1. Introduction
The need for sustainable energy generation in recent years has increased interest in microbial processes for the production of fuels and value-added products. Concurrently, the search for finding alternative low-cost substrates has also increased and the use of substrates that create direct competition with the food resources has been discouraged [
1]. Commonly used substrates for microbial fermentation are simple sugars such as glucose and sucrose. Solid organic wastes (SOWs) such as food and paper waste, and agricultural wastes (such as corn stover) produced by anthropogenic activities, are abundant substrates that can be converted to intermediate or end-use bioproducts using microbial metabolism [
2,
3]. An enriched microbial consortium consisting of different microbial groups with diverse functions can degrade these SOWs into value-added end products or platform chemicals. Within the microbial consortium, hydrolytic enzymes produced by microbes convert the complex substrates into simpler sugars and organic acids. These intermediate organic acids often called as volatile fatty acids (VFAs) can be further utilized for microbial production of higher-value chemicals and thus can replace sugar substrates in industrial fermentations. VFAs are attractive substrates because they can be produced from a variety of organic wastes fermentation [
4]. Therefore, the use of VFAs as carbon sources seems to be a feasible strategy for cost-effective microbial productions. Acetic acid or acetate is the most desirable VFA as it can serve as an alternate and sole source of carbon for microorganisms and can be used at comparatively higher concentrations to replace sugar substrates. The global demand of acetic acid is 14.3 billion pounds per year and it can be employed in the production of photographic film (cellulose acetate), wood glue (polyvinyl acetate), and synthetic fibers (triacetate cellulose) used for textile, carpets, and cigarette filters, among other products [
5,
6]. In addition to anaerobic fermentation by the acetogenic microbial consortium, acetic acid is also produced using chemical processes (e.g., methanol carbonylation, ethylene oxidation, and alkane oxidation) [
7]. In the United States, the price of acetate (
$350−
$450 per ton) is lower than the price of glucose (
$500 per ton) [
8]; however, it is possible to obtain high titers of acetate inexpensively using renewable, and abundant feedstocks such as SOWs.
The microbial consortium carrying out thermophilic anaerobic digestion (TAD) comprise a significant fraction of acetogenic (acetate-producing) bacteria such as
Acetomicrobium, Clostridium, and
Acetobacter sp. which can convert SOW to acetate more rapidly and without any chemical or physical pretreatment [
9]. The use of a TAD system is advantageous because the high-temperature fermentation process increases acetate end product formation and prevents the formation of other VFAs such as butyrate [
10]. Thermophilic bacteria are also reported to simultaneously utilize mixed hexose and pentose sugars which could be beneficial when the SOW used is a lignocellulosic substrate [
10,
11]. Thermophilic conditions have additional benefits of high substrate degradation rate, pathogen removal, and efficient heat utilization of SOWs. The thermophilic microbial consortium also harbors distinct microbial species that possess metabolic functions related to biomass degradation and utilization [
12,
13]. Therefore, a TAD system fed with SOWs can be rewired to an acetate-producing system by inhibition of methanogenesis. This results in simultaneous enrichment of a thermophilic acetogenic consortium (TAC) which primarily produces acetate.
Most prior studies have optimized the factors leading to high total VFA productions [
4,
14,
15] for fuels and chemical applications. All these research endeavors are important considering that raw material alone constitutes 40%–80% of biofuel production costs. Biofuels made from VFAs derived from waste organic biomass potentially offer significant economic advantages [
16]. To reduce production costs, some pure culture studies have been conducted and reviewed using SOWs [
17,
18,
19]. However, the literature is limited regarding thermophilic anaerobic fermentation processes using mixed consortia that would use SOWs and produce a higher fraction of acetate (in the VFA pool). Though the control of metabolite is difficult in mixed culture fermentations compared to pure culture counterparts, however, mixed culture fermentations are advantageous due to the absence of sterilization requirements, a stable operation when designed on proper ecological selection principles, the potential for stable continuous operation, and adaptive capacity to variations in feed or conditions [
20]. If optimized, mixed culture fermentation processes have a great chance to outcompete pure culture fermentations [
17]. In addition, the production of a higher fraction of a single VFA (i.e., acetate in this study) is desirable for simplifying downstream recovery processes. Among the different VFAs, acetic acid has the highest market size, with 3,500,000 tons/y and a price per ton of
$800 [
21]. The production of predominant acetate together with methane in extreme-thermophilic (70 °C) mixed culture fermentation was first studied by Zhang et al. [
12]. However, they used pure glucose as a substrate to achieve >90% acetate in the fermentate’s metabolite pool. In contrast, our study uses completely SOWs to produce acetate-rich fermentate. Knowledge of environmental factors affecting the acetate production from thermophilic acetogenic consortium is of considerable importance for the efficient conversion of SOWs into acetic acid at an industrial scale. Therefore, this work aims at developing a microbial consortium and maximizing acetate production using two SOWs—cafeteria (food and paper) wastes and corn stover—as a substrate. Initial screening experiments were conducted using two-level factorial design to screen for most influential parameters affecting the conversion of cafeteria and corn stover waste into acetic acid. Further, optimization of the process conditions, i.e., incubation temperature, pH, and C:N ratio, was done to understand their impact on acetic acid production at individual and interactive levels using statistical optimization with response surface methodology (RSM). Statistical optimization is a very effective tool in determining the process parameter values for increasing the desired product yield because it takes into account the interaction effects of the process parameters tested [
22,
23].
