Interaction of Varroa destructor and Sublethal Clothianidin Doses during the Larval Stage on Subsequent Adult Honey Bee (Apis mellifera L.) Health, Cellular Immunity, Deformed Wing Virus Levels and Differential Gene Expression




Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Source of Honeybees and Varroa destructor
2.3. Working Clothianidin Dilutions
2.4. Exposure to Clothianidin and/or V. destructor
2.5. Effect of Clothianidin and/or V. destructor on Bee Emergence and Weight
2.6. Effect of Clothianidin and/or V. destructor on Cellular Immunity
2.7. RNA Extraction
2.8. cDNA Synthesis and DWV Quantification
2.9. RNA Sequencing (RNAseq)
2.10. Statistical Analyses and Bioinformatics
3. Results
3.1. Effect of Clothianidin and/or V. destructor on Bee Emergence and Weight
3.2. Effect of Clothianidin and/or V. destructor on Cellular Immunity
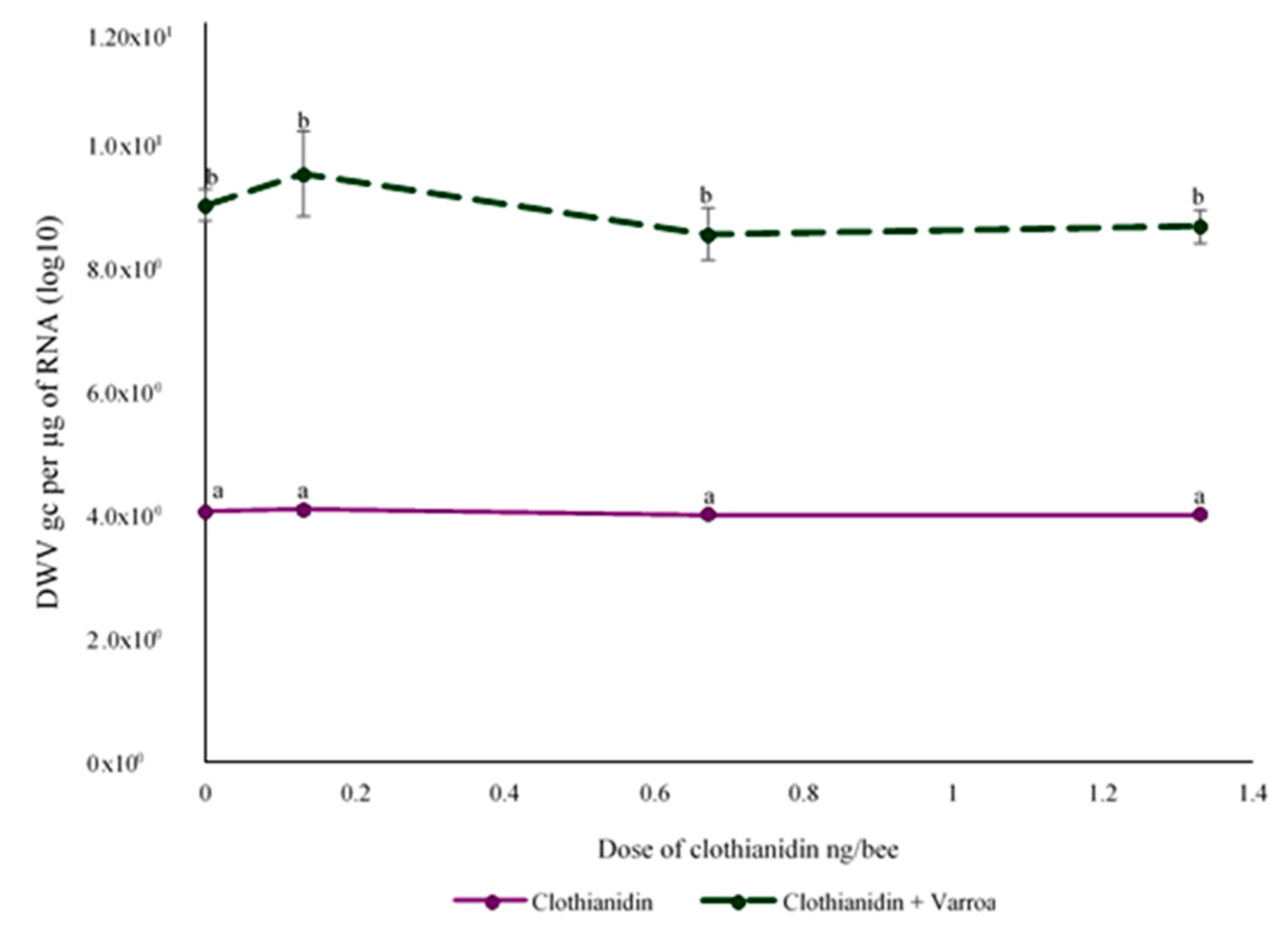
3.3. Effect of Clothianidin and/or V. destructor on DWV Levels
3.4. Comparisons of Up and Down-Regulated DEGs
3.5. KEGG Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Currie, R.; Pernal, S.F.; Guzman-Novoa, E. Honey bee colony losses in Canada. J. Apic. Res. 2010, 49, 104–106. [Google Scholar] [CrossRef]
- Potts, S.; Roberts, S.; Dean, R.; Marris, G.; Brown, M.; Jones, R.; Neumann, P.; Settele, J. Declines of managed honey bees and beekeepers in Europe. J. Apic. Res. 2010, 49, 15–22. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Novoa, E. Colony collapse disorder and other threats to honey bees. In One Health Case Studies, 1st ed.; Susan, C., David, H., Karen, L., Eds.; 5M Publishing Ltd.: Sheffield, UK, 2016; pp. 204–216. [Google Scholar]
- Tomizawa, M.; Casida, J.E. Selective toxicity of neonicotinoids attributable to specificity of insect and mammalian nicotinic receptors. Annu. Rev. Entomol. 2003, 48, 339–364. [Google Scholar] [CrossRef]
- Cresswell, J.E. A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (imidacloprid) on honey bees. Ecotoxicology 2010, 20, 149–157. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Goka, K. Pesticide Residues and Bees—A Risk Assessment. PLoS ONE 2014, 9, e94482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2014, 22, 68–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohorecka, K.; Skubida, P.; Miszczak, A.; Semkiw, P.; Sikorski, P.; Zagibajło, K.; Teper, D.; Koltowski, Z.; Skubida, M.; Zdańska, D.; et al. Residues of neonicotinoid insecticides in bee collected plant materials from oilseed rape crops and their effect on bee colonies. J. Apic. Sci. 2012, 56, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, A.; Evans, J.D.; Scharf, M.; Ellis, J.D. Gene expression in honey bee (Apis mellifera) larvae exposed to pesticides and Varroa mites (Varroa destructor). J. Insect Physiol. 2012, 58, 1042–1049. [Google Scholar] [CrossRef]
- Tasei, J.-N.; Lerin, J.; Ripault, G. Sub-lethal effects of imidacloprid on bumblebees, Bombus terrestris (Hymenoptera: Apidae), during a laboratory feeding test. Pest Manag. Sci. 2000, 56, 784–788. [Google Scholar] [CrossRef]
- Abbott, V.A.; Nadeau, J.L.; Higo, H.A.; Winston, M.L. Lethal and sublethal effects of imidacloprid on Osmia lignaria and clothianidin on Megachile rotundata (Hymenoptera: Megachilidae). J. Encon. Entomol. 2008, 101, 784–796. [Google Scholar] [CrossRef]
