Plant Growth Promotion Abilities of Phylogenetically Diverse Mesorhizobium Strains: Effect in the Root Colonization and Development of Tomato Seedlings

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
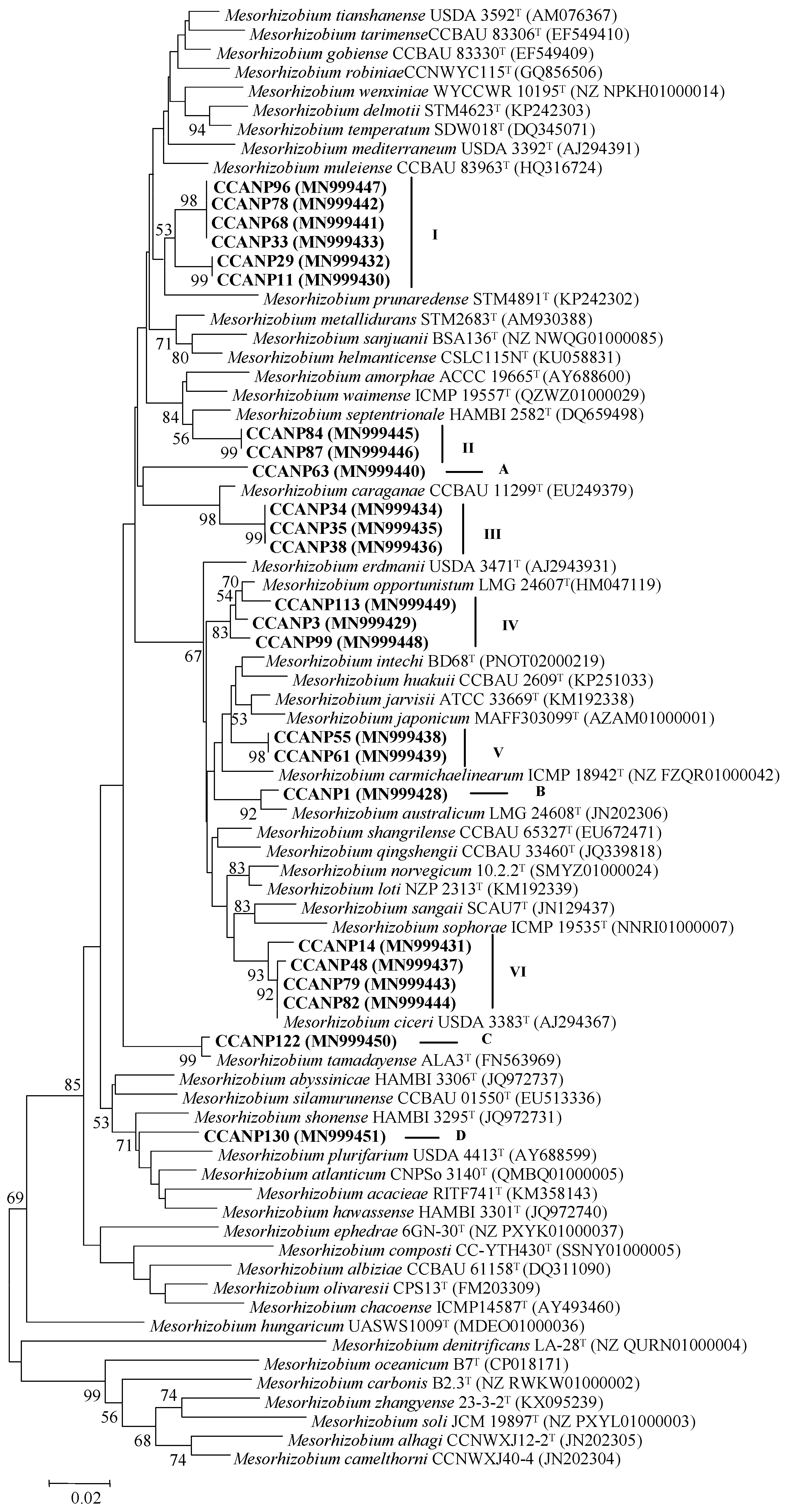
2.2. Phylogenetic Analysis
2.3. Analysis of In Vitro Plant Growth Promoting (PGP) Mechanisms
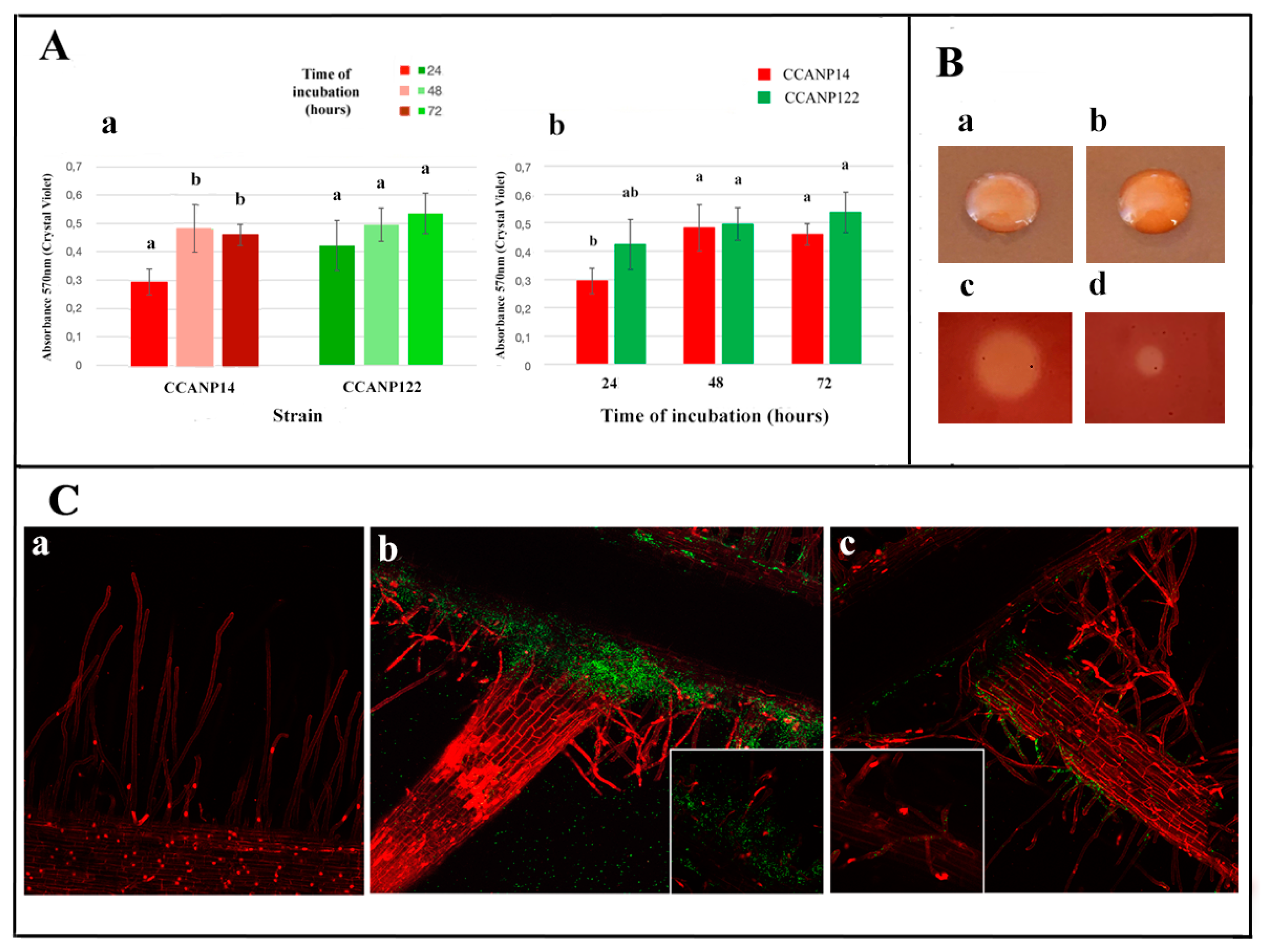
2.4. Tomato Root Colonization and Biofilm Production Assays
2.5. Microcosm Plant Assays
3. Results
3.1. Phylogenetic Analysis of the atpD Gene
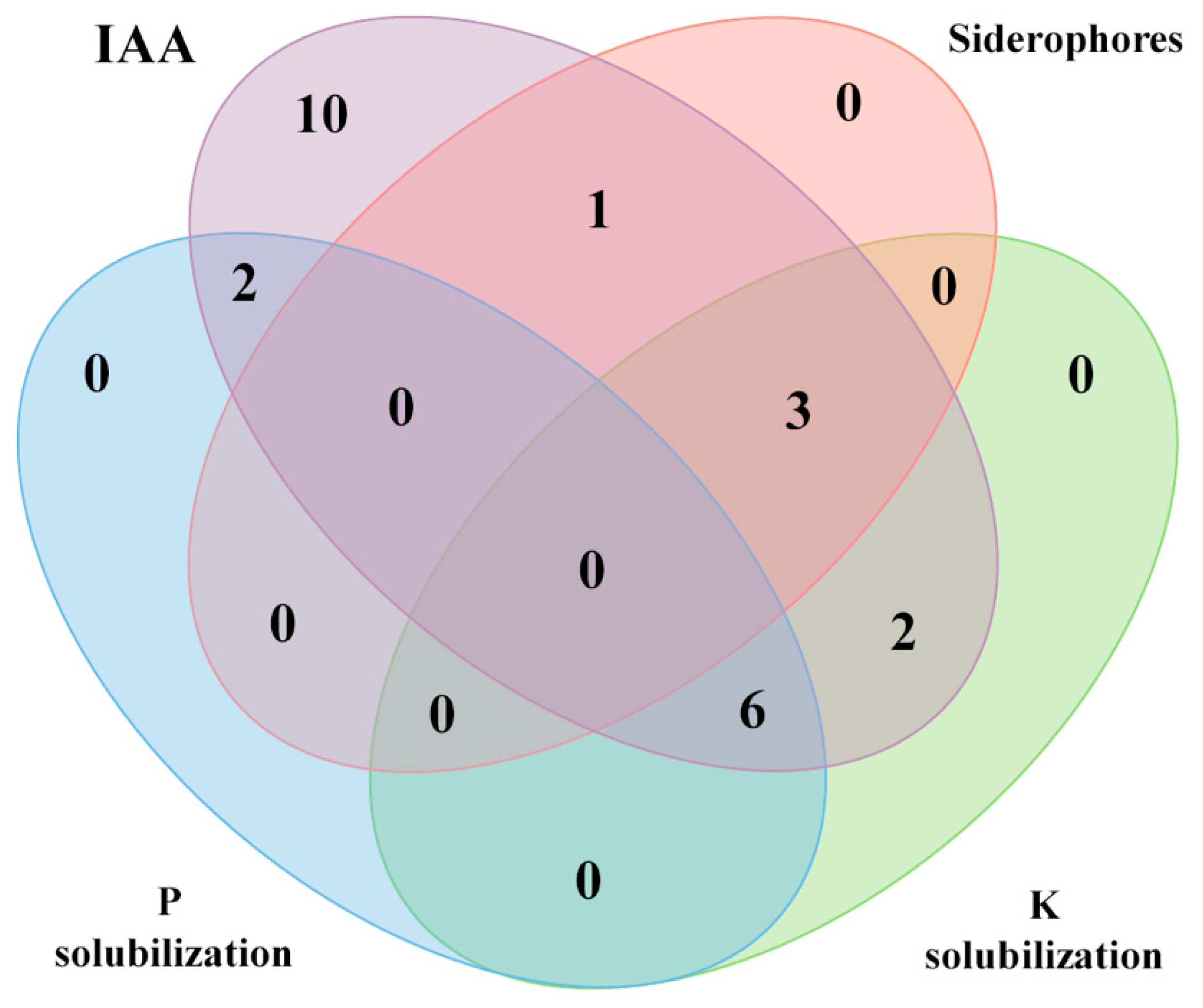
3.2. In Vitro PGP Mechanisms
3.3. In vitro Biofilm Formation and Tomato Root Colonization
3.4. Microcosm Plant Assays
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barquero, M.; Pastor-Bueis, R.; Urbano, B.; González-Andrés, F. Challenges, Regulations and Future Actions in Biofertilizers in the European Agriculture: From the Lab to the Field. In Microbial Probiotics for Agricultural Systems. Sustainability in Plant and Crop Protection; Zúñiga-Dávila, D., González-Andrés, F., Ormeño-Orrillo, E., Eds.; Springer: Cham, Switzerland, 2019; pp. 83–107. [Google Scholar]
- Hassan, M.K.; McInroy, J.A.; Jones, J.; Shantharaj, D.; Liles, M.R.; Kloepper, J.W. Pectin-rich amendment enhances soybean growth promotion and nodulation mediated by Bacillus velezensis strains. Plants 2019, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Ahmad, M.; Mumtaz, M.Z.; Ali, S.; Sarfraz, R.; Naveed, M.; Jamil, M.; Damalas, C.A. Integrated application of organic amendments with Alcaligenes sp. AZ9 improves nutrient uptake and yield of maize (Zea mays). J. Plant Growth Regul. 2020. [Google Scholar] [CrossRef]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.K.; McInroy, J.A.; Kloepper, J.W. The interactions of rhizodeposits with plant growth-promoting rhizobacteria in the rhizosphere: A review. Agriculture 2019, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, E.; Silva, L.R.; Ramírez-Bahena, M.H.; Peix, A. Diversity of potassium-solubilizing microorganisms and their interactions with plants. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V., Maurya, B., Verma, J., Meena, R., Eds.; Springer: New Delhi, India, 2018; pp. 99–110. [Google Scholar]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef] [Green Version]
- Patil, A.; Kale, A.; Ajane, G.; Sheikh, R.; Patil, S. Plant growth-promoting Rhizobium: Mechanisms and biotechnological prospective. In Rhizobium Biology and Biotechnology; Hansen, A., Choudhary, D., Agrawal, P., Varma, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 105–134. [Google Scholar]
- Vargas, L.K.; Volpiano, C.G.; Lisboa, B.B.; Giongo, A.; Beneduzi, A.; Passaglia, L.M.P. Potential of rhizobia as plant growth-promoting rhizobacteria. In Microbes for Legume Improvement; Zaidi, A., Khan, M., Musarrat, J., Eds.; Springer: Cham, Switzerland, 2017; pp. 153–174. [Google Scholar]
