Exploring the Ambiguous Status of Coagulase-Negative Staphylococci in the Biosafety of Fermented Meats: The Case of Antibacterial Activity Versus Biogenic Amine Formation

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Staphylococcal Strains
2.2. Screening for Antibacterial Activity
2.3. Screening for Anticlostridial Activity
2.4. Assessment of Temperature and Proteinase K on the Stability of Antibacterial Activity in Broth from Staphylococcus sciuri IMDO-S72 and its Spectrum of Activity
2.5. Analysis of Biogenic Amines
2.5.1. Growth Conditions and Sampling
2.5.2. Determination of Biogenic Amines by Ultra-High-Performance Liquid Chromatography Coupled to Tandem Mass Spectrometry (UPLC-MS/MS)
2.6. Graphical Representation
3. Results
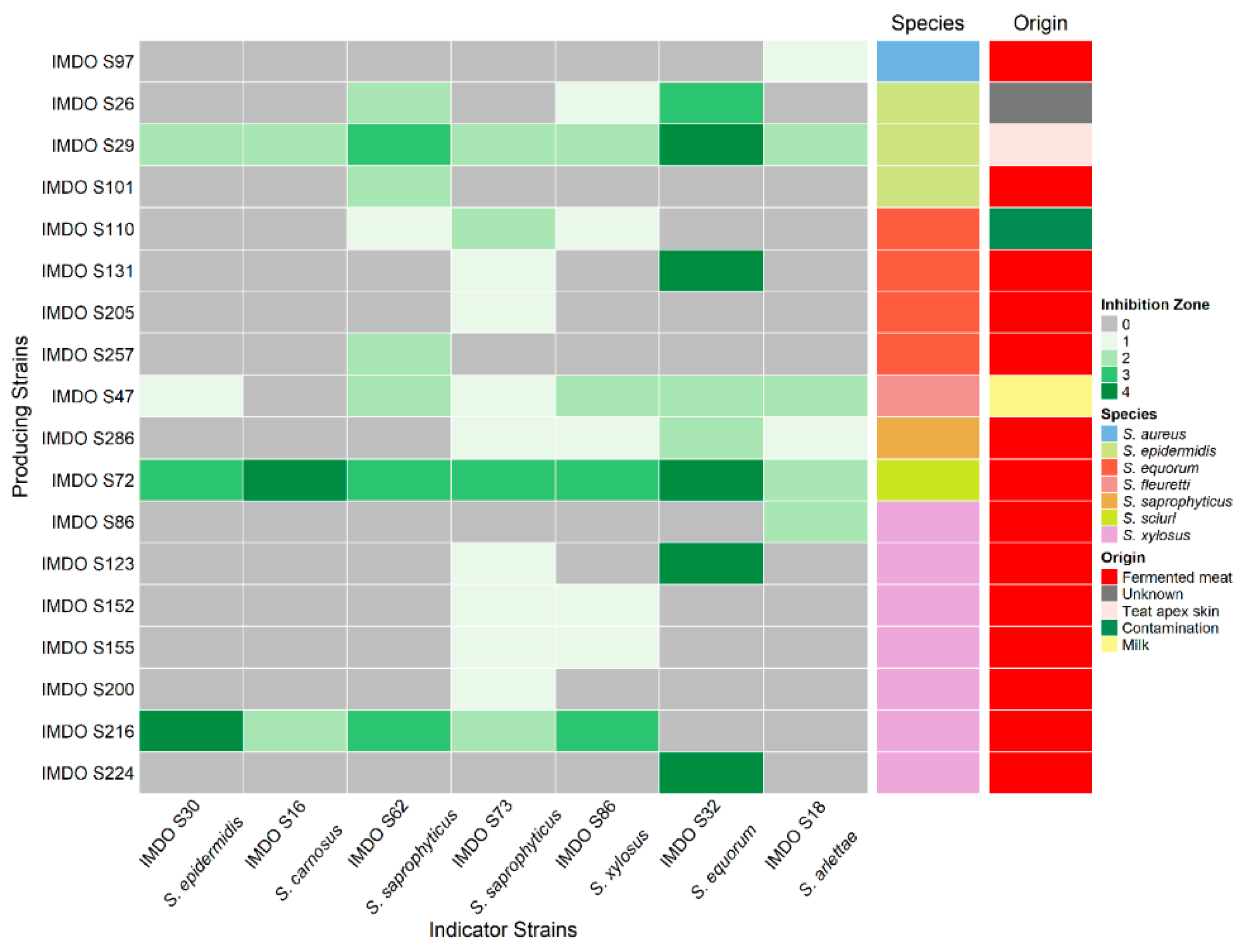
3.1. Prevalence of Antibacterial Activity in Staphylococci
3.2. Antibacterial Activity by Staphylococcus sciuri IMDO-S72
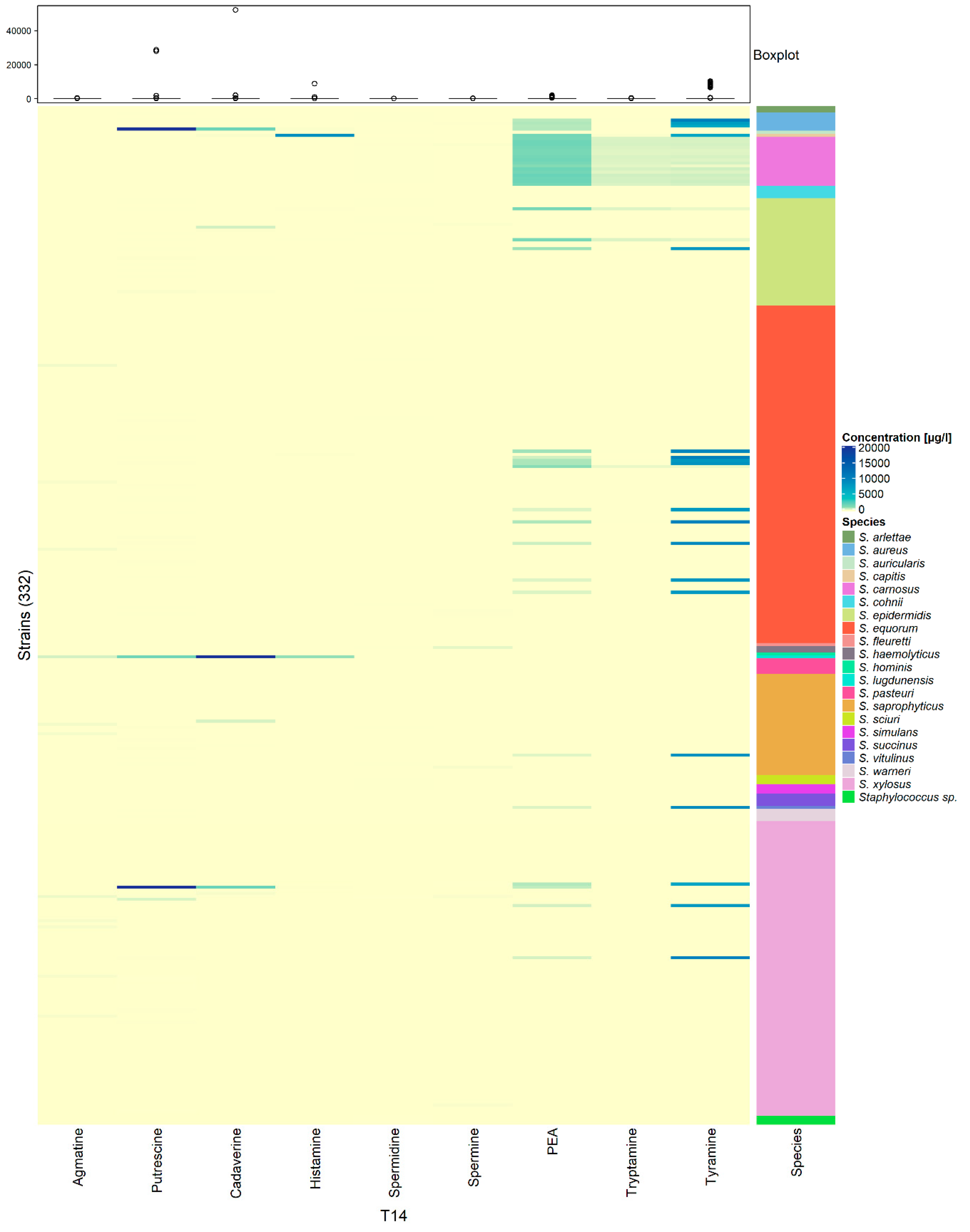
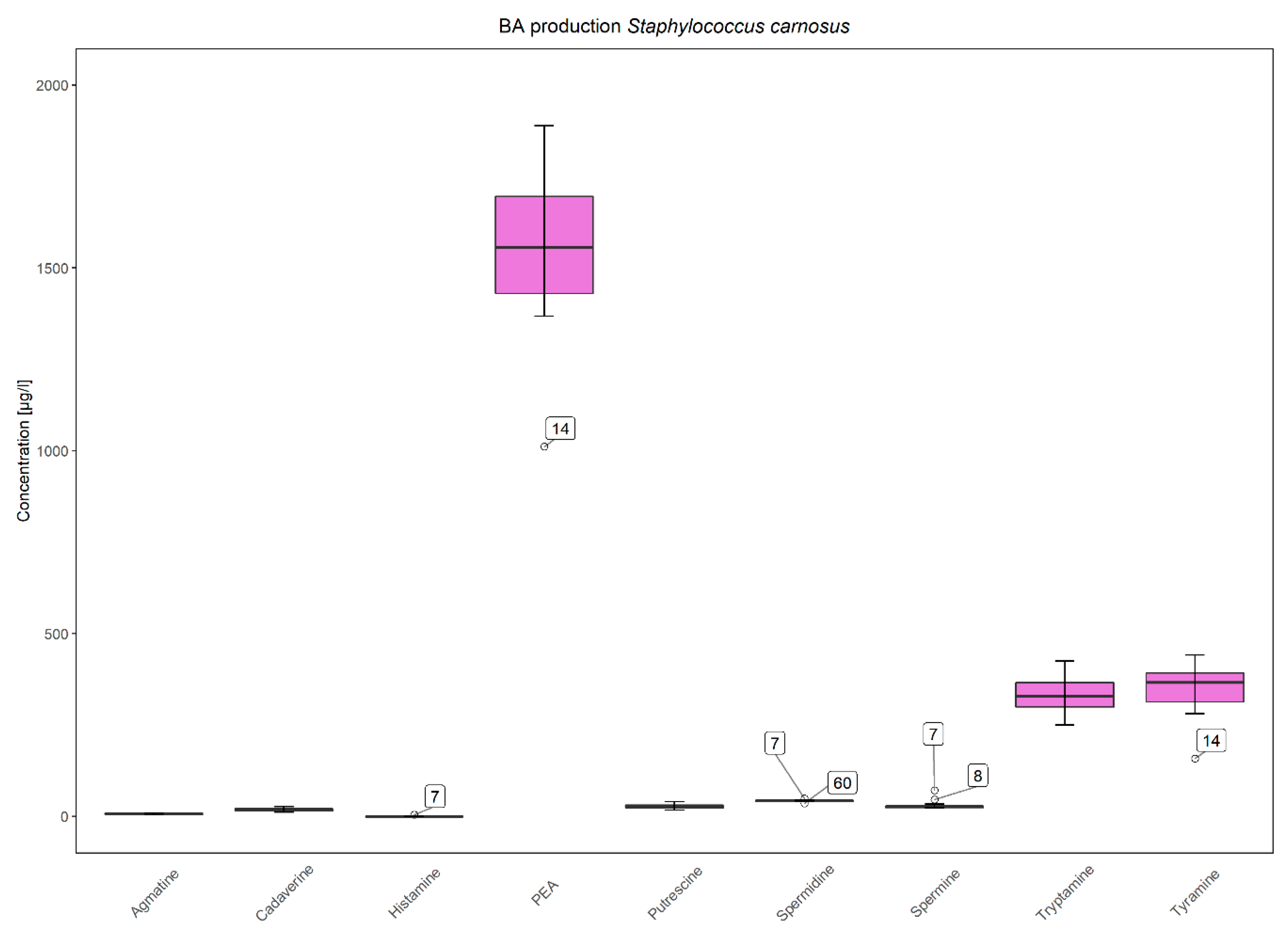
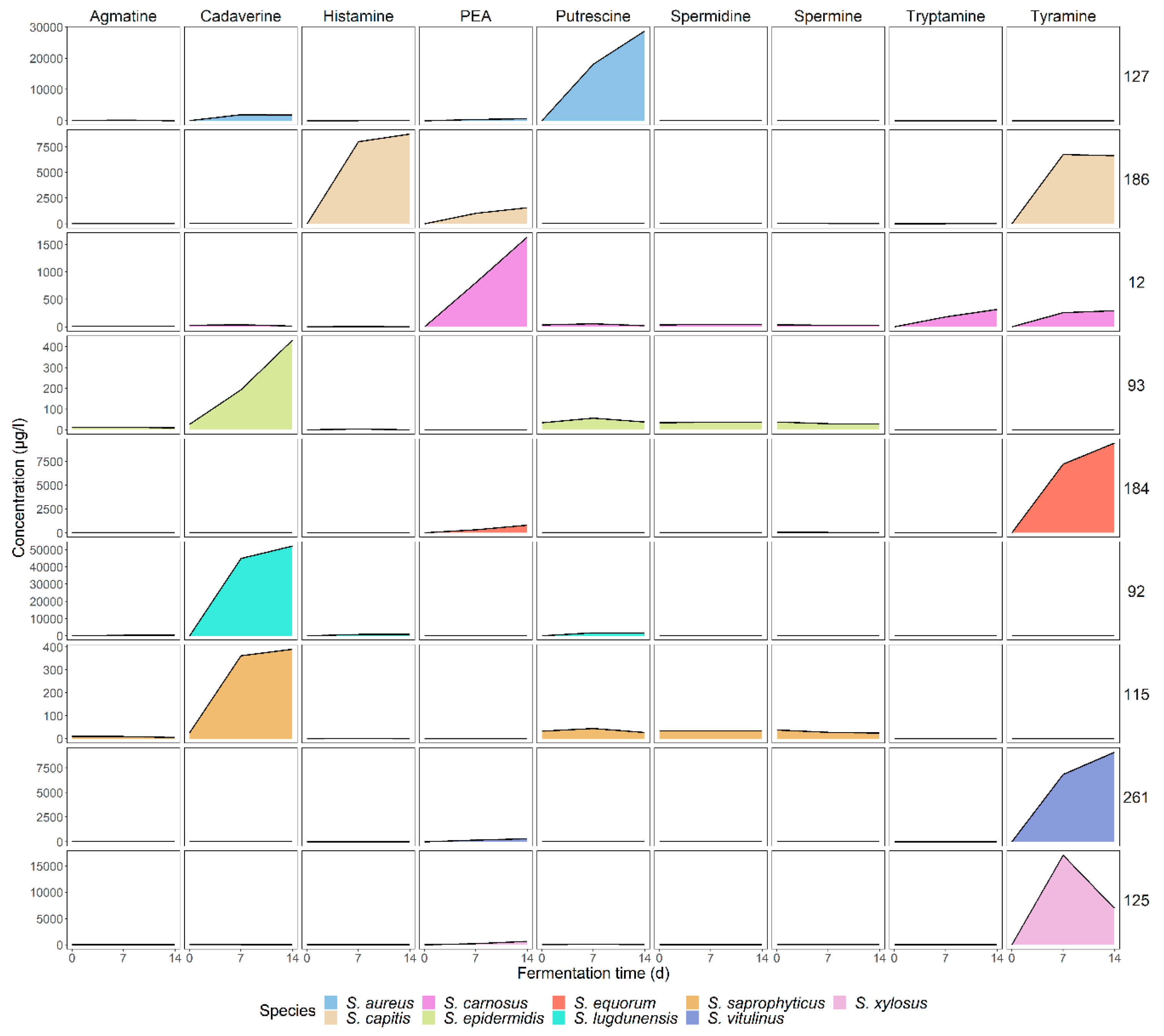
3.3. Low and Strain-Dependent Biogenic Amine Production in Staphylococci, Marked by Simultaneous Production of Tyramine and β-Phenylethylamine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leroy, F.; Geyzen, A.; Janssens, M.; De Vuyst, L.; Scholliers, P. Meat fermentation at the crossroads of innovation and tradition: A historical outlook. Trends Food Sci. Technol. 2013, 31, 130–137. [Google Scholar] [CrossRef]
- De Mey, E.; De Klerck, K.; De Maere, H.; Dewulf, L.; Derdelinckx, G.; Peeters, M.-C.; Fraeye, I.; Vander Heyden, Y.; Paelinck, H. The occurrence of N-nitrosamines, residual nitrite and biogenic amines in commercial dry fermented sausages and evaluation of their occasional relation. Meat Sci. 2014, 96, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Majou, D.; Christieans, S. Mechanisms of the bactericidal effects of nitrate and nitrite in cured meats. Meat Sci. 2018, 145, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Sebranek, J.G.; Bacus, J.N. Cured meat products without direct addition of nitrate or nitrite: What are the issues? Meat Sci. 2007, 77, 136–147. [Google Scholar] [CrossRef]
- Honikel, K.-O. The use and control of nitrate and nitrite for the processing of meat products. Meat Sci. 2008, 78, 68–76. [Google Scholar] [CrossRef]
- Hospital, X.F.; Hierro, E.; Stringer, S.; Fernández, M. A study on the toxigenesis by Clostridium botulinum in nitrate and nitrite-reduced dry fermented sausages. Int. J. Food Microbiol. 2016, 218, 66–70. [Google Scholar] [CrossRef]
- Lund, B.M.; Peck, M.W. Clostridium botulinum. In Guide to Foodborne Pathogens, 2nd ed.; Labbé, R.G., Garcia, S., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 91–111. [Google Scholar] [CrossRef]
- Ravyts, F.; Barbuti, S.; Frustoli, M.A.; Parolari, G.; Saccani, G.; De Vuyst, L.; Leroy, F. Competitiveness and antibacterial potential of bacteriocin-producing starter cultures in different types of fermented sausages. J. Food Protect. 2008, 71, 1817–1827. [Google Scholar] [CrossRef]
