Exploring Coagulase-Negative Staphylococci Diversity from Artisanal Llama Sausages: Assessment of Technological and Safety Traits


 and
and 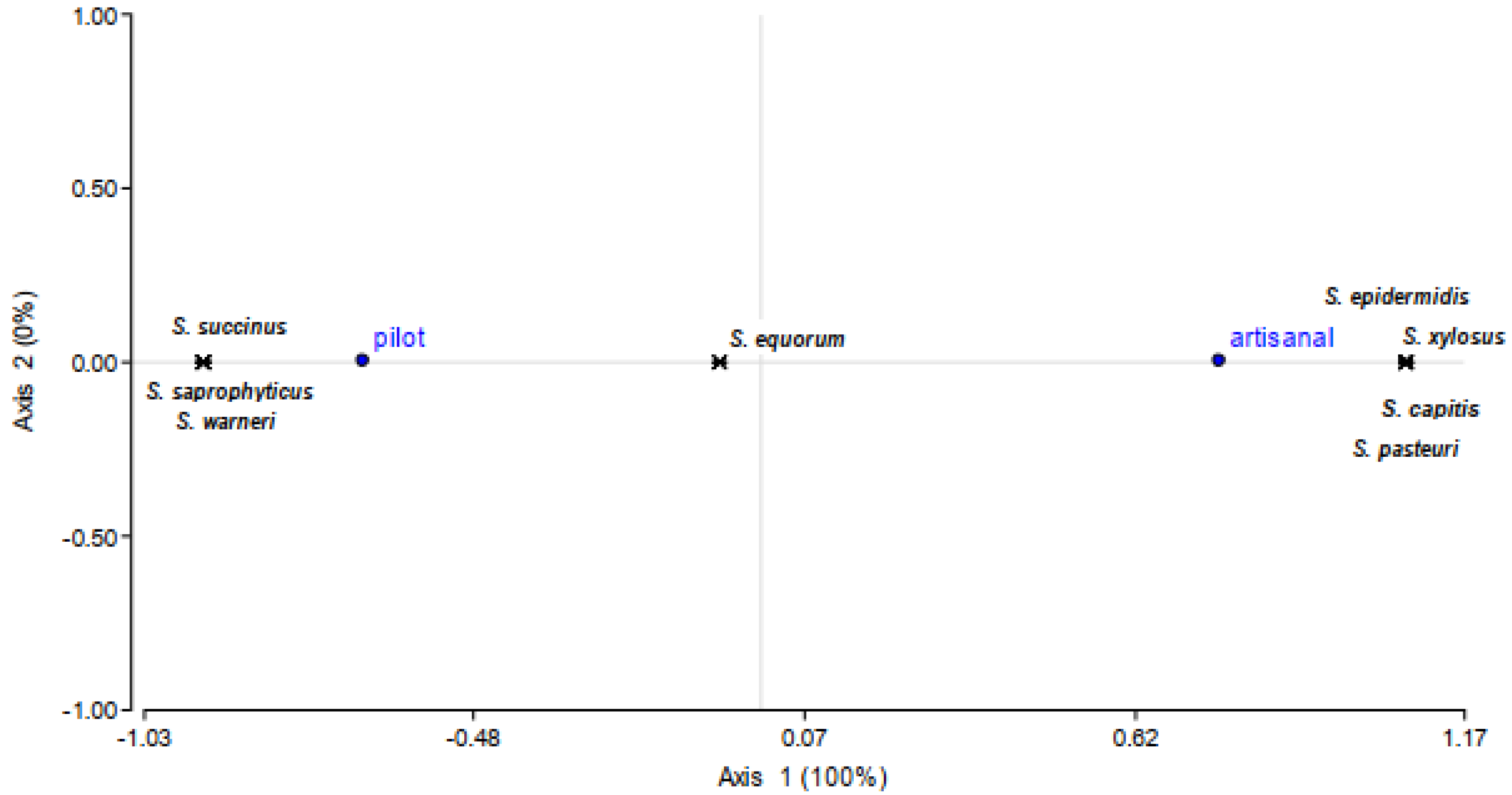{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sausage Production and Sampling
2.2. Molecular Identification and Typing
2.3. Technological Characterization
2.3.1. Proteolytic and Lipolytic Activity
2.3.2. Nitrate Reductase Activity
2.4. Phenotypic Traits and Safety Characterization
2.4.1. Production of Biogenic Amines (BA)
2.4.2. Hemolytic Activity
2.4.3. Biofilm Production Capacity
2.4.4. Antibiotic Resistance (AR)
2.5. Molecular Characterization of Antibiotic Resistance and Virulence Determinants
3. Results and Discussion
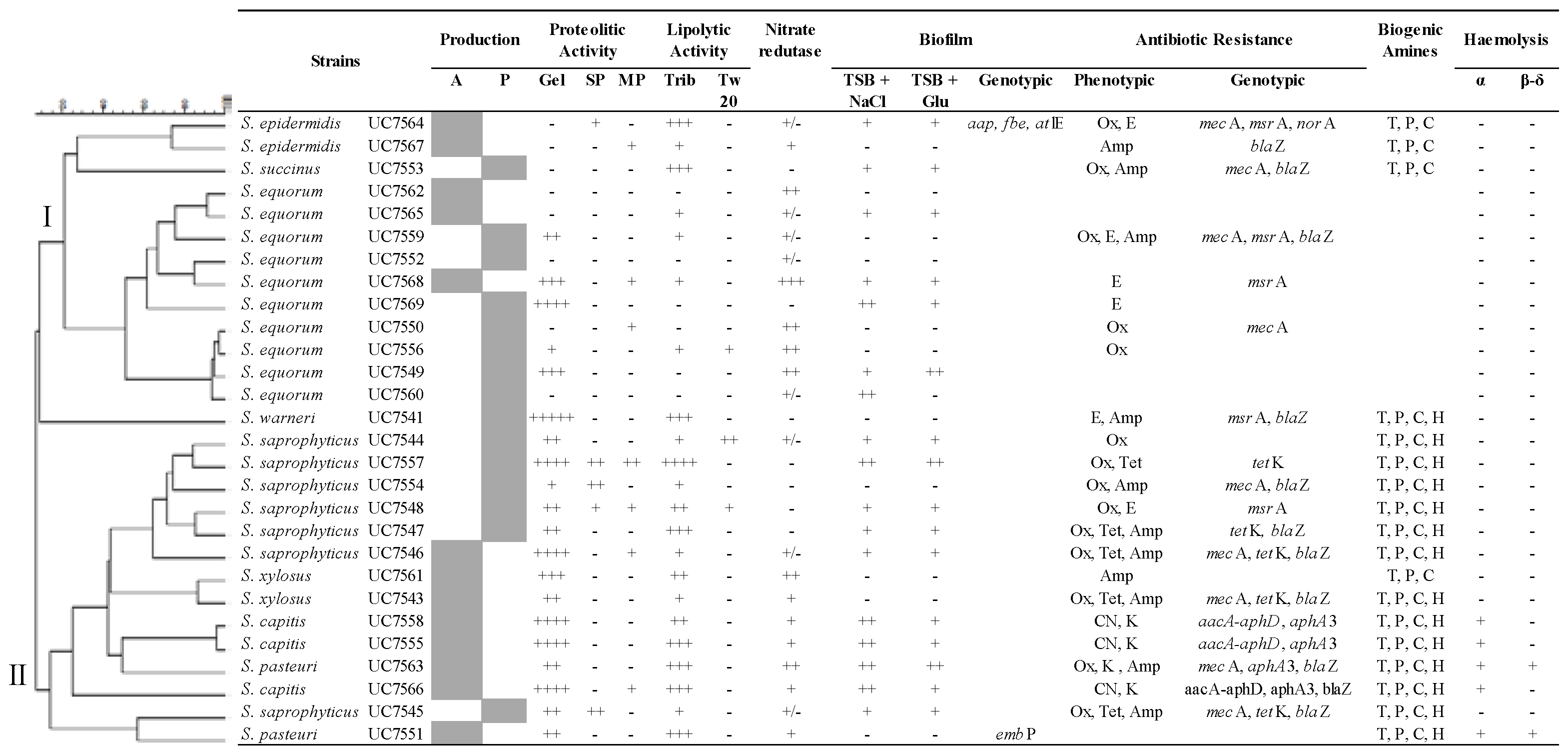
3.1. Typing and Identification of Staphylococci in Llama Sausages
3.2. Technological Properties
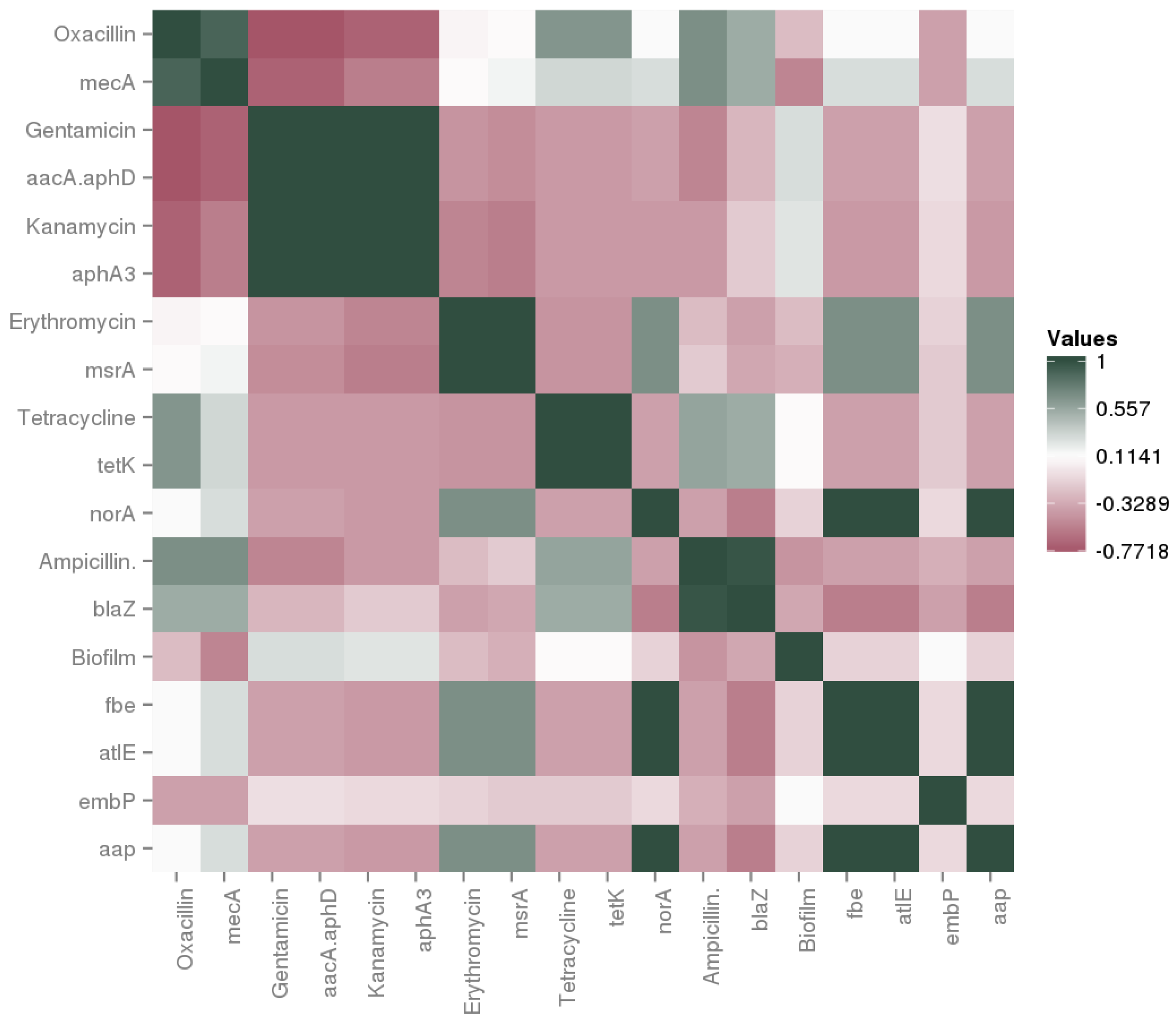
3.3. Safety Aspect
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pérez, P.; Maino, M.; Guzmán, R.; Vaquero, A.; Köbrich, C.; Pokniak, J. Carcass characteristics of llamas (Lama glama) reared in Central Chile. Small Rumin. Res. 2000, 37, 93–97. [Google Scholar] [CrossRef]
- Mendoza, L.M.; Padilla, B.; Belloch, C.; Vignolo, G. Diversity and enzymatic profile of yeasts isolated from traditional llama meat sausages from north-western Andean region of Argentina. Food Res. Int. 2014, 62, 572–579. [Google Scholar] [CrossRef]