- Morfin, N.; Goodwin, P.H.; Correa-Benitez, A.; Guzman-Novoa, E. Sublethal exposure to clothianidin during the larval stage causes long-term impairment of hygienic and foraging behaviours of honey bees. Apidologie 2019, 50, 595–605. [Google Scholar] [CrossRef]
- Sandrock, C.; Tanadini, M.; Tanadini, L.G.; Fauser-Misslin, A.; Potts, S.G.; Neumann, P. Impact of chronic neonicotinoid exposure on honeybee colony performance and queen supersedure. PLoS ONE 2014, 9, e103592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu-Smart, J.; Spivak, M. Sub-lethal effects of dietary neonicotinoid insecticide exposure on honey bee queen fecundity and colony development. Sci. Rep. 2016, 6, 32108. [Google Scholar] [CrossRef] [PubMed]
- Schmuck, R.; Schöning, R.; Stork, A.; Schramel, O. Risk posed to honeybees (Apis mellifera L, Hymenoptera) by an imidacloprid seed dressing of sunflowers. Pest Manag. Sci. 2001, 57, 225–238. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D. Exposure to clothianidin seed-treated canola has no long-term impact on honey bees. J. Econ. Entomol. 2007, 100, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Christen, V.; Bachofer, S.; Fent, K. Binary mixtures of neonicotinoids show different transcriptional changes than single neonicotinoids in honeybees (Apis mellifera). Environ. Pollut. 2017, 220, 1264–1270. [Google Scholar] [CrossRef]
- Tarek, H.; Hamiduzzaman, M.M.; Morfin, N.; Guzman-Novoa, E. Sub-lethal doses of neonicotinoid and carbamate insecticides reduce the lifespan and alter the expression of immune health and detoxification related genes of honey bees (Apis mellifera). Genet. Mol. Res. 2018, 17, 17. [Google Scholar] [CrossRef]
- Morfin, N.; Goodwin, P.H.; Guzman-Novoa, E. Interaction of field realistic doses of clothianidin and Varroa destructor parasitism on adult honey bee (Apis mellifera L.) health and neural gene expression, and antagonistic effects on differentially expressed genes. PLoS ONE 2020, 15, e0229030. [Google Scholar] [CrossRef] [Green Version]
- Boily, M.; Sarrasin, B.; Deblois, C.; Aras, P.; Chagnon, M. Acetylcholinesterase in honey bees (Apis mellifera) exposed to neonicotinoids, atrazine and glyphosate: Laboratory and field experiments. Environ. Sci. Pollut. Res. 2013, 20, 5603–5614. [Google Scholar] [CrossRef]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [Green Version]
- Williams, M. Drosophila hemopoiesis and cellular immunity. J. Immunol. 2007, 178, 4711–4716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Novoa, E.; Morfin, N. Disease Resistance in Honey Bees (Apis mellifera L.) at the Colony and Individual Levels. In Comprehensive Biotechnology, 3rd ed.; Elsevier BV: Amsterdam, The Netherlands, 2019; Volume 4, pp. 811–817. [Google Scholar]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Antiviral defense mechanisms in honey bees. Curr. Opin. Insect Sci. 2015, 10, 71–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigorian, M.; Mandal, L.; Hartenstein, V. Hematopoiesis at the onset of metamorphosis: Terminal differentiation and dissociation of the Drosophila lymph gland. Dev. Genes Evol. 2011, 221, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.; Faria, C.; Jacinto, A. Distinct mechanisms regulate hemocyte chemotaxis during development and wound healing in Drosophila melanogaster. J. Cell Boil. 2006, 173, 405–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson-Rich, N.; Dres, S.T.; Starks, P.T. The ontogeny of immunity: Development of innate immune strength in the honey bee (Apis mellifera). J. Insect Physiol. 