- Velázquez, E.; Carro, L.; Flores-Félix, J.D.; Menéndez, E.; Ramírez-Bahena, M.H.; Peix, A. Bacteria-inducing legume nodules involved in the improvement of plant growth, health and nutrition. In Microbiome in Plant Health and Disease; Kumar, V., Prasad, R., Kumar, M., Choudhary, D., Eds.; Springer: Singapore, 2019; pp. 79–104. [Google Scholar]
- Remigi, P.; Zhu, J.; Young, J.P.W.; Masson-Boivin, C. Symbiosis within symbiosis: Evolving nitrogen-fixing legume symbionts. Trends Microbiol. 2016, 24, 63–75. [Google Scholar] [CrossRef]
- García-Fraile, P.; Carro, L.; Robledo, M.; Ramírez-Bahena, M.H.; Flores-Félix, J.; Fernández, M.; Mateos, P.; Rivas, R.; Igual, J.; Martínez-Molina, E.; et al. Rhizobium promotes non-legumes growth and quality in several production steps: Towards a biofertilization of edible raw vegetables healthy for humans. PLoS ONE 2012, 7, e38122. [Google Scholar] [CrossRef] [Green Version]
- Flores-Félix, J.D.; Menéndez, E.; Rivera, L.P.; Marcos-García, M.; Martínez-Hidalgo, P.; Mateos, P.; Martínez-Molina, E.; Velázquez, E.; García-Fraile, P.; Rivas, R. Use of Rhizobium leguminosarum as a potential biofertilizer for Lactuca sativa and Daucus carota crops. J. Plant Nutr. Soil Sci. 2013, 176, 876–882. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; Flores-Félix, J.D.; García-Fraile, P.; Mateos, P.F.; Menéndez, E.; Velázquez, E.; Rivas, R. Probiotic activities of Rhizobium laguerreae on growth and quality of spinach. Sci. Rep. 2018, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Flores-Félix, J.D.; Silva, L.R.; Rivera, L.P.; Marcos-García, M.; García-Fraile, P.; Martínez-Molina, E.; Mateos, P.F.; Velázquez, E.; Andrade, P.; Rivas, R. Plants probiotics as a tool to produce highly functional fruits: The case of Phyllobacterium and vitamin C in strawberries. PLoS ONE 2015, 10, e0122281. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.D.; da Silva, P.R.D.; Stefenon, V.M. In vitro growth and indoleacetic acid production by Mesorhizobium loti SEMIA806 and SEMIA816 under the influence of copper ions. Microbiol. Res. 2017, 8, 57–58. [Google Scholar] [CrossRef]
- Brígido, C.; Glick, B.R.; Oliveira, S. Survey of plant growth-promoting mechanisms in native Portuguese chickpea Mesorhizobium isolates. Microb. Ecol. 2016, 73, 900–915. [Google Scholar] [CrossRef]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; Lai, W.A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Rivas, R.; Peix, A.; Mateos, P.F.; Trujillo, M.E.; Martínez-Molina, E.; Velázquez, E. Biodiversity of populations of phosphate solubilizing rhizobia that nodulates chickpea in different Spanish soils. Plant Soil 2006, 287, 23–33. [Google Scholar] [CrossRef]