- Di Gioia, D.; Mazzola, G.; Nikodinoska, I.; Aloisio, I.; Langerholc, T.; Rossi, M.; Raimondi, S.; Melero, B.; Rovira, J. Lactic acid bacteria as protective cultures in fermented pork meat to prevent Clostridium spp. growth. Int. J. Food Microbiol. 2016, 235, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Mainar, M.; Xhaferi, R.; Samapundo, S.; Devlieghere, F.; Leroy, F. Opportunities and limitations for the production of safe fermented meats without nitrate and nitrite using an antibacterial Staphylococcus sciuri starter culture. Food Control 2016, 69, 267–274. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Hugas, M. Bacteriocinogenic lactic acid bacteria for the biopreservation of meat and meat products. Meat Sci. 1998, 49, S139–S150. [Google Scholar] [CrossRef]
- Ammor, S.; Tauveron, G.; Dufour, E.; Chevallier, I. Antibacterial activity of lactic acid bacteria against spoilage and pathogenic bacteria isolated from the same meat small-scale facility: 1—Screening and characterization of the antibacterial compounds. Food Control 2006, 17, 454–461. [Google Scholar] [CrossRef]
- Hanlin, M.B.; Kalchayanand, N.; Ray, P.; Ray, B. Bacteriocins of lactic acid bacteria in combination have greater antibacterial activity. J. Food Protect. 1993, 56, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, A.; Gobbetti, M.; Smacchi, E. Antibacterial activity of sourdough lactic acid bacteria: Isolation of a bacteriocin-like inhibitory substance from Lactobacillus sanfrancisco C57. Food Microbiol. 1996, 13, 447–456. [Google Scholar] [CrossRef]
- Ghrairi, T.; Manai, M.; Berjeaud, J.M.; Frère, J. Antilisterial activity of lactic acid bacteria isolated from rigouta, a traditional Tunisian cheese. J. Appl. Microbiol. 2004, 97, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Alves, V.F.; Martinez, R.C.R.; Lavrador, M.A.S.; De Martinis, E.C.P. Antilisterial activity of lactic acid bacteria inoculated on cooked ham. Meat Sci. 2006, 74, 623–627. [Google Scholar] [CrossRef]
- Albano, H.; Oliveira, M.; Aroso, R.; Cubero, N.; Hogg, T.; Teixeira, P. Antilisterial activity of lactic acid bacteria isolated from “Alheiras” (traditional Portuguese fermented sausages): In situ assays. Meat Sci. 2007, 76, 796–800. [Google Scholar] [CrossRef]
- Fontana, C.; Cocconcelli, P.S.; Vignolo, G.; Saavedra, L. Occurrence of antilisterial structural bacteriocins genes in meat borne lactic acid bacteria. Food Control 2015, 47, 53–59. [Google Scholar] [CrossRef]
- Sánchez Mainar, M.; Stavropoulou, D.A.; Leroy, F. Exploring the metabolic heterogeneity of coagulase-negative staphylococci to improve the quality and safety of fermented meats: A review. Int. J. Food Microbiol. 2017, 247, 24–37. [Google Scholar] [CrossRef]
- Even, S.; Leroy, S.; Charlier, C.; Ben Zakour, N.; Chacornac, J.-P.; Lebert, I.; Jamet, E.; Desmonts, M.-H.; Coton, E.; Pochet, S.; et al. Low occurrence of safety hazards in coagulase-negative staphylococci isolated from fermented foodstuffs. Int. J. Food Microbiol. 2010, 139, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ruaro, A.; Andrighetto, C.; Torriani, S.; Lombardi, A. Biodiversity and characterization of indigenous coagulase-negative staphylococci isolated from raw milk and cheese of North Italy. Food Microbiol. 2013, 34, 106–111. [Google Scholar] [CrossRef]