- Cristofanelli, S.; Antonini, M.; Torres, D.; Polidori, P.; Renieri, C. Meat and carcass quality from Peruvian llama (Lama glama) and alpaca (Lama pacos). Meat Sci. 2004, 66, 589–593. [Google Scholar] [CrossRef]
- Aquilanti, L.; Garofalo, C.; Osimani, A.; Clementi, F. Ecology of lactic acid bacteria and coagulase negative cocci in fermented dry sausages manufactured in Italy and other Mediterranean countries: An overview. Int. Food Res. J. 2016, 23, 429–445. [Google Scholar]
- Martín, B.; Garriga, M.; Hugas, M.; Bover-Cid, S.; Veciana-Nogués, M.T.; Aymerich, T. Molecular, technological and safety characterization of Gram-positive catalase-positive cocci from slightly fermented sausages. Int. J. Food Microbiol. 2006, 107, 148–158. [Google Scholar] [CrossRef]
- Sánchez Mainar, M.; Stavropoulou, D.A.; Leroy, F. Exploring the metabolic heterogeneity of coagulase-negative staphylococci to improve the quality and safety of fermented meats: A review. Int. J. Food Microbiol. 2017, 247, 24–37. [Google Scholar] [CrossRef]
- Fontana, C.; Bassi, D.; López, C.; Pisacane, V.; Otero, M.C.; Puglisi, E.; Rebecchi, A.; Cocconcelli, P.S.; Vignolo, G. Microbial ecology involved in the ripening of naturally fermented llama meat sausages. A focus on lactobacilli diversity. Int. J. Food Microbiol. 2016, 236, 17–25. [Google Scholar] [CrossRef]