Various industrially relevant microorganisms can naturally utilize acetic acid and convert it into valuable bioproducts [
24,
25,
26,
27,
28]. Advancement in the field of microbial genetics and metabolic engineering has furthered the range of bioproducts synthesized and substrates that can be utilized by microorganisms [
29,
30,
31]. Research efforts have also been directed toward increasing the substrate uptake and its utilization. This is important to achieve higher bioproduct yields and productivity and, therefore, can improve the overall economics of industrial microbial processes. An industrially relevant host organism is the oleaginous yeast—
Yarrowia lipolytica—which has gained a lot of attention recently for the production of lipids using various pure substrate (e.g., glucose, fructose, lactose, sucrose, glycerol, and xylose) and raw feedstocks (e.g., olive oil mill waste, whey, waste cooking oil, and animal fats) [
26,
32]. Oleaginous yeasts have a distinctive ability to convert certain organic acids directly to acetyl-CoA—the central intermediate of lipid biosynthesis—by acetyl coenzyme A synthetase. Acetyl-CoA is subsequently utilized in fatty acid (FA) synthesis and results in lipid accumulation [
33]. Recent studies have also explored the potential of native and engineered
Y.lipolytica to synthesize products other than lipids including itaconic acid [
34], erythritol [
35], citric acid [
36], carotenoids [
37], and polyhydroxyalkanoates [
38]. So far, most studies on bioproduct synthesis by oleaginous microorganisms like
Yarrowia lipolytica have been carried out with glucose as a carbon source. Published literature on volatile fatty acids (derived from different SOWs) as a sole carbon source for this yeast is scarce and no previous study has attempted to convert these low-cost volatile fatty acids into itaconic acid (a dicarboxylic acid), which is industrial chemical serving as a precursor of polymers used in plastics, adhesives, and coatings. It is also one of the “top 12” building block chemicals listed by the United States Department of Energy [
39]. We are currently engineering a
Yarrowia lipolytica strain to increase its ability to uptake acetate as well as upregulate the metabolic pathways that increase acetate assimilation and divert its metabolic flux toward itaconic acid production (unpublished work). For the current study, we studied the growth of the wild-type
Y.lipolytica strain as a proof-of-concept for growing the engineered strain on the acetate-rich-fermentate in our future work. So, in addition to optimizing acetate production through anaerobic co-fermentation of cafeteria waste and corn stover, this study analyzes the feasibility of using this acetate-rich fermentate for growth of
Y.lipolytica.
2. Materials and Methods
2.1. Substrate
Mixed SOWs (corn stover and cafeteria wastes) were used as substrates for all optimization experiments. Corn stover was kindly provided by Dr. K. Muthukumarappan from South Dakota State University, Brookings, SD. It was crushed using a blender and sieved through sieve between 15- and 10- mm pores before being used as a substrate. Cafeteria wastes consisting of paper and food wastes were obtained from the South Dakota of Mines and Technology (SDSMT) cafeteria. The composition of cafeteria waste varies depending on many factors such as region and season. So to reduce some of this bias, cafeteria wastes were collected on 3 separate days over a span of 3 months, homogenized, and reduced in size using a mechanical mixer and stored at −20 °C until use.