- Marcobal, A.; de las Rivas, B.; Landete, J.M.; Tabera, L.; Munoz, R. Tyramine and phenylethylamine biosynthesis by food bacteria. Crit. Rev. Food Sci. Nutr. 2012, 52, 448–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jairath, G.; Singh, P.K.; Dabur, R.S.; Rani, M.; Chaudhari, M. Biogenic amines in meat and meat products and its public health significance: A review. J. Food Sci. Technol. 2015, 52, 6835–6846. [Google Scholar] [CrossRef]
- Pircher, A.; Bauer, F.; Paulsen, P. Formation of cadaverine, histamine, putrescine and tyramine by bacteria isolated from meat, fermented sausages and cheeses. Eur. Food Res. Technol. 2007, 226, 225–231. [Google Scholar] [CrossRef]
- Suzzi, G.; Gardini, F. Biogenic amines in dry fermented sausages: A review. Int. J. Food Microbiol. 2003, 88, 41–54. [Google Scholar] [CrossRef]
- del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernandez, M.; Martin, M.C.; Alvarez, M.A. The biogenic amines putrescine and cadaverine show in vitro cytotoxicity at concentrations that can be found in foods. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bover-Cid, S.; Hugas, M.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. Amino acid-decarboxylase activity of bacteria isolated from fermented pork sausages. Int. J. Food Microbiol. 2001, 66, 185–189. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Torriani, S.; Gatto, V.; Tofalo, R.; Suzzi, G.; Belletti, N.; Gardini, F. Relationships between microbial population dynamics and putrescine and cadaverine accumulation during dry fermented sausage ripening. J. Appl. Microbiol. 2009, 106, 1397–1407. [Google Scholar] [CrossRef]
- Curiel, J.A.; Ruiz-Capillas, C.; de las Rivas, B.; Carrascosa, A.V.; Jiménez-Colmenero, F.; Muñoz, R. Production of biogenic amines by lactic acid bacteria and enterobacteria isolated from fresh pork sausages packaged in different atmospheres and kept under refrigeration. Meat Sci. 2011, 88, 368–373. [Google Scholar] [CrossRef] [Green Version]
- de las Rivas, B.; Ruiz-Capillas, C.; Carrascosa, A.V.; Curiel, J.A.; Jimenez-Colmenero, F.; Munoz, R. Biogenic amine production by Gram-positive bacteria isolated from Spanish dry-cured “chorizo” sausage treated with high pressure and kept in chilled storage. Meat Sci. 2008, 80, 272–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komprda, T.; Sladkova, P.; Petirova, E.; Dohnal, V.; Burdychova, R. Tyrosine- and histidine-decarboxylase positive lactic acid bacteria and enterococci in dry fermented sausages. Meat Sci. 2010, 86, 870–877. [Google Scholar] [CrossRef]
- Durlu-Ozkaya, F.; Ayhan, K.; Vural, N. Biogenic amines produced by Enterobacteriaceae isolated from meat products. Meat Sci. 2001, 58, 163–166. [Google Scholar] [CrossRef]
- Lazaro, C.A.; Conte-Junior, C.A.; Canto, A.C.; Guerra Monteiro, M.L.; Costa-Lima, B.; da Cruz, A.G.; Marsico, E.T.; Franco, R.M. Biogenic amines as bacterial quality indicators in different poultry meat species. LWT Food Sci. Technol. 2015, 60, 15–21. [Google Scholar] [CrossRef]
- Li, L.; Wen, X.; Wen, Z.; Chen, S.; Wang, L.; Wei, X. Evaluation of the biogenic amines formation and degradation abilities of Lactobacillus curvatus from Chinese bacon. Front. Microbiol. 2018, 9, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masson, F.; Talon, R.; Montel, M.C. Histamine and tyramine production by bacteria from meat products. Int. J. Food Microbiol. 1996, 32, 199–207. [Google Scholar] [CrossRef]