- Blaiotta, G.; Pennacchia, C.; Villani, F.; Ricciardi, A.; Tofalo, R.; Parente, E. Diversity and dynamics of communities of coagulase-negative staphylococci in traditional fermented sausages. J. Appl. Microbiol. 2004, 97, 271–284. [Google Scholar] [CrossRef]
- Morot-Bizot, S.C.; Talon, R.; Leroy, S. Development of a multiplex PCR for the identification of Staphylococcus genus and four staphylococcal species isolated from food. J. Appl. Microbiol. 2004, 97, 1087–1094. [Google Scholar] [CrossRef]
- Fontana, C.; Cocconcelli, P.S.; Vignolo, G. Monitoring the bacterial population dynamics during fermentation of artisanal Argentinean sausages. Int. J. Food Microbiol. 2005, 103, 131–142. [Google Scholar] [CrossRef]
- Mauriello, G.; Casaburi, A.; Villani, F. Proteolytic activity of Staphylococcus xylosus strains on pork myofibrillar and sarcoplasmic proteins and use of selected strains in the production of “Naples type” salami. J. Appl. Microbiol. 2002, 92, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Landeta, G.; Curiel, J.A.; Carrascosa, A.V.; Muñoz, R.; de las Rivas, B. Characterization of coagulase-negative staphylococci isolated from Spanish dry cured meat