2.2. Growth Medium and Experimental Set-Up
A modified basal anaerobic medium was used for inoculum development and comprised of (g/L): K2HPO4, 0.3; KH2PO4, 0.3; NaCl, 0.1; CaCl2, 0.05; NH4Cl, 1.0; MgCl2·6H2O 0.5; KCl, 0.3; cysteine HCl, 0.5; yeast extract, 0.05 and Na2S·9H2O, 0.003. NaHCO3 and Nitsch trace element solution were added to the medium to a final concentration of 20 mM and 2.5 mL per liter, respectively. All the experiments were performed in duplicates in 500 mL serum bottles with 200 mL working volume consisting of 10% v/v inoculum in anaerobic basal medium. The serum bottle reactors were autoclaved at 121 °C and 15 psi for 20 min prior to inoculation. A 50 mM solution of 2-bromoethanesulfonate (BES) was added to inhibit methanogenesis and enhance acetate production in different experimental runs according to the design of experiments. Following the addition of BES and inoculum, the serum bottle reactors were sealed with butyl rubber stoppers, crimped with aluminum crimps, and purged with nitrogen gas for 20 min to establish anaerobic conditions. They were incubated at different temperatures and in the presence or absence of shaking at 100 rpm according to the design of experiments.
2.3. Inoculum Development
The inoculum used was a combination of two different types of anaerobic sludge collected from the Wastewater Reclamation Plant (Rapid City, SD, USA) and from a lab-scale anaerobic digester fed with food, paper, and lignocellulosic wastes (at the Chemical and Biochemical Engineering Department, SDSMT, Rapid City, SD, USA). The inoculum was prepared by mixing the two anaerobic sludges in equal amounts (50/50
v/v) and adjusting the pH to 6 with 1 M HCl while flushing with N
2 to ensure anaerobic conditions. The inoculum was sieved through a 2 mm net after collection to remove large particles. The acetogenic consortia were developed and maintained in the lab using sub-culturing techniques as described by David and coll. (2018) [
40]. Briefly, one-gram volatile solids (VS) of mixed SOW (containing equal amounts of corn stover and cafeteria wastes) was added to the modified anaerobic basal medium and the bottles were incubated at 60 °C and 100 rpm. When acetate production reached a stable level, 10% (
v/v) of the actively growing anaerobic culture was transferred into fresh basal media (200 mL) containing another 1 g of mixed waste. After 10 serial transfers, a thermophilic acetogenic consortium was obtained that produced acetate using corn stover and cafeteria wastes as carbon and energy source. All experiments and chemical analyses were done in duplicates and the value presented is an average of the two values from the duplicate set up. The microbial community analysis of the thermophilic consortium while shifting from biogas production to primarily acetate production is being studied using Illumina sequencing and will be part of a separate manuscript (in preparation).
