Pinocembrin-7-Glucoside (P7G) Reduced Postharvest Blue Mold of Navel Orange by Suppressing Penicillium italicum Growth



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Material
2.2. Phytopathogenic Fungi Isolation
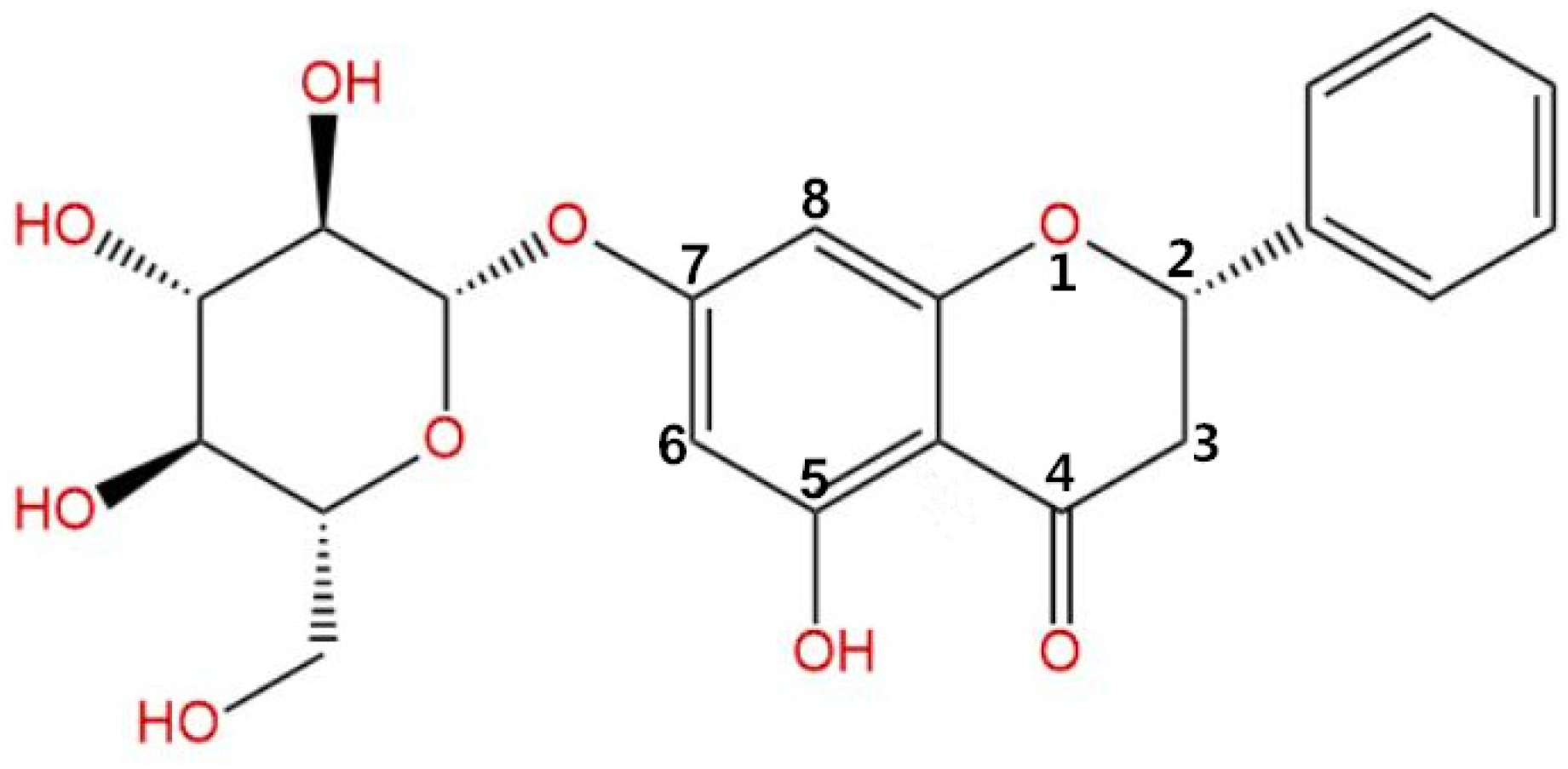
2.3. P7G Isolation and Purification
2.4. In Vitro Antifungal Activity Assay
2.4.1. Minimal Inhibitory Concentration (MIC) and Minimum Fungicidal Concentration (MFC)
2.4.2. Assay of P7G on Mycelial Growth of P. italicum
2.4.3. Assay of P7G on Spore Germination of P. italicum
2.5. In Vivo Antifungal Activity Assay on Navel Orange Fruit
2.6. Microscopic Observations
2.7. Determination of Cell Membrane Permeability
2.8. Measurement of Intracellular Constituent Contents
2.8.1. Mycelial Growth Weight
2.8.2. Evaluation of Soluble Protein and Reducing Sugar Levels
2.8.3. Total Lipid Content
2.9. Assays of Cell Wall Components Content and Enzyme Activities
2.10. Statistical Analysis
3. Results
3.1. P7G Displays Potential Used as Antifungal Agent in Citrus Phytopathogenic Fungi
3.2. P7G has Antifungal Activity Against P. italicum
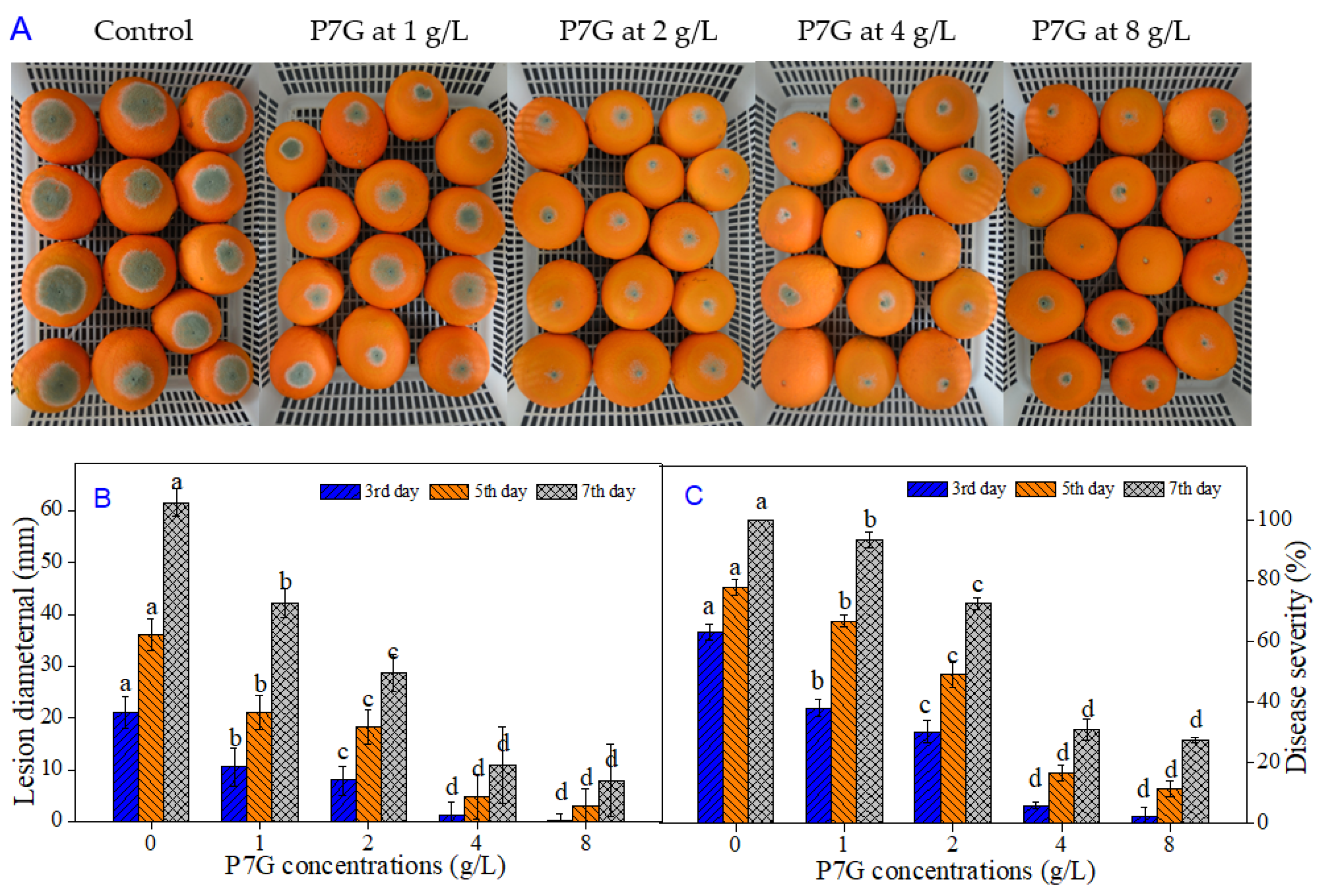
3.3. P7G Inhibits Blue Mold Development on Navel Orange Infected with P. italicum
3.4. P7G Alters the Morphology and Ultrastructure of P. italicum
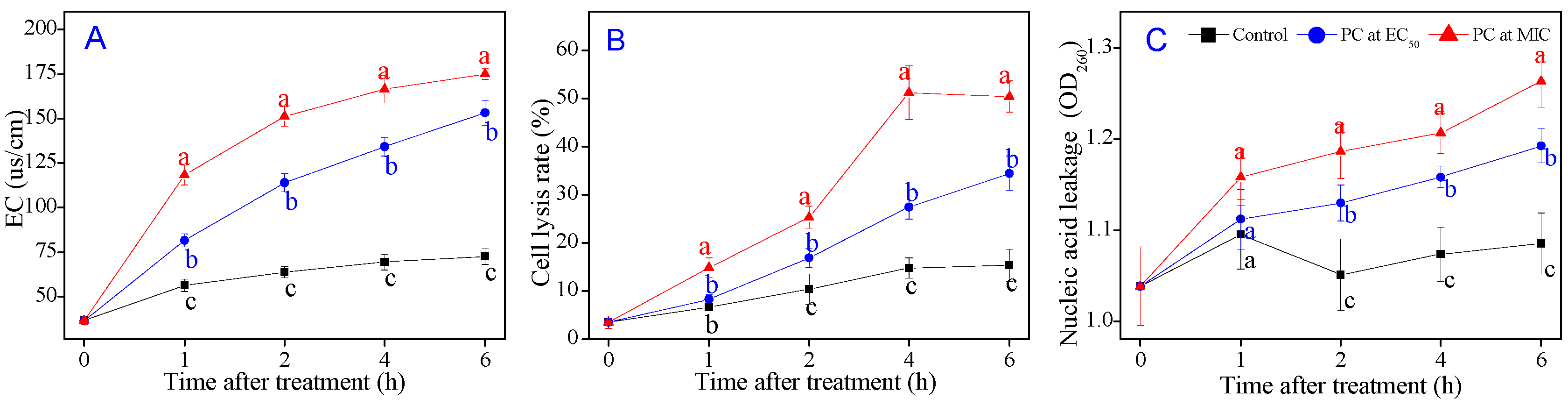
3.5. P7G Causes Cell Membrane Damage of P. italicum
3.6. P7G Induces the Outflow of Intercellular Inclusions of P. italicum
3.7. P7G Modifies the Cell Wall Structure of P. italicum
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jiang, C.C.; Zhang, Y.F.; Lin, Y.J.; Chen, Y.; Lu, X.K. Illuminafi Sequencing Reveals Candidate Genes of Carotenoid Metabolism in Three Pummelo Cultivars (Citrus Maxima) with Different Pulp Color. Int. J. Mol. Sci. 2019, 20, 2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Otmani, M.; Ait-Oubahou, A.; Zacarías, L. Citrus Spp.: Orange, Mandarin, Tangerine, Clementine, Grapefruit, Pomelo, Lemon and Lime. In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Yahia, E.M., Ed.; Woodhead Publishing: Cambridge, UK, 2011; pp. 437–516. [Google Scholar]