- Landeta, G.; Curiel, J.A.; Carrascosa, A.V.; Munoz, R.; de las Rivas, B. Characterization of coagulase-negative staphylococci isolated from Spanish dry cured meat products. Meat Sci. 2013, 93, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Landeta, G.; de las Rivas, B.; Carrascosa, A.V.; Muñoz, R. Screening of biogenic amine production by coagulase-negative staphylococci isolated during industrial Spanish dry-cured ham processes. Meat Sci. 2007, 77, 556–561. [Google Scholar] [CrossRef] [Green Version]
- Marty, E.; Bodenmann, C.; Buchs, J.; Hadorn, R.; Eugster-Meier, E.; Lacroix, C.; Meile, L. Prevalence of antibiotic resistance in coagulase-negative staphylococci from spontaneously fermented meat products and safety assessment for new starters. Int. J. Food Microbiol. 2012, 159, 74–83. [Google Scholar] [CrossRef]
- Seitter, M.; Geng, B.; Hertel, C. Binding to extracellular matrix proteins and formation of biogenic amines by food-associated coagulase-negative staphylococci. Int. J. Food Microbiol. 2011, 145, 483–487. [Google Scholar] [CrossRef]
- Simonova, M.; Strompfova, V.; Marcinakova, M.; Laukovda, A.; Vesterlund, S.; Moratalla, M.L.; Bover-Cid, S.; Vidal-Carou, C. Characterization of Staphylococcus xylosus and Staphylococcus carnosus isolated from Slovak meat products. Meat Sci. 2006, 73, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulou, D.A.; Borremans, W.; De Vuyst, L.; De Smet, S.; Leroy, F. Amino acid conversions by coagulase-negative staphylococci in a rich medium: Assessment of inter- and intraspecies heterogeneity. Int. J. Food Microbiol. 2015, 212, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Coton, E.; Mulder, N.; Coton, M.; Pochet, S.; Trip, H.; Lolkema, J.S. Origin of the putrescine-producing ability of the coagulase-negative bacterium Staphylococcus epidermidis 2015B. Appl. Environ. Microbiol. 2010, 76, 5570–5576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braem, G.; De Vliegher, S.; Supré, K.; Haesebrouck, F.; Leroy, F.; De Vuyst, L. (GTG)5-PCR fingerprinting for the classification and identification of coagulase-negative Staphylococcus species from bovine milk and teat apices: A comparison of type strains and field isolates. Vet. Microbiol. 2011, 147, 67–74. [Google Scholar] [CrossRef]
- Tagg, J.R.; Bannister, L.V. “Fingerprinting” β-haemolytic streptococci by their production of and sensitivity to bacteriocine-like inhibitors. J. Med. Microbiol. 1979, 12, 397–411. [Google Scholar] [CrossRef]
- Lee, N.-K.; Jun, S.-A.; Ha, J.-U.; Paik, H.-D. Screening and characterization of bacteriocinogenic lactic acid bacteria from Jeot-Gal, a Korean fermented fish food. J. Microbiol. Biotechnol. 2000, 10, 423–428. [Google Scholar]