2.4. Analytical Methods
Total solids (TS) and volatile solids (VS) of the substrate and inoculum were measured according to APHA standard methods [
41]. The cellulose, hemicellulose, and lignin content were measured according to NREL analytical methods [
42]. Elemental analysis was done by Atlantic Microlab, Norcross, GA. Liquid samples from the serum bottle reactors were analyzed for three major volatile fatty acids (VFAs) (acetate, propionate, and butyrate) at regular time intervals over a course of 8 days or until the acetate production ceased increasing. The liquid samples were centrifuged at 10,000 rpm for 10 min to remove suspended solids and biomass prior to VFA measurement. VFA concentrations were determined using HPLC equipped with Aminex HPX-97 H column with 0.005 N sulfuric acid as the mobile phase. Acetate percent was calculated by dividing the concentration of acetate (mg/L) in the reactor liquid by the total concentration of the measured VFAs (i.e., the sum of acetate, propionate, and butyrate concentrations in mg/L). Carbon to nitrogen ratio was calculated using the following equation [
40,
43]:
where TOC = total organic carbon (%VS) and TN = total nitrogen (%VS)
2.5. Design of Experiment for Optimization of Acetate Production
2.5.1. Selection of Influential Factors
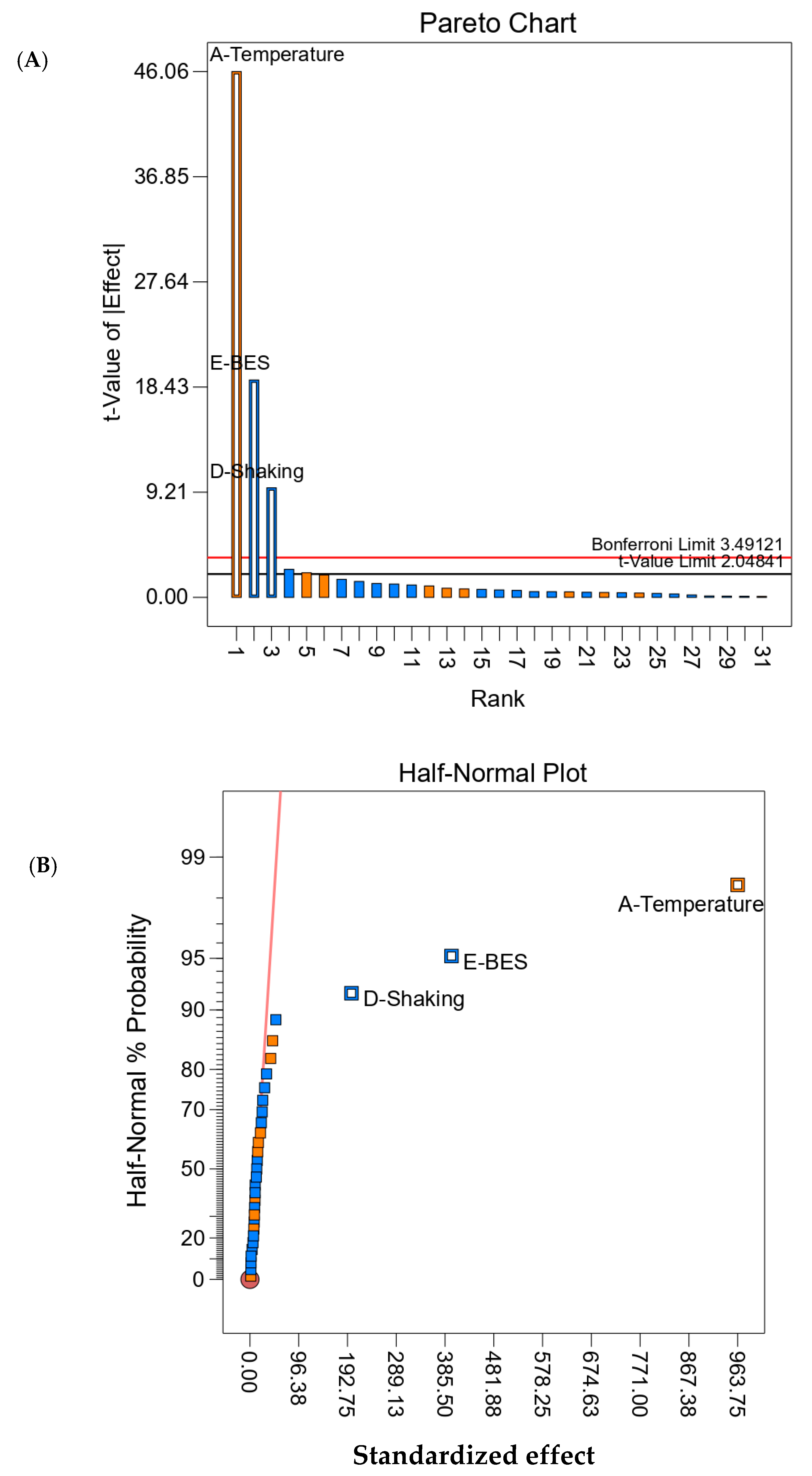
Ten factors that could influence acetate fermentation and growth of the acetogenic consortium were selected and tested using two-level factorial design in Design Expert Version 12.0.6.0 (Stat-Ease Inc., Minneapolis, Minnesota, MN, USA) statistical software. In a two-level factorial experiment each independent variable, i.e., a “factor” is investigated at two discrete possible values or “levels”. This is beneficial for estimating the main effects as well as the interaction effects of the factors tested. The 10 factors chosen are shown in
Table 1. Thirty-two different experimental runs were performed. The factors included physical parameters (temperature, shaking), chemical factors (pH and C:N ratio), and inorganic medium components (NaHCO
3, NH
4Cl, MgCl
2.6H
2O, and KCl). The impact of BES (methanogen inhibitor) and Na
2S.9H
2O (to give basal media reducing power) addition was also investigated. With a signal-to-noise ratio of 3, the power obtained for the statistical design was 99.9%. This was greater than the minimum recommended (>80%) for the effects that were to be detected. A low substrate load was used to dilute the inhibitory molecules that may be present in either cafeteria waste or corn stover, and that could potentially impact acetate fermentation by the newly developed consortium. Therefore, two-gram VS was used as a substrate for all the runs of this screening experiment. The factors that positively affected acetate concentration in the test runs were either selected for further optimization through RSM or were incorporated in all the RSM runs.
