Tcr Kmr | [48] |
| pUB1601 | PstI-ligation derivative of RP1; lacks parA-resIncP region; Tcr Kmr | [49] |
| pSP116 c | pUB1601tet::Tn6603c; Kmr Cmr | This work |
| pSP144 | pUB1601tet::Ω1Tn502; Kmr Hgr | [37] |
| pSP145 | pUB1601tet::Tn501; Kmr Hgr | This work |
| Nonconjugative Plasmids | ||
| pBluescript II SK+ (pSK) | E. coli vector; Apr | Stratagene |
| pACYC184 | E. coli vector; Cmr Tcr | [50] |
| pBR322 | E. coli vector; Apr Tcr | [51] |
| pVS76 (pBR322-resIncP) | pBR322 with bla disrupted by a clone of the RP1 parA-resIncP region; Tcr | [52] |
| pVS982 | pBR322::Tn501; Apr Tcr Hgr | [53] |
| pVS983 | pBR322tet::Tn502; Apr Hgr | [53] |
| pSP111 d (pSK-Tn6603) | pSK with a clone of Tn6603 from P. aeruginosa PA65; Apr | This work |
| pSP112 e (pSK-Tn6603c) | pSP111 with a clone of the cat gene from pACYC184; Apr Cmr | This work |
| pSP114 f (pBR322-res6603) | pVS76 with parA-resIncP sequences removed and replaced with tnpA-orf-res region of Tn6603; Tcr | This work |
| P. aeruginosa Strain | Year Isolated a | Transfer Frequency to pUB307; Tn Detected f,g | Transfer Frequency to pUB1601; Tn Detected f,h,i | Location of Nested Tn (or Tn) in Clinical Strain j |
|---|---|---|---|---|
| RH19 | 1992 b | 6.2 × 10−3; Tn502 | 1.6 × 10−6; Tn6603::Tn502 | Ω1 |
| AW32 | 1991 c | 7.2 × 10−3; Tn502 | 3.2 × 10−8; Tn6603::Tn502 | orfX |
| AW57 | 1991 c | 1.1 × 10−3; Tn502 | 1.2 × 10−8; Tn502 | Ω2 |
| PA9 | 2015 d | 2.8 × 10−2; Tn502 | 2.0 × 10−8; Tn502 | Ω3 |
| AW54b | 1991 c | 2.5 × 10−4; Tn512 | <1.0 × 10−8; ND | Ω4 |
| AW60 | 1991 c | 1.6 × 10−4; Tn512 | <1.0 × 10−8; ND | Ω4 |
| PA8 | 2015 d | 1.6 × 10−4; Tn5053 | 1.0 × 10−5; Tn6603::Tn5053 | orfX |
| PS1 | 1988 e | 0.5 × 10−4; Tn5053 | 7.0 × 10−6; Tn6603::Tn5053 | orfX |
| AB1 | 1992 c | 1.2 × 10−4; Tn5053 | 3.4 × 10−6; Tn6603::Tn5053 | orfX |
| AW1 | 1991 c | 0.6 × 10−4; Tn5053 | 1.0 × 10−5; Tn6603::Tn5053 | orfX |
| PA118 | 2015 d | 1.2 × 10−2; Tn5058v2 | 1.2 × 10−6; Tn1013#::Tn5058v2 | Ω5 |
| PA41 | 2015 d | 2.9 × 10−5; Tn6604 | 3.0 × 10−7; Tn6603::Tn6604 | orfX |
| Bacterium | Source (Decade) | Transposon and Carrier | Ref |
|---|---|---|---|
| Pseudomonas putida mt-2 | Field soil/Japan (1960) | Tn6606::Tn4651 in pWW0 (IncP-9) | [74] |
| P. putida DOT-T1E | Wastewater/Spain (1990) | Tn6606 in pGRT1 a | [73] |
| P. putida W619 | Poplar root/stem/Belgium (2000) b | Tn6606 in chromosome | [75] |
| Pseudomonas stutzeri OX | Wastewater/Italy (1980) | Tn6606A in pPB a | [76] |
| Proteus mirabilis | Clinical isolate/USA (1970) | Tn6606Δ in R772 (IncP-1β) | [77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajabal, V.; Stanisich, V.A.; Petrovski, S. Tn6603, a Carrier of Tn5053 Family Transposons, Occurs in the Chromosome and in a Genomic Island of Pseudomonas aeruginosa Clinical Strains. Microorganisms 2020, 8, 1997. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121997
Rajabal V, Stanisich VA, Petrovski S. Tn6603, a Carrier of Tn5053 Family Transposons, Occurs in the Chromosome and in a Genomic Island of Pseudomonas aeruginosa Clinical Strains. Microorganisms. 2020; 8(12):1997. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121997
Chicago/Turabian StyleRajabal, Vaheesan, Vilma A. Stanisich, and Steve Petrovski. 2020. "Tn6603, a Carrier of Tn5053 Family Transposons, Occurs in the Chromosome and in a Genomic Island of Pseudomonas aeruginosa Clinical Strains" Microorganisms 8, no. 12: 1997. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121997