products. Meat Sci. 2013, 93, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Casaburi, A.; Blaiotta, G.; Mauriello, G.; Pepe, O.; Villani, F. Technological activities of Staphylococcus carnosus and Staphylococcus simulans strains isolated from fermented sausages. Meat Sci. 2005, 71, 643–650. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- Jeong, D.W.; Han, S.; Lee, J.H. Safety and technological characterization of Staphylococcus equorum isolates from jeotgal, a Korean high-salt-fermented seafood, for starter development. Int. J. Food Microbiol. 2014, 188, 108–115. [Google Scholar] [CrossRef]
- Zell, C.; Resch, M.; Rosenstein, R.; Albrecht, T.; Hertel, C.; Götz, F. Characterization of toxin production of coagulase-negative staphylococci isolated from food and starter cultures. Int. J. Food Microbiol. 2008, 127, 246–251. [Google Scholar] [CrossRef]
- Kwasny, S.M.; Opperman, T.J. Static Biofilm Cultures of Gram Positive. Curr. Protoc. Pharmacol. 2010, 50, 1–27. [Google Scholar]
- Asai, K.; Yamada, K.; Yagi, T.; Baba, H.; Kawamura, I.; Ohta, M. Effect of incubation atmosphere on the production and composition of staphylococcal biofilms. J. Infect. Chemother. 2015, 21, 55–61. [Google Scholar] [CrossRef]
- Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012; Volume 32, ISBN 1562387855.