2.5.2. Response Surface Methodology for Statistical Optimization of Influential Factors for Acetate Production
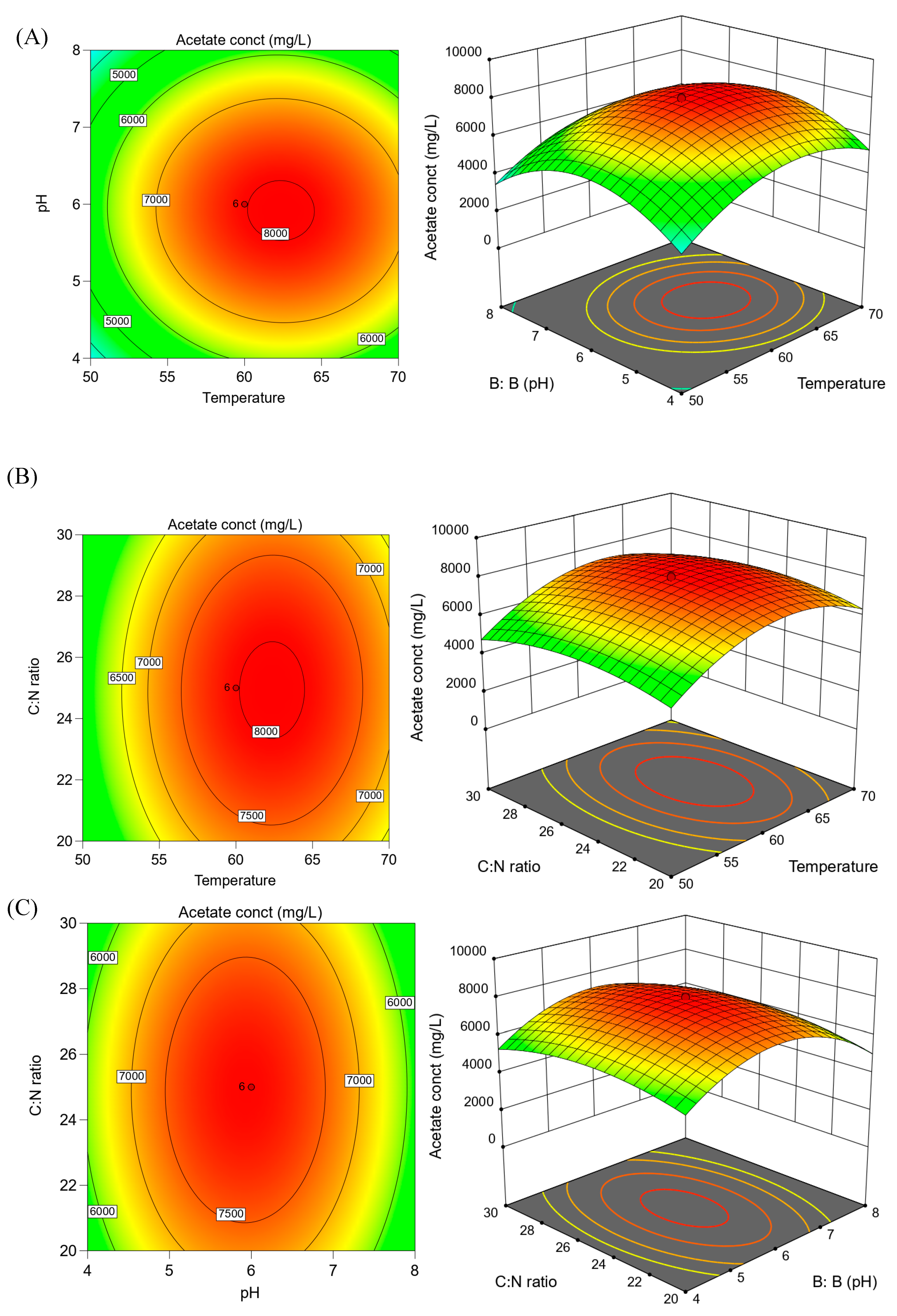
RSM is a widely used statistical tool for modeling or analyzing a process in which the parameters which affect the process are called independent variables, whereas the responses are called dependent variables [
44]. The objective for the RSM method is to optimize the response of interest which is affected by various variables, i.e., factors incorporated in the RSM design [
45]. Response surface methodology (RSM) was used to optimize factors having a significant effect on acetate production by anaerobic fermentation of mixed SOW. Three factors (independent variables)—temperature, pH, and C:N ratio—were chosen for statistical optimization of acetate production (dependent variable) in this study. A substrate load of 5 g VS was used for all the experimental runs. A 2
3 factorial central composite design with 14 non-center points and six replications at the center points (α = 1.68) yielded a total of 20 experimental runs. Each factor/variable was set to 5 levels: +α and −α (axial points), +1 and −1 (factorial points), and the center point (