- Imen, H.; Neila, A.; Adnane, B.; Manel, B.; Mabrouk, Y.; Saidi, M.; Bouaziz, S. Inoculation with phosphate solubilizing Mesorhizobium strains improves the performance of chickpea (Cicer aritenium L.) under phosphorus deficiency. J. Plant Nutr. 2015, 38, 1656–1671. [Google Scholar] [CrossRef]
- Pandey, R.P.; Srivastava, A.K.; Gupta, V.K.; O’Donovan, A.; Ramteke, P.W. Enhanced yield of diverse varieties of chickpea (Cicer arietinum L.) by different isolates of Mesorhizobium ciceri. Environ. Sustain. 2018, 1, 425–435. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, X.; Chen, W.; Huang, A. Isolation and identification of three potassium-solubilizing bacteria from rape rhizospheric soil and their effects on ryegrass. Geomicrobiol. J. 2017, 10, 873–880. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Flores-Félix, J.D.; Marcos-García, M.; Silva, L.R.; Menéndez, E.; Martínez-Molina, E.; Mateos, P.M.; Velázquez, E.; García-Fraile, P.; Andrade, P.; Rivas, R. Rhizobium as plant probiotic for strawberry production under microcosm conditions. Symbiosis 2015, 67, 25–32. [Google Scholar]
- Flores-Félix, J.D.; Menéndez, E.; Marcos-García, M.; Celador-Lera, L.; Rivas, R. Calcofluor white, an alternative to propidium iodide for plant tissues staining in studies of root colonization by fluorescent-tagged rhizobia. J. Adv. Biol. Biotechnol. 2015, 2, 65–70. [Google Scholar] [CrossRef]
- Hassen, A.I.; Labuschagne, N. Root colonization and growth enhancement in wheat and tomato by rhizobacteria isolated from the rhizoplane of grasses. World J. Microbiol. Biotechnol. 2010, 26, 1837–1846. [Google Scholar] [CrossRef]
- Bernabeu, P.R.; Pistorio, M.; Torres-Tejerizo, G.; Estrada-De los Santos, P.; Galar, M.L.; Boiardi, J.L.; Luna, M.F. Colonization and plant growth-promotion of tomato by Burkholderia tropica. Sci. Hortic. 2015, 191, 113–120. [Google Scholar] [CrossRef]
- Fujita, M.; Kusajima, M.; Okumura, Y.; Nakajima, M.; Minamisawa, K. Effects of colonization of a bacterial endophyte, Azospirillum sp. B510, on disease resistance in tomato. Biosci. Biotechnol. Biochem. 2017, 81, 1657–1662. [Google Scholar] [CrossRef] [Green Version]
- Al-Ali, A.; Deravel, J.; Krier, F.; Béchet, M.; Ongena, M.; Jacques, P. Biofilm formation is determinant in tomato rhizosphere colonization by Bacillus velezensis FZB42. Environm. Sci. Pollut. Res. 2018, 25, 29910–29920. [Google Scholar] [CrossRef]
- Robledo, M.; Rivera, L.; Jiménez-Zurdo, J.I.; Rivas, R.; Dazzo, F.; Velázquez, E.; Martínez-Molina, E.; Hirsch, A.M.; Mateos, P.F. Role of Rhizobium endoglucanase CelC2 in cellulose biosynthesis and biofilm formation on plant roots and abiotic surfaces. Microb. Cell Fact. 2012, 11, 125. [Google Scholar] [CrossRef] [Green Version]