- Połka, J.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Bacterial diversity in typical Italian salami at different ripening stages as revealed by high-throughput sequencing of 16S rRNA amplicons. Food Microbiol. 2015, 46, 342–356. [Google Scholar] [CrossRef]
- Fonseca, S.; Cachaldora, A.; Gómez, M.; Franco, I.; Carballo, J. Monitoring the bacterial population dynamics during the ripening of Galician chorizo, a traditional dry fermented Spanish sausage. Food Microbiol. 2013, 33, 77–84. [Google Scholar] [CrossRef]
- Cachaldora, A.; García, G.; Lorenzo, J.M.; García-Fontán, M.C. Effect of modified atmosphere and vacuum packaging on some quality characteristics and the shelf-life of “morcilla”, a typical cooked blood sausage. Meat Sci. 2013, 93, 220–225. [Google Scholar] [CrossRef]
- Corbiere Morot-Bizot, S.; Leroy, S.; Talon, R. Monitoring of staphylococcal starters in two French processing plants manufacturing dry fermented sausages. J. Appl. Microbiol. 2007, 102, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Mainar, M.; Xhaferi, R.; Samapundo, S.; Devlieghere, F.; Leroy, F. Opportunities and limitations for the production of safe fermented meats without nitrate and nitrite using an antibacterial Staphylococcus sciuri starter culture. Food Control 2016, 69, 267–274. [Google Scholar] [CrossRef]
- Bermúdez, R.; Lorenzo, J.M.; Fonseca, S.; Franco, I.; Carballo, J. Strains of Staphylococcus and Bacillus Isolated from Traditional Sausages as Producers of Biogenic Amines. Front. Microbiol. 2012, 3, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaia, C.M.; Gouveia, I.M.; Fernandes, M.H.; Fernandes, M.J.; Semedo-Lemsaddek, T.; Barreto, A.S.; Fraqueza, M.J. Assessment of Coagulase-Negative Staphylococci and Lactic Acid Bacteria Isolated from Portuguese Dry Fermented Sausages as Potential Starters Based on Their Biogenic Amine Profile. J. Food Sci. 2018, 83, 2544–2549. [Google Scholar] [CrossRef]
- Sun, J.; Cao, C.C.; Feng, M.Q.; Xu, X.L.; Zhou, G. hong Technological and safety characterization of coagulase-negative staphylococci with high protease activity isolated from Traditional Chinese fermented sausages. LWT 2019, 114, 1–6. [Google Scholar] [CrossRef]
- Alvarez-Ordóñez, A.; Coughlan, L.M.; Briandet, R.; Cotter, P.D. Biofilms in Food Processing Environments: Challenges and Opportunities. Annu. Rev. Food Sci. Technol. 2019, 10, 173–195. [Google Scholar] [CrossRef]
- Khusro, A.; Aarti, C.; Dusthackeer, A.; Agastian, P. Anti-Pathogenic and Technological Traits of Coagulase-Negative Staphylococci Isolated from Koozh, a Fermented Food Product of South India. Food Biotechnol. 2018, 32, 286–304. [Google Scholar] [CrossRef]
- Potter, A.; Ceotto, H.; Giambiagi-deMarval, M.; dos Santos, K.R.N.; Nes, I.F.; Bastos, M.; de Freire Bastos, M.D.C. The gene bap, involved in biofilm production, is present in Staphylococcus spp. strains from nosocomial infections. J. Microbiol. 2009, 47, 319–326. [Google Scholar] [CrossRef]
- Talon, R.; Leroy, S. Diversity and safety hazards of bacteria involved in meat fermentations. Meat Sci. 2011, 89, 303–309. [Google Scholar] [CrossRef]