Table 2). The Design Expert (Version 12.0.6.0) statistical software was used to analyze the results of the experimental design. Response was recorded in terms of acetate concentration in mg L
−1. Response data generated by the experimental runs are recorded in
Table 3. It was subjected to analysis of regression through RSM to solve multivariate equations. The behavior of the system was explained by the following quadratic equation:
where Y is the predicted response (acetate concentration, mgL
-1), X represents individual factors being considered in the design (in this case temperature, pH, and C:N ratio), β
0 is the intercept, β
i linear effect coefficient, β
ii is the quadratic effect coefficient, and β
ij is interaction effect coefficient. The significance of each effect was estimated by analysis of variance with
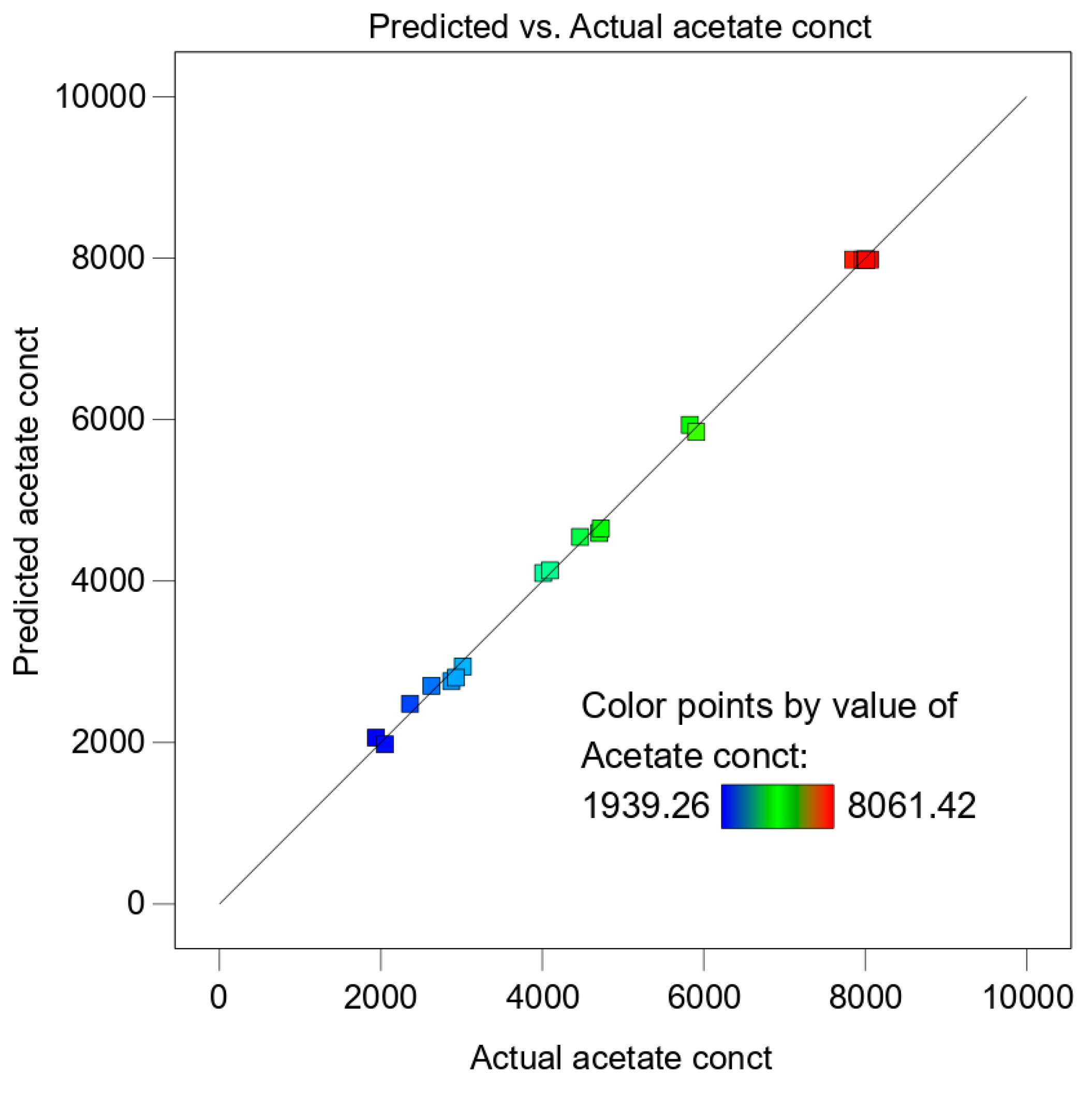
p-values < 0.05, i.e., above a 95% confidence interval. The Statistical Design Expert software was applied for multiple regression analysis and to create the plots of the obtained data. Each experiment was performed in triplicates and the data presented is mean of the triplicate values.