- Clauwers, C.; Vanoirbeek, K.; Delbrassinne, L.; Michiels, C.W. Construction of nontoxigenic mutants of nonproteolytic Clostridium botulinum NCTC 11219 by insertional mutagenesis and gene replacement. Appl. Environ. Microbiol. 2016, 82, 3100–3108. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: New York, NY, USA, 2016; p. 260. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Stavropoulou, D.A.; De Maere, H.; Berardo, A.; Janssens, B.; Filippou, P.; De Vuyst, L.; De Smet, S.; Leroy, F. Species pervasiveness within the group of coagulase-negative staphylococci associated with meat fermentation is modulated by pH. Front. Microbiol. 2018, 9, 2232. [Google Scholar] [CrossRef] [Green Version]
- Stavropoulou, D.A.; Van Reckem, E.; De Smet, S.; De Vuyst, L.; Leroy, F. The narrowing down of inoculated communities of coagulase-negative staphylococci in fermented meat models is modulated by temperature and pH. Int. J. Food Microbiol. 2018, 274, 52–59. [Google Scholar] [CrossRef]
- Coppola, S.; Mauriello, G.; Aponte, M.; Moschetti, G.; Villani, F. Microbial succession during ripening of Naples-type salami, a southern Italian fermented sausage. Meat Sci. 2000, 56, 321–329. [Google Scholar] [CrossRef]
- Stavropoulou, D.A.; De Maere, H.; Berardo, A.; Janssens, B.; Filippou, P.; De Vuyst, L.; De Smet, S.; Leroy, F. Pervasiveness of Staphylococcus carnosus over Staphylococcus xylosus is affected by the level of acidification within a conventional meat starter culture set-up. Int. J. Food Microbiol. 2018, 274, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus epidermidis—The “accidental” pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raz, R.; Colodner, R.; Kunin, C.M. Who Are You—Staphylococcus saprophyticus? Clin. Infect. Dis. 2005, 40, 896–898. [Google Scholar] [CrossRef] [PubMed]
- Janek, D.; Zipperer, A.; Kulik, A.; Krismer, B.; Peschel, A. High frequency and diversity of antimicrobial activities produced by nasal Staphylococcus strains against bacterial competitors. PLoS Pathog. 2016, 12, e1005812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandiford, S.; Upton, M. Identification, characterization, and recombinant expression of epidermicin NI01, a novel unmodified bacteriocin produced by Staphylococcus epidermidis that displays potent activity against staphylococci. Antimicrob. Agents Chemother. 2012, 56, 1539–1547. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, J.D.S.; Coelho, M.L.V.; Ceotto, H.; Potter, A.; Fleming, L.R.; Salehian, Z.; Nes, I.F.; Bastos, M.C.F. Genes involved in immunity to and secretion of aureocin A53, an atypical class II bacteriocin produced by Staphylococcus aureus A53. J. Bacteriol. 2012, 194, 875–883. [Google Scholar] [CrossRef] [Green Version]
- Navaratna, M.A.D.B.; Sahl, H.-G.; Tagg, J.R. Identification of genes encoding two-component lantibiotic production in Staphylococcus aureus C55 and other phage group II S. aureus strains and demonstration of an association with the exfoliative toxin B gene. Infect. Immun. 1999, 67, 4268–4271. [Google Scholar] [CrossRef] [Green Version]
- Bennallack, P.R.; Burt, S.R.; Heder, M.J.; Robison, R.A.; Griffitts, J.S. Characterization of a novel plasmid-borne thiopeptide gene cluster in Staphylococcus epidermidis strain 115. J. Bacteriol. 2014, 196, 4344–4350. [Google Scholar] [CrossRef] [Green Version]
- Brito, M.A.V.P.; Somkuti, G.A.; Renye, J.A. Production of antilisterial bacteriocins by staphylococci isolated from bovine milk. J. Dairy Sci. 2011, 94, 1194–1200. [Google Scholar] [CrossRef]