- Even, S.; Leroy, S.; Charlier, C.; Zakour, N.B.; Chacornac, J.P.; Lebert, I.; Jamet, E.; Desmonts, M.H.; Coton, E.; Pochet, S.; et al. Low occurrence of safety hazards in coagulase negative staphylococci isolated from fermented foodstuffs. Int. J. Food Microbiol. 2010, 139, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Soares Casaes Nunes, R.; Mere Del Aguila, E.; Paschoalin, V.M.F. Safety evaluation of the coagulase-negative staphylococci microbiota of salami: Superantigenic toxin production and antimicrobial resistance. Biomed Res. Int. 2015, 2015, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, P.; Barragán, I.; Seseña, S.; Palop, M.L. Is staphylococci population from milk of healthy goats safe? Int. J. Food Microbiol. 2016, 238, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Rebecchi, A.; Pisacane, V.; Callegari, M.L.; Puglisi, E.; Morelli, L. Ecology of antibiotic resistant coagulase-negative staphylococci isolated from the production chain of a typical Italian salami. Food Control 2015, 53, 14–22. [Google Scholar] [CrossRef]
- Fijałkowski, K.; Peitler, D.; Karakulska, J. Staphylococci isolated from ready-to-eat meat – Identification, antibiotic resistance and toxin gene profile. Int. J. Food Microbiol. 2016, 238, 113–120. [Google Scholar] [CrossRef]
- Marty, E.; Bodenmann, C.; Buchs, J.; Hadorn, R.; Eugster-Meier, E.; Lacroix, C.; Meile, L. Prevalence of antibiotic resistance in coagulase-negative staphylococci from spontaneously fermented meat products and safety assessment for new starters. Int. J. Food Microbiol. 2012, 159, 74–83. [Google Scholar] [CrossRef]
- Pyzik, E.; Marek, A.; Stȩpień-Pyśniak, D.; Urban-Chmiel, R.; Jarosz, L.S.; Jagiełło-Podȩbska, I. Detection of antibiotic resistance and classical enterotoxin genes in coagulase-negative staphylococci isolated from poultry in Poland. J. Vet. Res. 2019, 63, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Chajecka-Wierzchowska, W.; Zadernowska, A.; Nalepa, B.; Sierpińska, M.; Laniewska-Trokenheim, L. Coagulase-negative staphylococci (CoNS) isolated from ready-to-eat food of animal origin - Phenotypic and genotypic antibiotic resistance. Food Microbiol. 2015, 46, 222–226. [Google Scholar] [CrossRef]
- Kamelia, G.; Ali Nazeri, A.; Robabeh, M.; Neda, A.; Zahra, M. Comparison of therapeutic effects of mirtazapine and citalopram on out-patients with major depressive disorder with anxiety symptoms. World J. Pharm. Res. 2015, 4, 346–356. [Google Scholar]
- Genetic Basis of Resistance Waves among Methicillin Resistant Staphylococcus Aureus Isolates Recovered from Milk and Meat Products in Egypt | Ammar | Cellular and Molecular Biology. Available online: https://www.cellmolbiol.org/index.php/CMB/article/view/599 (accessed on 20 January 2020).
- Artini, M.; Cellini, A.; Papa, R.; Tilotta, M.; Scoarughi, G.L.; Gazzola, S.; Fontana, C.; Tempera, G.; Cocconcelli, P.S.; Selan, L. Adhesive behaviour and virulence of coagulase negative staphylococci isolated from Italian cheeses. Int. J. Immunopathol. Pharmacol. 2015, 28, 341–350. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebecchi, A.; Miragoli, F.; Lopez, C.; Bassi, D.; Fontana, C. Exploring Coagulase-Negative Staphylococci Diversity from Artisanal Llama Sausages: Assessment of Technological and Safety Traits. Microorganisms 2020, 8, 629. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050629
Rebecchi A, Miragoli F, Lopez C, Bassi D, Fontana C. Exploring Coagulase-Negative Staphylococci Diversity from Artisanal Llama Sausages: Assessment of Technological and Safety Traits. Microorganisms. 2020; 8(5):629. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050629
Chicago/Turabian StyleRebecchi, Annalisa, Francesco Miragoli, Constanza Lopez, Daniela Bassi, and Cecilia Fontana. 2020. "Exploring Coagulase-Negative Staphylococci Diversity from Artisanal Llama Sausages: Assessment of Technological and Safety Traits" Microorganisms 8, no. 5: 629. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050629