- Jimenéz-Zurdo, J.I.; Mateos, P.F.; Dazzo, F.B.; Martínez-Molina, E. Cell-bound cellulase and polygalacturonase production by Rhizobium and Bradyrhizobium species. Soil Biol. Biochem. 1996, 28, 917–921. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; Ramírez-Bahena, M.H.; Flores-Félix, J.D.; Rivas, R.; Igual, J.M.; Mateos, P.F.; Martínez-Molina, E.; León-Barrios, M.; Peix, Á.; Velázquez, E. Revision of the taxonomic status of type strains of Mesorhizobium loti and reclassification of strain USDA 3471T as the type strain of Mesorhizobium erdmanii sp. nov. and ATCC 33669T as the type strain of Mesorhizobium jarvisii sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 1703–1708. [Google Scholar] [CrossRef]
- Pillay, V.K.; Nowak, J. Inoculum density, temperature, and genotype effects on in vitro growth promotion and epiphytic and endophytic colonization of tomato (Lycopersicon esculentum L.) seedlings inoculated with a pseudomonad bacterium. Can. J. Microbiol. 1997, 43, 354–361. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Saber, F.M.A.; Abdelhafez, A.A.; Hassan, E.A.; Ramadan, E.M. Characterization of fluorescent pseudomonads isolates and their efficiency on the growth promotion of tomato plant. Ann. Agric. Sci. 2015, 60, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Win, K.T.; Tanaka, F.; Okazaki, K.; Ohwaki, Y. The ACC deaminase expressing endophyte Pseudomonas spp. Enhances NaCl stress tolerance by reducing stress-related ethylene production, resulting in improved growth, photosynthetic performance, and ionic balance in tomato plants. Plant Physiol. Biochem. 2018, 127, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Abanda-Nkpwatt, D.; Müsch Jochen Tschiersch, M.; Schwab, M.B.W. Molecular interaction between Methylobacterium extorquens and seedlings: Growth promotion, methanol consumption, and localization of the methanol emission site. J. Exp. Bot. 2006, 57, 4025–4032. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Duraivadivel, P.; Hariprasad, P.; Niranjana, S.R. A novel split-pot bioassay to screen indole acetic acid producing rhizobacteria for the improvement of plant growth in tomato Solanum lycopersicum L. Sci. Hortic. 2017, 224, 351–357. [Google Scholar] [CrossRef]
- Hariprasad, P.; Niranjana, S.R. Isolation and characterization of phosphate solubilizing rhizobacteria to improve plant health of tomato. Plant Soil 2009, 316, 13–24. [Google Scholar] [CrossRef]
- Shen, M.; Kang, Y.J.; Wang, H.L.; Zhang, X.S.; Zhao, Q.X. Effect of Plant Growth-promoting Rhizobacteria (PGPRs) on plant growth, yield, and quality of tomato (Lycopersicon esculentum Mill.) under simulated seawater irrigation. J. Gen. Appl. Microbiol. 2012, 58, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; Walia, A.; Kulshrestha, S.; Chauhan, A.; Shirkot, C.K. Efficiency of plant growth-promoting P-solubilizing Bacillus circulans CB7 for enhancement of tomato growth under net house conditions. J. Basic Microbiol. 2015, 55, 33–44. [Google Scholar] [CrossRef]