2008, 54, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Negri, P.; Maggi, M.; Ramirez, L.; Szawarski, N.; De Feudis, L.; Lamattina, L.; Eguaras, M. Cellular immunity in Apis mellifera: Studying hemocytes brings light about bees skills to confront threats. Apidologie 2015, 47, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Novoa, E.; Eccles, L.; Calvete, Y.; Mcgowan, J.; Kelly, P.G.; Correa-Benítez, A. Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada. Apidologie 2010, 41, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Weinberg, K.P.; Madel, G. The influence of the mite Varroa jacobsoni oud. On the protein concentration and the haemolymph volume of the brood of worker bees and drones of the honey bee Apis mellifera L. Apidologie 1985, 16, 421–436. [Google Scholar] [CrossRef] [Green Version]
- Bowen-Walker, P.L.; Gunn, A. The effect of the ectoparasitic mite, Varroa destructor on adult worker honeybee (Apis mellifera) emergence weights, water, protein, carbohydrate, and lipid levels. Entomol. Exp. et Appl. 2001, 101, 207–217. [Google Scholar] [CrossRef]
- Reyes-Quintana, M.; Espinosa-Montaño, L.G.; Prieto-Merlos, D.; Koleoglu, G.; Petukhova, T.; Correa-Benítez, A.; Guzman-Novoa, E. Impact of Varroa destructor and deformed wing virus on emergence, cellular immunity, wing integrity and survivorship of Africanized honey bees in Mexico. J. Invertebr. Pathol. 2019, 164, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.G.; Evans, J.D.; Rinderer, T.; De Guzman, L. Conditional immune-gene suppression of honeybees parasitized by Varroa mites. J. Insect Sci. 2005, 5, 1–5. [Google Scholar] [CrossRef]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koleoglu, G.; Goodwin, P.H.; Reyes-Quintana, M.; Hamiduzzaman, M.M.; Guzman-Novoa, E. Effect of Varroa destructor, wounding and Varroa homogenate on gene expression in brood and adult honey bees. PLoS ONE 2017, 12, e0169669. [Google Scholar] [CrossRef] [Green Version]
- Koleoglu, G.; Goodwin, P.H.; Reyes-Quintana, M.; Hamiduzzaman, M.M.; Guzmán-Novoa, E. Varroa destructor parasitism reduces hemocyte concentrations and prophenol oxidase gene expression in bees from two populations. Parasitol. Res. 2018, 117, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [Green Version]
- Anguiano-Baez, R.; Guzman-Novoa, E.; Hamiduzzaman, M.M.; Espinosa-Montaño, L.G.; Correa-Benítez, A. Varroa destructor (Mesostigmata: Varroidae) parasitism and climate differentially influence the prevalence, levels, and overt infections of deformed wing virus in honey bees (Hymenoptera: Apidae). J. Insect Sci. 2016, 16, 44. [Google Scholar] [CrossRef] [Green Version]
- Di Prisco, G.; Annoscia, D.; Margiotta, M.; Ferrara, R.; Varricchio, P.; Zanni, V.; Caprio, E.; Nazzi, F.; Pennacchio, F. A mutualistic symbiosis between a parasitic mite and a pathogenic virus undermines honey bee immunity and health. Proc. Natl. Acad. Sci. USA 2016, 113, 3203–3208. [Google Scholar] [CrossRef] [Green Version]
- Alaux, C.; Brunet, J.-L.; Dussaubat, C.; Mondet, F.; Tchamitchan, S.; Cousin, M.; Brillard, J.; Baldy, A.; Belzunces, L.P.; Le Conte, Y. Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera). Environ. Microbiol. 2010, 12, 774–782. [Google Scholar] [CrossRef] [Green Version]