- Rahmdel, S.; Shekarforoush, S.S.; Hosseinzadeh, S.; Torriani, S.; Gatto, V. Antimicrobial spectrum activity of bacteriocinogenic Staphylococcus strains isolated from goat and sheep milk. J. Dairy Sci. 2019, 102, 2928–2940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carson, D.A.; Barkema, H.W.; Naushad, S.; De Buck, J. Bacteriocins of non-aureus staphylococci isolated from bovine milk. Appl. Environ. Microbiol. 2017, 83, e01015-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, M.A.; Somkuti, G.A.; Renye, J.A. Isolation of bacteriocin-producing staphylococci from Brazilian cheese. J. Food Saf. 2011, 31, 365–370. [Google Scholar] [CrossRef]
- Hong, J.; Quan, L.-H.; Heu, S.; Jung, K.S.; Han, S.-W.; Moon, E.; Roh, E. A new antimicrobial substance produced by Staphylococcus pasteuri isolated from vegetables. Food Sci. Biotechnol. 2014, 23, 983–990. [Google Scholar] [CrossRef]
- Braem, G.; Stijlemans, B.; Van Haken, W.; De Vliegher, S.; De Vuyst, L.; Leroy, F. Antibacterial activities of coagulase-negative staphylococci from bovine teat apex skin and their inhibitory effect on mastitis-related pathogens. J. Appl. Microbiol. 2014, 116, 1084–1093. [Google Scholar] [CrossRef]
- Nascimento, J.D.S.; Fagundes, P.C.; Brito, M.A.V.P.; dos Santos, K.R.N.; Bastos, M.C.F. Production of bacteriocins by coagulase-negative staphylococci involved in bovine mastitis. Vet. Microbiol. 2005, 106, 61–71. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9, eaah4680. [Google Scholar] [CrossRef] [Green Version]
- Kleerebezem, M. Quorum sensing control of lantibiotic production; nisin and subtilin autoregulate their own biosynthesis. Peptides 2004, 25, 1405–1414. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Quadri, L.E. Peptide pheromone-dependent regulation of antimicrobial peptide production in Gram-positive bacteria: A case of multicellular behavior. Peptides 2001, 22, 1579–1596. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Kuipers, O.P.; de Vos, W.M.; Stiles, M.E.; Quadri, L.E.N. A two-component signal-transduction cascade in Carnobacterium piscicola LV17B: Two signaling peptides and one sensor-transmitter. Peptides 2001, 22, 1597–1601. [Google Scholar] [CrossRef] [Green Version]
- Leroy, F.; de Vuyst, L. Temperature and pH conditions that prevail during fermentation of sausages are optimal for production of the antilisterial bacteriocin sakacin K. Appl. Environ. Microbiol. 1999, 65, 974–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sarrau, B.; Clavel, T.; Clerté, C.; Carlin, F.; Giniès, C.; Nguyen-The, C. Influence of anaerobiosis and low temperature on Bacillus cereus growth, metabolism, and membrane properties. Appl. Environ. Microbiol. 2012, 78, 1715–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kröckel, L. The role of lactic acid bacteria in safety and flavour development of meat and meat products. In Lactic Acid Bacteria—R & D for Food, Health and Livestock Purposes; Kongo, J.M., Ed.; IntechOpen Limited: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Peypoux, F.; Bonmatin, J.M.; Wallach, J. Recent trends in the biochemistry of surfactin. Appl. Microbiol. Biotechnol. 1999, 51, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Souto, G.I.; Correa, O.S.; Montecchia, M.S.; Kerber, N.L.; Pucheu, N.L.; Bachur, M.; García, A.F. Genetic and functional characterization of a Bacillus sp. strain excreting surfactin and antifungal metabolites partially identified as iturin-like compounds. J. Appl. Microbiol. 2004, 97, 1247–1256. [Google Scholar] [CrossRef]