- Abdallah, D.B.; Frikha-Gargouri, O.; Tounsi, S. Rhizospheric competence, plant growth promotion and biocontrol efficacy of Bacillus amyloliquefaciens subsp. plantarum strain 32a. Biol. Control 2018, 124, 61–67. [Google Scholar] [CrossRef]
- Cao, P.; Liu, C.; Sun, P.; Fu, X.; Wang, S.; Wu, F.; Wang, X. An endophytic Streptomyces sp. strain DHV3-2 from diseased root as a potential biocontrol agent against Verticillium dahliae and growth elicitor in tomato (Solanum lycopersicum). Antonie Van Leeuwenhoek 2016, 109, 1573–1582. [Google Scholar] [CrossRef]
- Romero, F.M.; Marina, M.; Pieckenstain, F.L. Novel components of leaf bacterial communities of field-grown tomato plants and their potential for plant growth promotion and biocontrol of tomato diseases. Res. Microbiol. 2016, 167, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.H.; Zhang, S.A.; Mo, X.D.; Li, Y.C.; Fu, Y.Q.; Liu, Z.G. Effects of plant growth-promoting rhizobacteria and N source on plant growth and N and P uptake by tomato grown on calcareous soils. Pedosphere 2017, 27, 1027–1036. [Google Scholar] [CrossRef]
- Cordero, I.; Balaguer, L.; Rincón, A.; Pueyo, J.J. Inoculation of tomato plants with selected PGPR represents a feasible alternative to chemical fertilization under salt stress. J. Plant Nutr. Soil Sci. 2018, 181, 694–703. [Google Scholar] [CrossRef]
- Syed-Ab-Rahman, S.F.; Xiao, Y.; Carvalhais, L.C.; Ferguson, B.J.; Schenk, P.M. Suppression of Phytophthora capsici infection and promotion of tomato growth by soil bacteria. Rhizosphere 2019, 9, 72–75. [Google Scholar] [CrossRef] [Green Version]
- Armas-Capote, N.; Pérez-Yépez, J.; Martínez-Hidalgo, P.; Garzón-Machado, V.; Del Arco-Aguilar, M.; Velázquez, E.; León-Barrios, M. Core and symbiotic genes reveal nine Mesorhizobium genospecies and three symbiotic lineages among rhizobia nodulating Cicer canariense in natural habitat (La Palma, Canary Is.). Syst. Appl. Microbiol. 2014, 37, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, M.W.; Turner, S.L.; Rigottier-Gois, L.; Lloyd-Macgilp, S.A.; Young, J.P. Phylogenies of atpD and recA support the small subunit rRNA-based classification of rhizobia. Int. J. Syst. Evol. Microbiol. 2001, 51, 2037–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The clustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. A neighbour-joining method: A new method for reconstructing phylogenetics trees. Mol. Biol. Evol. 1987, 44, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 3, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.M. A Manual for the Practical Study of the Root-Nodule Bacteria; Black Well Scientific Publications: Oxford, UK, 1970. [Google Scholar]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indol-acetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.; Seem, K.; Mishra, A.; Sopory, S. Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression of leaf and rhizosphere community in rice during salt stress. Plant Physiol. Biochem. 2013, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Yodov, K.S.; Kundu, B.S. Prospects of using phosphate solubilizing Pseudomonas as biofungicide. Indian J. Microbiol. 2004, 44, 91–94. [Google Scholar]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fert. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Hu, X.; Chen, J.; Guo, J. Two Phosphate- and Potassium-solubilizing Bacteria Isolated from Tianmu Mountain, Zhejiang, China. World J. Microbiol. Biotechnol. 2006, 22, 983. [Google Scholar] [CrossRef]
- Robledo, M.; Jiménez-Zurdo, J.I.; Soto, M.J.; Velázquez, E.; Dazzo, F.; Martínez-Molina, E.; Mateos, P.F. Development of functional symbiotic white clover root hairs and nodules requires tightly regulated production of rhizobial cellulase CelC2. Mol. Plant Microbe Interact. 2011, 24, 798–807. [Google Scholar] [CrossRef] [Green Version]
- Mateos, P.F.; Jimenez-Zurdo, J.I.; Chen, J.; Squartini, A.S.; Haack, S.K.; Martinez-Molina, E.; Hubbell, D.H.; Dazzo, F.B. Cell-associated pectinolytic and cellulolytic enzymes in Rhizobium leguminosarum biovar trifolii. Appl. Environ. Microbiol. 1992, 58, 1816–1822. [Google Scholar] [CrossRef] [Green Version]
- Rigaud, J.; Puppo, A. Indole-3-acetic acid catabolism by soybean bacteroids. J. Gen. Microbiol. 1975, 88, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Yépez, J.; Armas-Capote, N.; Velázquez, E.; Pérez-Galdona, R.; Rivas, R.; León-Barrios, M. Evaluation of seven housekeeping genes for multilocus sequence analysis of the genus Mesorhizobium: Resolving the taxonomic affiliation of the Cicer canariense rhizobia. Syst. Appl. Microbiol. 2014, 37, 553–559. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; Ramírez-Bahena, M.H.; Flores-Félix, J.D.; Igual, J.M.; Sanjuán, J.; León-Barrios, M.; Peix, A.; Velázquez, E. Reclassification of strains MAFF 303099T and R7A into Mesorhizobium japonicum sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 4936–4941. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Alexandre, A.; Oliveira, S. Legume growth-promoting rhizobia: An overview on the Mesorhizobium genus. Microbiol. Res. 2014, 169, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Peix, A.; Rivas-Boyero, A.; Mateos, P.; Rodriguez-Barrueco, C.; Martínez-Molina, E.; Velázquez, E. Growth promotion of chickpea and barley by a phosphate solubilizing strain of Mesorhizobium mediterraneum under growth chamber conditions. Soil Biol. Biochem. 2001, 33, 103–110. [Google Scholar] [CrossRef]