- Vidau, C.; Diogon, M.; Aufauvre, J.; Fontbonne, R.; Viguès, B.; Brunet, J.-L.; Texier, C.; Biron, D.G.; Blot, N.; El Alaoui, H.; et al. Exposure to sublethal doses of fipronil and thiacloprid highly increases mortality of honeybees previously infected by Nosema ceranae. PLoS ONE 2011, 6, e21550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Retschnig, G.; Neumann, P.; Williams, G. Thiacloprid–Nosema ceranae interactions in honey bees: Host survivorship but not parasite reproduction is dependent on pesticide dose. J. Invertebr. Pathol. 2014, 118, 18–19. [Google Scholar] [CrossRef] [PubMed]
- Blanken, L.J.; Van Langevelde, F.; Van Dooremalen, C. Interaction between Varroa destructor and imidacloprid reduces flight capacity of honeybees. Proc. R. Soc. B-Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doublet, V.; Labarussias, M.; De Miranda, J.R.; Moritz, R.; Paxton, R.J. Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 2014, 17, 969–983. [Google Scholar] [CrossRef] [PubMed]
- Morfin, N.; Goodwin, P.H.; Hunt, G.J.; Guzman-Novoa, E. Effects of sublethal doses of clothianidin and/or V. destructor on honey bee (Apis mellifera) self-grooming behavior and associated gene expression. Sci. Rep. 2019, 9, 5196. [Google Scholar] [CrossRef] [Green Version]
- Arechavaleta-Velasco, M.E.; Guzman-Novoa, E. Relative effect of four characteristics that restrain the population growth of the mite Varroa destructor in honey bee (Apis mellifera) colonies. Apidologie 2001, 32, 157–174. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Statement on the findings in recent studies investigating sub-lethal effects in bees of some neonicotinoids in consideration of the uses currently authorised in Europe. EFSA 2012, 10, 2752. [Google Scholar]
- Babendreier, D.; Kalberer, N.M.; Romeis, J.; Fluri, P.; Bigler, F. Pollen consumption in honey bee larvae: A step forward in the risk assessment of transgenic plants. Apidologie 2004, 35, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Hamiduzzaman, M.M.; Sinia, A.; Guzman-Novoa, E.; Goodwin, P.H. Entomopathogenic fungi as potential biocontrol agents of the ecto-parasitic mite, Varroa destructor, and their effect on the immune response of honey bees (Apis mellifera L.). J. Invertebr. Pathol. 2012, 111, 237–243. [Google Scholar] [CrossRef]
- R Studio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA; Available online: http://www.rstudio.com/ (accessed on 9 April 2020).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information, US National Library of Medicine Amel_4.5. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/assembly/GCF_000002195.4/ (accessed on 3 June 2020).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2012, 29, 15–21. [Google Scholar] [CrossRef]
- Roberts, A.; Pimentel, H.; Trapnell, C.; Pachter, L. Identification of novel transcripts in annotated genomes using RNA-Seq. Bioinformatics 2011, 27, 2325–2329. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Theodor Pyl, P.; Wolfgang, H. HTSeq-A Python framework to work with high throughput sequencing data. Bioinformatics 2014. [Google Scholar] [CrossRef]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Bioinformatics and Evolutionary Genomics. Available online: http://bioinformatics.psb.ugent.be/webtools/Venn/ (accessed on 3 June 2020).