- Tran, T.T.; Munita, J.M.; Arias, C.A. Mechanisms of drug resistance: Daptomycin resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 32–53. [Google Scholar] [CrossRef] [Green Version]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [Green Version]
- Nemeghaire, S.; Argudín, M.A.; Feßler, A.T.; Hauschild, T.; Schwarz, S.; Butaye, P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Vet. Microbiol. 2014, 171, 342–356. [Google Scholar] [CrossRef]
- Zeman, M.; Mašlaňová, I.; Indráková, A.; Šiborová, M.; Mikulášek, K.; Bendíčková, K.; Plevka, P.; Vrbovská, V.; Zdráhal, Z.; Doškař, J.; et al. Staphylococcus sciuri bacteriophages double-convert for staphylokinase and phospholipase, mediate interspecies plasmid transduction, and package mecA gene. Sci. Rep. 2017, 7, 46319. [Google Scholar] [CrossRef]
- Lucas, P.M.; Wolken, W.A.M.; Claisse, O.; Lolkema, J.S.; Lonvaud-Funel, A. Histamine-producing pathway encoded on an unstable plasmid in Lactobacillus hilgardii 0006. Appl. Environ. Microbiol. 2005, 71, 1417–1424. [Google Scholar] [CrossRef] [Green Version]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Mainar, M.; Weckx, S.; Leroy, F. Coagulase-negative staphylococci favor conversion of arginine into ornithine despite a widespread genetic potential for nitric oxide synthase activity. Appl. Environ. Microbiol. 2014, 80, 7741–7751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcobal, Á.; de las Rivas, B.; García-Moruno, E.; Muñoz, R. The tyrosine decarboxylation test does not differentiate Enterococcus faecalis from Enterococcus faecium. Syst. Appl. Microbiol. 2004, 27, 423–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aymerich, T.; Martín, B.; Garriga, M.; Vidal-Carou, M.C.; Bover-Cid, S.; Hugas, M. Safety properties and molecular strain typing of lactic acid bacteria from slightly fermented sausages. J. Appl. Microbiol. 2006, 100, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Pardo, I.; Ferrer, S. Tyramine and phenylethylamine production among lactic acid bacteria isolated from wine. Int. J. Food Microbiol. 2007, 115, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Gardini, F.; Bover-Cid, S.; Tofalo, R.; Belletti, N.; Gatto, V.; Suzzi, G.; Torriani, S. Modeling the aminogenic potential of Enterococcus faecalis EF37 in dry fermented sausages through chemical and molecular approaches. Appl. Environ. Microbiol. 2008, 74, 2740–2750. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of biogenic amines on food quality and safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- de las Rivas, B.; Rodríguez, H.; Carrascosa, A.V.; Muñoz, R. Molecular cloning and functional characterization of a histidine decarboxylase from Staphylococcus capitis. J. Appl. Microbiol. 2008, 104, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Drosinos, E.H.; Paramithiotis, S.; Kolovos, G.; Tsikouras, I.; Metaxopoulos, I. Phenotypic and technological diversity of lactic acid bacteria and staphylococci isolated from traditionally fermented sausages in Southern Greece. Food Microbiol. 2007, 24, 260–270. [Google Scholar] [CrossRef]