- Berraho, E.; Lesueur, D.; Diem, H.; Sasson, A. Iron requirement and siderophore production in Rhizobium ciceri during growth on an iron-deficient medium. World J. Microbiol. Biotechnol. 1997, 13, 501–510. [Google Scholar] [CrossRef]
- Poitout, A.; Martinière, A.; Kucharczyk, B.; Queruel, N.; Silva-Andia, J.; Mashkoor, S.; Gamet, L.; Varoquaux, F.; Paris, N.; Sentenac, H.; et al. Local signalling pathways regulate the Arabidopsis root developmental response to Mesorhizobium loti inoculation. J. Exp. Bot. 2017, 68, 1199–1211. [Google Scholar] [CrossRef]
- Jiang, J.; Li, J.; Dong, Y. Effect of calcium nutrition on resistance of tomato against bacterial wilt induced by Ralstonia solanacearum. Eur. J. Plant Pathol. 2013, 136, 547–555. [Google Scholar] [CrossRef]
- Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Panigrahi, R.; Mohapatra, P.K.; Ito, J.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity. Plant Sci. 2011, 180, 368–374. [Google Scholar] [CrossRef]
- Jones, J.B. Plant Nutrition. In Tomato Plant Culture: In the Field, Greenhouse, and Home Garden; Jones, J.B., Ed.; CRC Press: Boca Raton, FL, USA, 1999; pp. 51–54. [Google Scholar]
- Chandra, S.; Choure, K.; Dubey, R.C.; Maheshwari, D.K. Rhizosphere competent Mesorhizobium loti MP6 induces hair curling, inhibits Sclerotinia sclerotiorum and enhances growth of indian mustard (Brassica campestris). Braz. J. Microbiol. 2007, 38, 124–130. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strains | Closest Species | atpD Gene Similarity (%) | Cluster or Lineage | IAA (µg mL−1) | Phosphate Solubilization ¥ | Potassium Solubilization ¥ | Siderophore Production § |
|---|---|---|---|---|---|---|---|
| CCANP 1 | M. australicum | 98.6 | B | 8 | 0 | 1.50 | 1 |
| CCANP 3 | M. opportunistum | 99.2 | IV | 23 | 2.00 | 1.41 | 0 |
| CCANP 99 | M. opportunistum | 98.7 | IV | 23 | 0 | 0 | 0 |
| CCANP 113 | M. opportunistum | 98.9 | IV | 23 | 0 | 1.10 | 0 |
| CCANP 11 | M. muleiense | 96.8 | I | 33 | 0 | 1.10 | 0 |
| CCANP 29 | M. muleiense | 97.0 | I | 40 | 1.40 | 1.10 | 0 |
| CCANP 33 | M. muleiense | 96.8 | I | 35 | 0 | 0 | 0 |
| CCANP 68 | M. muleiense | 96.8 | I | 37 | 1.72 | 0 | 0 |
| CCANP 78 | M. muleiense | 96.8 | I | 24 | 0 | 0 | 0 |
| CCANP 96 | M. muleiense | 96.8 | I | 33 | 2.10 | ng | 0 |
| CCANP 14 | M. ciceri | 99.1 | VI | 68 | 2.40 | 1.39 | 0 |
| CCANP 48 | M. ciceri | 99.8 | VI | 35 | 2.25 | 1.82 | 0 |
| CCANP 79 | M. ciceri | 100 | VI | 49 | 2.06 | 1.57 | 0 |
| CCANP 82 | M. ciceri | 100 | VI | 42 | 1.58 | 1.49 | 0 |
| CCANP 34 | M. caraganae | 97.9 | III | 10 | 0 | 0 | 0 |
| CCANP 35 | M. caraganae | 98.0 | III | 8 | 0 | 0 | 0 |