- Ramsey, S.; Ochoa, R.; Bauchan, G.R.; Gulbronson, C.; Mowery, J.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, E.H.; Jones, B.; Bowman, A. Salivary secretions from the honeybee mite, Varroa destructor: Effects on insect haemocytes and preliminary biochemical characterization. Parasitology 2011, 138, 602–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Cox-Foster, D. Effects of parasitization by Varroa destructor on survivorship and physiological traits of Apis mellifera in correlation with viral incidence and microbial challenge. Parasitology 2006, 134, 405–412. [Google Scholar] [CrossRef]
- Ritter, W.; Hansen, H. Interactions among Varroa jacobsoni mites, acute paralysis virus, and Paenibacillus larvae larvae and their influence on mortality of larval honeybees in vitro. Apidologie 2000, 31, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Mattos, I.M.; Chaud-Neto, J. Analysis of mortality in Africanized honey bee colonies with high levels of infestation by Varroa destructor. Sociobiology 2014, 59, 369. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Jiang, L.; Wang, H.; Qiao, K.; Wang, N.; Wang, K. Toxicities and sublethal effects of seven neonicotinoid insecticides on survival, growth and reproduction of imidacloprid-resistant cotton aphid, Aphis gossypii. Pest Manag. Sci. 2011, 67, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Jong, D.D.; Morse, R.A.; Eickwort, G.C. Mite pests of honey bees. Annu. Rev. Entomol. 1982, 27, 229–252. [Google Scholar] [CrossRef]
- Salem, M.H.; Abir, A.G.; Ramadan, H. Effect of Varroa destructor on different haemocyte count, total haemolymph protein on larvae, pupae and adults of Apis mellifera drones. J. Egypt. Soc. Toxicol. 2006, 35, 93–96. [Google Scholar]
- Amdam, G.V.; Hartfelder, K.; Norberg, K.; Hagen, A.; Omholt, S.W. Altered physiology in worker honey bees (Hymenoptera: Apidae) infested with the mite Varroa destructor (Acari: Varroidae): A factor in colony loss during overwintering? J. Econom. Entomol. 2004, 97, 741–747. [Google Scholar] [CrossRef]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Büchler, R. The neonicotinoids thiacloprid, imidacloprid, and clothianidin affect the immunocompetence of honey bees (Apis mellifera L.). J. Insect Physiol. 2016, 86, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormesis: A revolution in toxicology, risk assessment and medicine. EMBO Rep. 2004, 5, S37–S40. [Google Scholar] [CrossRef] [Green Version]
- Guedes, N.M.P.; Tolledo, J.; Correa, A.S.; Guedes, R.N.C. Insecticide-induced hormesis in an insecticide-resistant strain of the maize weevil, Sitophilus zeamais. J. Appl. Èntomol. 2010, 134, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Ramanaidu, K.; Cutler, G.C. Different toxic and hormetic responses of Bombus impatiens to Beauveria bassiana, Bacillus subtilis and spirotetramat. Pest Manag. Sci. 2012, 69, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A Virulent strain of deformed wing virus (DWV) of honeybees (Apis mellifera) prevails after Varroa destructor-mediated, or in vitro, transmission. PLoS Pathog. 2014, 10, e1004230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, C.; Genersch, E. RT-PCR analysis of deformed wing virus in honeybees (Apis mellifera) and mites (Varroa destructor). J. Gen. Virol. 2005, 86, 3419–3424. [Google Scholar] [CrossRef] [PubMed]