- Seitter, M.; Nerz, C.; Rosenstein, R.; Goetz, F.; Hertel, C. DNA microarray-based detection of genes involved in safety and technologically relevant properties of food associated coagulase-negative staphylococci. Int. J. Food Microbiol. 2011, 145, 449–458. [Google Scholar] [CrossRef]
- Martin, B.; Garriga, M.; Hugas, M.; Bover-Cid, S.; Veciana-Nogues, M.T.; Aymerich, T. Molecular, technological and safety characterization of Gram-positive catalase-positive cocci from slightly fermented sausages. Int. J. Food Microbiol. 2006, 107, 148–158. [Google Scholar] [CrossRef]
- Parente, E.; Martuscelli, M.; Gardini, F.; Grieco, S.; Crudele, M.A.; Suzzi, G. Evolution of microbial populations and biogenic amine production in dry sausages produced in Southern Italy. J. Appl. Microbiol. 2001, 90, 882–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bover-Cid, S.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. Effect of the interaction between a low tyramine-producing Lactobacillus and proteolytic staphylococci on biogenic amine production during ripening and storage of dry sausages. Int. J. Food Microbiol. 2001, 65, 113–123. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Compound | Ionisation Mode | SRM | |
|---|---|---|---|
| Transition (m/z) | CV (V)/CE (eV) | ||
| Agmatine | ES + | 131.00 > 72.02 | 15/15 |
| Cadaverine | ES + | 103.08 > 86.04 | 15/12 |
| Histamine | ES + | 112.06 > 95.02 | 18/15 |
| β-Phenylethylamine | ES + | 122.08 > 105.02 | 23/12 |
| Putrescine | ES + | 89.05 > 72.02 | 11/11 |
| Tryptamine | ES + | 161.13 > 144.03 | 10/11 |
| Tyramine | ES + | 138.09 > 121.04 | 10/12 |
| Spermidine | ES + | 146.00 > 72.05 | 20/12 |
| Spermine | ES + | 203.22 > 129.12 | 22/12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van der Veken, D.; Benhachemi, R.; Charmpi, C.; Ockerman, L.; Poortmans, M.; Van Reckem, E.; Michiels, C.; Leroy, F. Exploring the Ambiguous Status of Coagulase-Negative Staphylococci in the Biosafety of Fermented Meats: The Case of Antibacterial Activity Versus Biogenic Amine Formation. Microorganisms 2020, 8, 167. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8020167
Van der Veken D, Benhachemi R, Charmpi C, Ockerman L, Poortmans M, Van Reckem E, Michiels C, Leroy F. Exploring the Ambiguous Status of Coagulase-Negative Staphylococci in the Biosafety of Fermented Meats: The Case of Antibacterial Activity Versus Biogenic Amine Formation. Microorganisms. 2020; 8(2):167. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8020167
Chicago/Turabian StyleVan der Veken, David, Rafik Benhachemi, Christina Charmpi, Lore Ockerman, Marijke Poortmans, Emiel Van Reckem, Chris Michiels, and Frédéric Leroy. 2020. "Exploring the Ambiguous Status of Coagulase-Negative Staphylococci in the Biosafety of Fermented Meats: The Case of Antibacterial Activity Versus Biogenic Amine Formation" Microorganisms 8, no. 2: 167. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8020167