- Chen, C.; Qi, W.; Peng, X.; Chen, J.; Wan, C. Inhibitory effect of 7-demethoxytylophorine on Penicillium italicum and its possible mechanism. Microorganisms 2019, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Qian, X.; Dhanasekaran, S.; Boateng, N.A.S.; Yan, X.; Zhu, H.; He, F.; Zhang, H. Study on the infection mechanism of Penicillium digitatum on postharvest citrus (Citrus reticulata Blanco) based on transcriptomics. Microorganisms 2019, 7, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, S.; Ouyang, Q.; You, K.; Qian, J.; Tao, N. An inclusion complex of thymol into β-cyclodextrin and its antifungal activity against Geotrichum citri-aurantii. Postharvest Biol. Tech. 2018, 138, 31–36. [Google Scholar] [CrossRef]
- Xu, L.; Tao, N.; Yang, W.; Jing, G. Cinnamaldehyde damaged the cell membrane of Alternaria alternata and induced the degradation of mycotoxins in vivo. Ind. Crop. Prod. 2018, 112, 427–433. [Google Scholar] [CrossRef]
- Galsurker, O.; Diskin, S.; Maurer, D.; Feygenberg, O.; Alkan, N. Fruit stem-end rot. Horticulturae 2018, 4, 50. [Google Scholar] [CrossRef] [Green Version]
- Erasmus, A.; Lennox, C.L.; Korsten, L.; Lesar, K.; Fourie, P.H. Imazalil resistance in Penicillium digitatum and P. italicum causing citrus postharvest green and blue mould: Impact and options. Postharvest Biol. Tech. 2015, 107, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Hao, W.; Zhong, G.; Hu, M.; Luo, J.; Weng, Q.; Rizwan-ul-Haq, M. Control of citrus postharvest green and blue mold and sour rot by tea saponin combined with imazalil and prochloraz. Postharvest Biol. Tech. 2010, 56, 39–43. [Google Scholar] [CrossRef]
- Altieri, G.; Di Renzo, G.C.; Genovese, F.; Calandra, M.; Strano, M.C. A new method for the postharvest application of imazalil fungicide to citrus fruit. Biosyst. Eng. 2013, 115, 434–443. [Google Scholar] [CrossRef]
- Tripathi, P.; Dubey, N.K. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables. Postharvest Biol. Tech. 2004, 32, 235–245. [Google Scholar] [CrossRef]
- Talibi, I.; Boubaker, H.; Boudyach, E.; Ait Ben Aoumar, A. Alternative methods for the control of postharvest citrus diseases. J. Appl. Microbiol. 2014, 117, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shen, Y.; Chen, C.; Wan, C. Inhibition of key citrus postharvest fungal strains by plant extracts in vitro and in vivo: A review. Plants 2019, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, T.; Chen, Q.L.; He, X.C.; So, S.W.; Lo, Y.L.; Fan, L.L.; Xu, J.; Tang, Y.; Zhang, J.Y.; Zhao, Z.Z.; et al. Chemical quantification and antioxidant assay of four active components in Ficus hirta root using UPLC-PAD-MS fingerprinting combined with cluster analysis. Chem. Cent. J. 2013, 7, 1752–1760. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Yi, X.; Chen, H.; Wang, Y.; He, X. Anti-inflammatory phenylpropanoids and phenolics from Ficus hirta Vahl. Fitoterapia 2017, 121, 229–234. [Google Scholar] [CrossRef]
- Ya, J.; Zhang, X.Q.; Wang, Y.; Zhang, Q.W.; Chen, J.X.; Ye, W.C. Two new phenolic compounds from the roots of Ficus hirta. Nat. Prod. Res. 2010, 24, 621–625. [Google Scholar] [CrossRef]
- Zeng, Y.W.; Liu, X.Z.; Lv, Z.C.; Peng, Y.H. Effects of Ficus hirta Vahl. (Wuzhimaotao) extracts on growth inhibition of Hela cells. Exp. Toxicol. Pathol. 2012, 64, 743–749. [Google Scholar] [CrossRef]
- Chen, C.; Wan, C.; Peng, X.; Chen, Y.; Chen, M.; Chen, J. Optimization of antifungal extracts from Ficus hirta fruits using response surface methodology and antifungal activity tests. Molecules 2015, 20, 19647–19659. [Google Scholar] [CrossRef]
- Chen, C.; Peng, X.; Zeng, R.; Chen, M.; Wan, C.; Chen, J. Ficus hirta fruits extract incorporated into an alginate-based edible coating for nanfeng mandarin preservation. Sci. Hortic. 2016, 202, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Cai, N.; Chen, J.; Peng, X.; Wan, C. Chitosan-based coating enriched with hairy fig (Ficus hirta Vahl.) fruit extract for “Newhall” navel orange preservation. Coatings 2018, 8, 445. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Nie, Z.; Wan, C.; Chen, J. Preservation of xinyu tangerines with an edible coating using Ficus hirta Vahl. fruits extract-incorporated chitosan. Biomolecules 2019, 9, 46. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.B.; Zhai, Y.D.; Ma, Z.P.; Yang, C.J.; Pan, R.; Yu, J.L.; Wang, Q.H.; Yang, B.Y.; Kuang, H.X. Triterpenoids and flavonoids from the leaves of astragalus membranaceus and their inhibitory effects on nitric oxide production. Chem. Biodivers. 2015, 12, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Chen, C.; Li, M.; Yang, Y.; Chen, M.; Chen, J. Chemical constituents and antifungal activity of Ficus hirta Vahl. fruits. Plants 2017, 6, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Zhao, B.; Wang, Y.; Lin, N.; Wang, L.; Song, T.; Yang, H. A new flavonoid glycoside and potential antifungal activity of isolated constituents from the flowers of Lilium lancifolium. J. Chem. Res. 2015, 39, 260–262. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, Y.; Ye, J.; Fan, G.; Peng, L.; Pan, S. Effects of poplar buds as an alternative to propolis on postharvest diseases control of strawberry fruits. J. Sci. Food Agric. 2016, 96, 2136–2141. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, X.; Chen, X.; Ba, Y.; Zhang, N.; Xu, R.; Zhao, W.; Wu, X. Flavonones from Penthorum chinense ameliorate hepatic steatosis by activating the sirt1/ampk pathway in HepG2 cells. Int. J. Mol. Sci. 2018, 19, 2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.W.; Jiang, Y.; Zhang, D.Y.; Wang, M.; Chen, W.S.; Su, H.X.; Wang, Y.T.; Wan, J.B. Protective effects of Penthorum chinense Pursh against chronic ethanol-induced liver injury in mice. J. Ethnopharmacol. 2015, 161, 92–98. [Google Scholar] [CrossRef]
- Chen, C.; Wan, C.; Peng, X.; Chen, J. A flavonone pinocembroside inhibits Penicillium italicum growth and blue mold development in ‘Newhall’ navel oranges by targeting membrane damage mechanism. Pestic. Biochem. Phys. 2020. [Google Scholar] [CrossRef]
- Chen, C.; Cai, N.; Chen, J.; Wan, C. UHPLC-Q-TOF/MS-based metabolomics approach reveals the antifungal potential of pinocembroside against citrus green mold phytopathogen. Plants 2020, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Cai, N.; Chen, J.; Wan, C. Clove essential oil as an alternative approach to control postharvest blue mold caused by Penicillium italicum in citrus fruit. Biomolecules 2019, 9, 197. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Peng, X.; Chen, J.; Wan, C. Antioxidant, antifungal activities of ethnobotanical Ficus hirta Vahl. and analysis of main constituents by HPLC-MS. Biomedicines 2020, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Tao, N.; OuYang, Q.; Jia, L. Citral inhibits mycelial growth of Penicillium italicum by a membrane damage mechanism. Food Control 2014, 41, 116–121. [Google Scholar] [CrossRef]
- Huang, F.; Kong, J.; Ju, J.; Zhang, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Xie, Y.; Yao, W. Membrane damage mechanism contributes to inhibition of trans-cinnamaldehyde on Penicillium italicum using Surface- Enhanced Raman Spectroscopy (SERS). Sci. Rep. 2019, 9, 490. [Google Scholar] [CrossRef] [PubMed]