| CCANP 38 | M. caraganae | 98.0 | III | 6 | 0 | 0 | 0 |
| CCANP 63 | M. robiniae | 95.4 | A | 5 | 0 | 0 | 0 |
| CCANP 55 | M. jarvisii | 96.6 | V | 40 | 0 | 0 | 0 |
| CCANP 61 | M. jarvisii | 96.6 | V | 36 | 0 | 0 | 1 |
| CCANP 84 | M. septentrionale | 97.7 | II | 31 | 0 | 0 | 0 |
| CCANP 87 | M. septentrionale | 97.7 | II | 35 | 0 | 0 | 0 |
| CCANP 122 | M. tamadayense | 99.4 | C | 69 | 0 | 1.22 | 1 |
| CCANP 130 | M. shonense | 96.4 | D | 53 | 0 | 1.44 | 1 |
| Treatments | SL (cm/Plant) | RL (cm/Plant) | SFW (g/Plant) | RFW (g/Plant) | SDW (g/Plant) | RDW (g/Plant) |
|---|---|---|---|---|---|---|
| Control | 12.71 (±1.17) a | 24.89 (±1.02) a | 4.18 (±0.53) a | 0.43 (±0.01) a | 0.27 (±0.02) a | 0.05 (±0.01) a |
| CCANP14 | 20.46 (±1.03) b | 34.33 (±1.64) c | 15.95 (±1.44) c | 3.15 (±0.38) c | 1.52 (±0.15) c | 0.38 (±0.05) c |
| CCANP122 | 23.25 (±1.80) b | 29.08 (±1.20) b | 14.34 (±1.11) b | 2.16 (±0.17) b | 1.15 (±0.09) b | 0.21 (±0.02) b |
| Treatments | N (g kg−1) | P (g kg−1) | Ca (g kg−1) | K (g kg−1) | Mg (g kg−1) | Na (g kg−1) | Fe (mg kg−1) | Mn (mg kg−1) | Cu (mg kg−1) | Zn (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 4.4 (±0.2) a | 3.1 (±0.2) a | 12.6 (±0.8) a | 54.5 (±2.4) a | 4.9 (±0.4) a | 1.4 (±0.1) a | 157.3 (±8.6) ab | 45.8 (±2.5) a | 7.8 (±0.5) a | 66.0 (±2.8) a |
| CCANP14 | 3.9 (±0.1) a | 3.1 (±0.2) a | 18.4 (±1.8) b | 58.1 (±1.6) a | 5.4 (±0.4) a | 1.3 (±0.1) a | 195.8 (±3.6) a | 38.0 (±2.5) ab | 9.5 (±1.2) a | 47.3 (±2.3) b |
| CCANP122 | 4.2 (±0.1) a | 3.5 (±0.1) a | 11.6 (±0.4) a | 66.4 (±1.2) b | 3.8 (±0.1) a | 2.0 (±0.1) b | 107.5 (±5.4) b | 36.0 (±2.3) b | 8.8 (±0.3) a | 44.8 (±4.7) b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menéndez, E.; Pérez-Yépez, J.; Hernández, M.; Rodríguez-Pérez, A.; Velázquez, E.; León-Barrios, M. Plant Growth Promotion Abilities of Phylogenetically Diverse Mesorhizobium Strains: Effect in the Root Colonization and Development of Tomato Seedlings. Microorganisms 2020, 8, 412. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030412
Menéndez E, Pérez-Yépez J, Hernández M, Rodríguez-Pérez A, Velázquez E, León-Barrios M. Plant Growth Promotion Abilities of Phylogenetically Diverse Mesorhizobium Strains: Effect in the Root Colonization and Development of Tomato Seedlings. Microorganisms. 2020; 8(3):412. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030412
Chicago/Turabian StyleMenéndez, Esther, Juan Pérez-Yépez, Mercedes Hernández, Ana Rodríguez-Pérez, Encarna Velázquez, and Milagros León-Barrios. 2020. "Plant Growth Promotion Abilities of Phylogenetically Diverse Mesorhizobium Strains: Effect in the Root Colonization and Development of Tomato Seedlings" Microorganisms 8, no. 3: 412. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030412