2.6. Growth of Yarrowia Lipolytica on SOW-Derived Acetate-Rich Anaerobic Fermentation Product
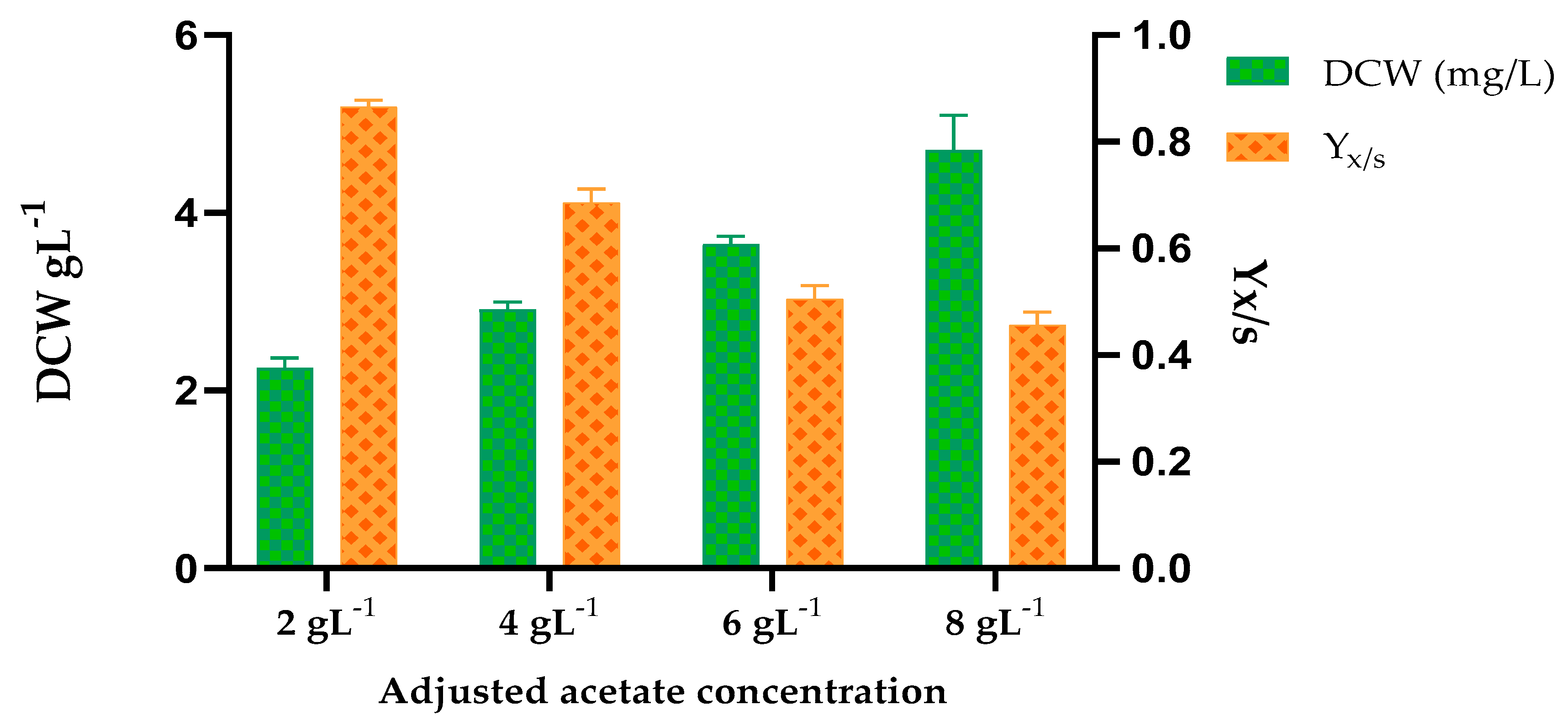
The yeast strain Y.lipolytica strain Po1f was used to demonstrate the feasibility of using acetate-rich fermentate derived from optimized anaerobic fermentation of SOWs. Y.lipolytica (American Type Culture Collection (ATCC) no. MYA 2613) was procured from American Type Culture Collection (ATCC), USA. It was maintained as a glycerol stock at −80 °C prior to growth study. The anaerobically fermented product of SOW was centrifuged at 10,000 rpm for 10 min to precipitate biomass and suspended/undigested solids. The acetate and other VFA concentrations were measured in the supernatant obtained after filtration through a 0.2 µM membrane. This acetate-rich VFA supernatant was used as a substrate for Y.lipolytica’s growth without additional nutrients. The final acetate concentrations of 2, 4, 6, and 8 g L−1 were adjusted by diluting with sterile de-ionized (DI) water. The pH was adjusted to 6.0 and the growth experiments were conducted in 500 mL Erlenmeyer flasks with a 200 mL working volume of fermentate and inoculum.