- Stalhberger, T.; Simenel, C.; Clavaud, C.; Eijsink, V.G.; Jourdain, R.; Delepierre, M.; Latgé, J.P.; Breton, L.; Fontaine, T. Chemical organization of the cell wall polysaccharide core of Malassezia restricta. J. Biol. Chem. 2014, 289, 12647–12656. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zheng, J.; Wan, C.; Chen, M.; Chen, J. Effect of carboxymethyl cellulose coating enriched with clove oil on postharvest quality of ’Xinyu’ mandarin oranges. Fruits 2016, 71, 319–327. [Google Scholar] [CrossRef]
- Peng, L.; Yang, S.; Cheng, Y.J.; Chen, F.; Pan, S.; Fan, G. Antifungal activity and action mode of pinocembrin from propolis against Penicillium italicum. Food Sci. Biotech. 2012, 21, 1533–1539. [Google Scholar] [CrossRef]
- Tayel, A.A.; Moussa, S.H.; Salem, M.F.; Mazrou, K.E.; El-Tras, W.F. Control of citrus molds using bioactive coatings incorporated with fungal chitosan/plant extracts composite. J. Sci. Food Agric. 2015, 96, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.X.; Zhao, H.F.; Lyu, L.F.; Huang, Z.J.; Fan, S.F.; Wu, W.L.; Li, W.L. Synergistic effect of natural antifungal agents for postharvest diseases of blackberry fruits. J. Sci. Food Agric. 2019, 99, 3343–3349. [Google Scholar] [CrossRef]
- Ouyang, Q.L.; Jia, L.; Tao, N.G.; He, X.L. Inhibitory effect of α-terpineol on Penicillium italicum. J. Food Sci. 2014, 35, 32–35. [Google Scholar] [CrossRef]
- D’Aquino, S.; Schirra, M.; Palma, A.; Angioni, A.; Cabras, P.; Migheli, Q. Residue levels and effectiveness of pyrimethanil vs imazalil when using heated postharvest dip treatments for control of Penicillium decay on citrus fruit. J. Agric. Food Chem. 2006, 54, 4721–4726. [Google Scholar] [CrossRef]
- Pérez-Alfonso, C.O.; Martínez-Romero, D.; Zapata, P.J.; Serrano, M.; Valero, D.; Castillo, S. The effects of essential oils carvacrol and thymol on growth of Penicillium digitatum and P. italicum involved in lemon decay. Int. J. Food Microbiol. 2012, 158, 101–106. [Google Scholar] [CrossRef]
- Li Destri Nicosia, M.G.; Pangallo, S.; Raphael, G.; Romeo, F.V.; Strano, M.C.; Rapisarda, P.; Droby, S.; Schena, L. Control of postharvest fungal rots on citrus fruit and sweet cherries using a pomegranate peel extract. Postharvest Biol. Tech. 2016, 114, 54–61. [Google Scholar] [CrossRef]