- Möckel, N.; Gisder, S.; Genersch, E. Horizontal transmission of deformed wing virus: Pathological consequences in adult bees (Apis mellifera) depend on the transmission route. J. Gen. Virol. 2010, 92, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, I. Nicotine to Nicotinoids: 1962 to 1997. In Nicotinoid Insecticides and the Nicotinic Acetylcholine Receptor, 1st ed.; Izuru, Y., John, E.C., Eds.; Springer: Tokyo, Japan, 1999; pp. 3–27. [Google Scholar]
- Shi, T.-F.; Wang, Y.-F.; Liu, F.; Qi, L.; Yu, L. Sublethal effects of the neonicotinoid insecticide thiamethoxam on the transcriptome of the honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 2017, 110, 2283–2289. [Google Scholar] [CrossRef]
- Christen, V.; Schirrmann, M.; Frey, J.E.; Fent, K. Global transcriptomic effects of environmentally relevant concentrations of the neonicotinoids clothianidin, imidacloprid, and thiamethoxam in the brain of honey bees (Apis mellifera). Environ. Sci. Technol. 2018, 52, 7534–7544. [Google Scholar] [CrossRef]
- Elsik, C.G.; Tayal, A.; Diesh, C.M.; Unni, D.R.; Emery, M.L.; Nguyen, H.N.; Hagen, D.E. Hymenoptera Genome Database: Integrating genome annotations in HymenopteraMine. Nucleic Acids Res. 2016, 44, D793–D800. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. g:Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic Acids Res. 2016, 44, W83–W89. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | Mean Proportion of Emerged Bees (± S.E.) |
|---|---|
| 0 ng/bee | 0.98 ± 0.0027 a |
| 0.13 ng/bee | 0.96 ± 0.0069 a |
| 0.67 ng/bee | 0.97 ± 0.0047 a |
| 1.33 ng/bee | 0.95 ± 0.0050 a |
| 0 ng/bee + V. destructor | 0.68 ± 0.083 b |
| 0.13 ng/bee + V. destructor | 0.65 ± 0.12 b |
| 0.67 ng/bee + V. destructor | 0.56 ± 0.12 b |
| 1.33 ng/bee + V. destructor | 0.60 ± 0.10 b |
| Treatment | Mean Weight of Newly Emerged Bees (mg ± S.E.) |
|---|---|
| 0 ng/bee | 121.76 ±1.21 a,b |
| 0.13 ng/bee | 122.89 ±1.09 a |
| 0.67 ng/bee | 120.81 ± 1.16 a,b |
| 1.33 ng/bee | 117.76 ± 1.42 b,c |
| 0 ng/bee + V. destructor | 111.82 ± 1.09 d |
| 0.13 ng/bee + V. destructor | 109.68 ± 0.94 e,d |
| 0.67 ng/bee + V. destructor | 113.67 ± 1.05 c,e |
| 1.33 ng/bee + V. destructor | 111.38 ± 0.92 e,d |
| Treatment | Haemocytes/µL of Haemolymph (± S.E.) |
|---|---|
| 0 ng/bee | 16077.93 ± 920.24 a |
| 0.13 ng/bee | 23645.80 ± 1695.45 b |
| 0.67 ng/bee | 18058.90 ± 943.02 a |
| 1.33 ng/bee | 17321.6 ± 934.07 a |
| 0 ng/bee + V. destructor | 12445.00 ± 1209.67 c |
| 0.13 ng/bee + V. destructor | 12571.70 ± 1105.83 c |
| 0.67 ng/bee + V. destructor | 12962.70 ± 1307.27 c |
| 1.33 ng/bee+ V. destructor | 11998.00 ± 1094.89 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morfin, N.; Goodwin, P.H.; Guzman-Novoa, E. Interaction of Varroa destructor and Sublethal Clothianidin Doses during the Larval Stage on Subsequent Adult Honey Bee (Apis mellifera L.) Health, Cellular Immunity, Deformed Wing Virus Levels and Differential Gene Expression. Microorganisms 2020, 8, 858. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8060858
Morfin N, Goodwin PH, Guzman-Novoa E. Interaction of Varroa destructor and Sublethal Clothianidin Doses during the Larval Stage on Subsequent Adult Honey Bee (Apis mellifera L.) Health, Cellular Immunity, Deformed Wing Virus Levels and Differential Gene Expression. Microorganisms. 2020; 8(6):858. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8060858
Chicago/Turabian StyleMorfin, Nuria, Paul H. Goodwin, and Ernesto Guzman-Novoa. 2020. "Interaction of Varroa destructor and Sublethal Clothianidin Doses during the Larval Stage on Subsequent Adult Honey Bee (Apis mellifera L.) Health, Cellular Immunity, Deformed Wing Virus Levels and Differential Gene Expression" Microorganisms 8, no. 6: 858. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8060858