Y.lipolytica was precultured from glycerol stocks by inoculating 200 mL of YPD broth (20 g L−1 peptone, 10 g L−1 yeast extract, 20 g L−1 glucose) in Erlenmeyer flasks and incubated at 28 °C (200 rpm) until the late exponential growth phase (24 h–26 h). For growth study, this preculture was used to inoculate 200 mL VFA supernatant to a starting OD600 of 0.2. A control experiment with glucose (2 g L−1) as the sole carbon source was also set up. Growth of Y.lipolytica was assessed by measuring biomass density in terms of dry cell weight (DCW). DCW was determined by vacuum filtering 5 mL of culture in the stationary growth phase through a pre-dried and pre-weighed nitrocellulose filter, and the sample was washed several times. The samples were dried to a constant weight before analyzing DCW. All growth experiments were performed in triplicates. In the presence of high VFA concentration (>6 g L−1), the batch cultures were incubated for 20 days in order to accommodate for adaptation of the yeast (lag phase of growth) to unfavorable culture conditions. All other batch cultures having different concentrations of acetate were cultivated until a measurable biomass production was observed.
4. Conclusions
The present study was conducted to screen the effects of physical and chemical factors and to optimize acetate production by anaerobic fermentation of two SOWs—cafeteria waste and corn stover. The screening of influential factors using two-level factorial design revealed that incubation temperature, BES addition, and shaking conditions (100 rpm) had the most significant effect on acetate production among the 10 factors tested. The ANOVA analysis confirmed that the selected model was significant. The statistical optimization experiment using RSM resulted in a maximum acetate production of ~8000 mg L−1 at a temperature, pH, and C:N ratio of 60 °C, 6, and 25, respectively. It is noteworthy that the difference in the three levels selected by the central composite design for the C:N ratio had a considerable difference among them and fine-tuning of this parameter can further improve acetate production as is observed in previous studies. So, further optimization with a narrow range of C:N ratio can be done. Supplementation of methanogen inhibitor, BES, may not be needed if higher substrate loading is used as higher acetate and/or VFA production is known to inhibit methanogenesis. In the absence of acetoclastic methanogenesis, this can lead to further acetate accumulation.
This study also demonstrated the use of acetate-rich fermentate as a sole carbon source (without any nutrient addition) for the growth of an industrially relevant yeast selected for this study—Y.lipolytica—which can convert VFAs in the fermentation product (derived from cafeteria waste and corn stover) into higher-value-added products. According to the results of this study, different initial acetate concentrations exerted different inhibitory effects on cell growth which is mainly evident by the different duration of lag phase and biomass production (in terms of DCW). Higher initial acetate concentrations required longer lag phase though the biomass density was not affected significantly. Our future work is being directed toward engineering Y.lipolytica to improve acetate uptake and direct its carbon flux toward the heterologous expression of itaconic acid which has widespread industrial applications. Thus, developing an acetogenic consortium and optimizing VFA production for higher acetate concentrations was an important milestone in the direction of using low-cost waste substrate for the production of higher-value biochemicals using Y.lipolytica as a host organism. In addition, the acetate-rich fermentate can be used by other industrial microorganisms for the production of various by-products, the feasibility of which can be checked in future studies.








{kind=link}
{kind=link}
{kind=link}
{kind=link}