- Duan, X.; Jing, G.; Fan, F.; Tao, N. Control of postharvest green and blue molds of citrus fruit by application of sodium dehydroacetate. Postharvest Biol. Tech. 2016, 113, 17–19. [Google Scholar] [CrossRef]
- Wang, F.; Deng, J.; Jiao, J.; Lu, Y.; Yang, L.; Shi, Z. The combined effects of carboxymethyl chitosan and Cryptococcus laurentii treatment on postharvest blue mold caused by Penicillium italicum in grapefruit fruit. Sci. Hortic. 2019, 253, 35–41. [Google Scholar] [CrossRef]
- Palou, L.; Ali, A.; Fallik, E.; Romanazzi, G. GRAS, plant- and animal-derived compounds as alternatives to conventional fungicides for the control of postharvest diseases of fresh horticultural produce. Postharvest Biol. Tech. 2016, 122, 41–52. [Google Scholar] [CrossRef]
- Mari, M.; Bautista-Baños, S.; Sivakumar, D. Decay control in the postharvest system: Role of microbial and plant volatile organic compounds. Postharvest Biol. Tech. 2016, 122, 70–81. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, J.; Li, D.; Shang, C.; Peng, L.; Pan, S. The structure-antifungal activity relationship of 5,7-dihydroxyflavonoids against Penicillium italicum. Food Chem. 2017, 224, 26–31. [Google Scholar] [CrossRef]
- Chen, Y.H.; Lu, M.H.; Guo, D.S.; Zhai, Y.Y.; Miao, D.; Yue, J.Y.; Yuan, C.H.; Zhao, M.M.; An, D.R. Antifungal effect of magnolol and honokiol from Magnolia officinalis on Alternaria alternata causing tobacco brown spot. Molecules 2019, 24, 2140. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, H.; Robles-Kelly, C.; Rubio, J.; Olea, A.F.; Martínez, R.; Silva-Moreno, E. Antifungal effect of polygodial on Botrytis cinerea, a fungal pathogen affecting table grapes. Int. J. Mol. Sci. 2017, 18, 2251. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Jin, L.; Cai, Y.; Zheng, X.; Yu, T. (1→3)-β-d-glucan from yeast cell wall: Characteristic and potential application in controlling postharvest disease of pear. Postharvest Biol. Tech. 2019, 154, 105–114. [Google Scholar] [CrossRef]
- Bartnicki-Garcia, S. Cell wall chemistry, morphogenesis, and taxonomy of fungi. Ann. Rev. Microbiol. 1968, 22, 87–108. [Google Scholar] [CrossRef]
- OuYang, Q.; Duan, X.; Li, L.; Tao, N. Cinnamaldehyde exerts its antifungal activity by disrupting the cell wall integrity of Geotrichum citri-aurantii. Front. Microbiol. 2019, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Y.; Wu, X.M.; Yin, X.H.; Liang, J.N.; Li, M. The natural product citral can cause significant damage to the hyphal cell walls of Magnaporthe grisea. Molecules 2014, 19, 10279–10290. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of P7G (g/L) | Pathogenic Fungus | ||||||
|---|---|---|---|---|---|---|---|
| Alternaria citri | Colletotrichum gloeosporioides | Diaporthe citri | Geotrichum citri-aurantii | Penicillium digitatum | Penicillium italicum | ||
| Mycelial growth for 2nd d | 0 | + | + | + | + | + | + |
| 0.025 | + | + | + | + | + | + | |
| 0.05 | + | + | + | + | + | + | |
| 0.1 | + | + | − | + | + | + | |
| 0.2 | + | − | − | − | − | − | |
| 0.4 | + | − | − | − | − | − | |
| 0.8 | − | − | − | − | − | − | |
| Mycelial growth for 6th d | 0 | + | + | + | + | + | + |
| 0.025 | + | + | + | + | + | + | |
| 0.05 | + | + | + | + | + | + | |
| 0.1 | + | + | + | + | + | + | |
| 0.2 | + | + | + | + | + | + | |
| 0.4 | + | − | − | + | − | + | |
| 0.8 | − | − | − | − | − | − | |
| MIC | 0.8 | 0.2 | 0.1 | 0.2 | 0.2 | 0.2 | |
| MFC | 0.8 | 0.4 | 0.4 | 0.8 | 0.4 | 0.8 | |
| P7G Concentration (mg/mL) | Spore Germination (%) | Inhibitory Rate (%) | EC50 (mg/mL) |
|---|---|---|---|
| 0 | 93.40 ± 1.90 a | 0.00 ± 0.00 g | 0.032 |
| 0.025 | 67.17 ± 2.05 b | 28.09 ± 2.20 f | |
| 0.05 | 46.50 ± 1.60 c | 50.21 ± 1.71 e | |
| 0.1 | 24.60 ± 1.20 d | 73.66 ± 1.28 d | |
| 0.2 | 13.92 ± 0.87 e | 85.10 ± 0.94 c | |
| 0.4 | 3.50 ± 0.20 f | 96.25 ± 0.21 b | |
| 0.8 | 0.00 ± 0.00 g | 100.00 ± 0.00 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Chen, J.; Wan, C. Pinocembrin-7-Glucoside (P7G) Reduced Postharvest Blue Mold of Navel Orange by Suppressing Penicillium italicum Growth. Microorganisms 2020, 8, 536. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040536
Chen C, Chen J, Wan C. Pinocembrin-7-Glucoside (P7G) Reduced Postharvest Blue Mold of Navel Orange by Suppressing Penicillium italicum Growth. Microorganisms. 2020; 8(4):536. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040536
Chicago/Turabian StyleChen, Chuying, Jinyin Chen, and Chunpeng Wan. 2020. "Pinocembrin-7-Glucoside (P7G) Reduced Postharvest Blue Mold of Navel Orange by Suppressing Penicillium italicum Growth" Microorganisms 8, no. 4: 536. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040536