Characterization of Bacteriophages against Pseudomonas Syringae pv. Actinidiae with Potential Use as Natural Antimicrobials in Kiwifruit Plants

 , , and
, , and 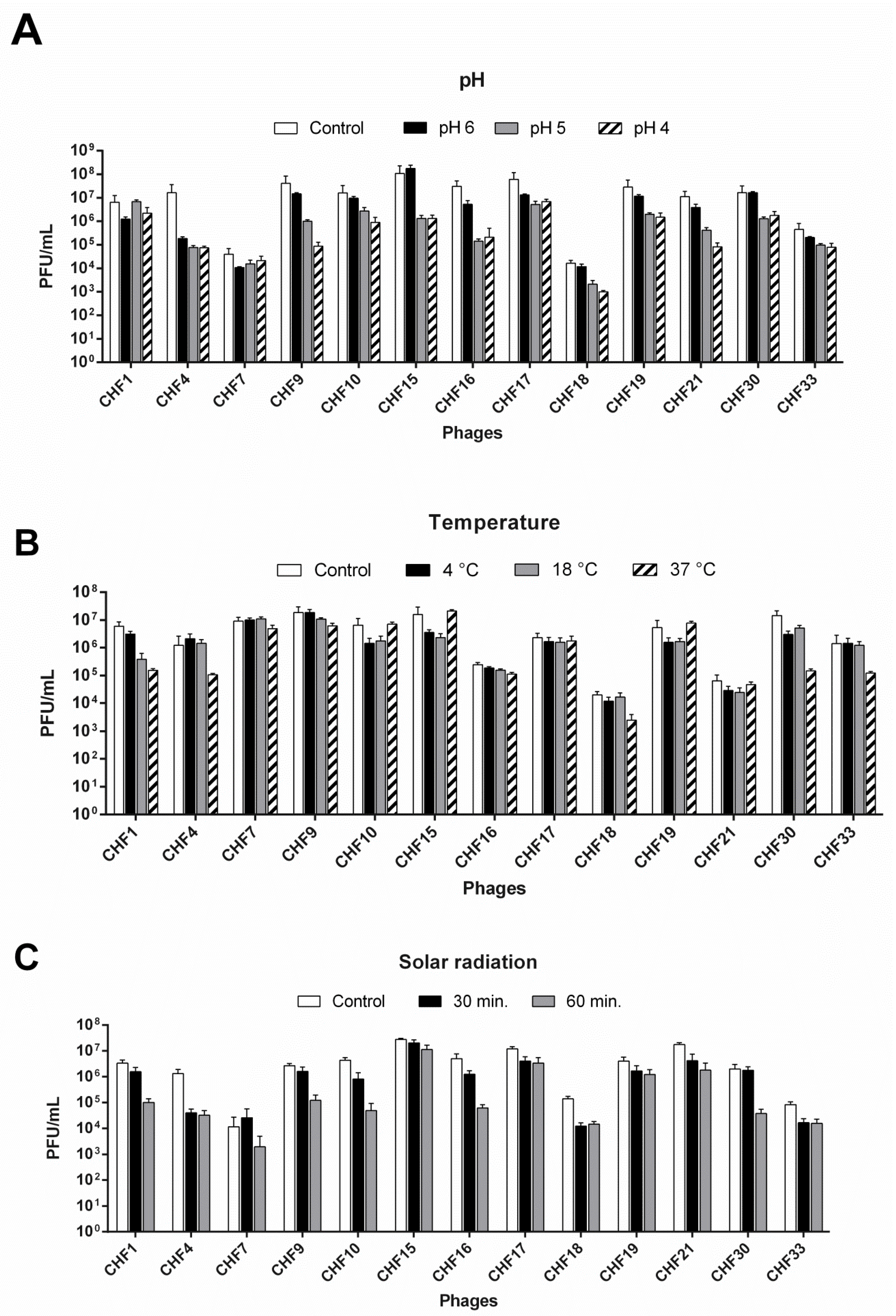{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Bacteriophage Isolation, Propagation, and Concentration
2.3. Bacteriophage Viability under Different Conditions
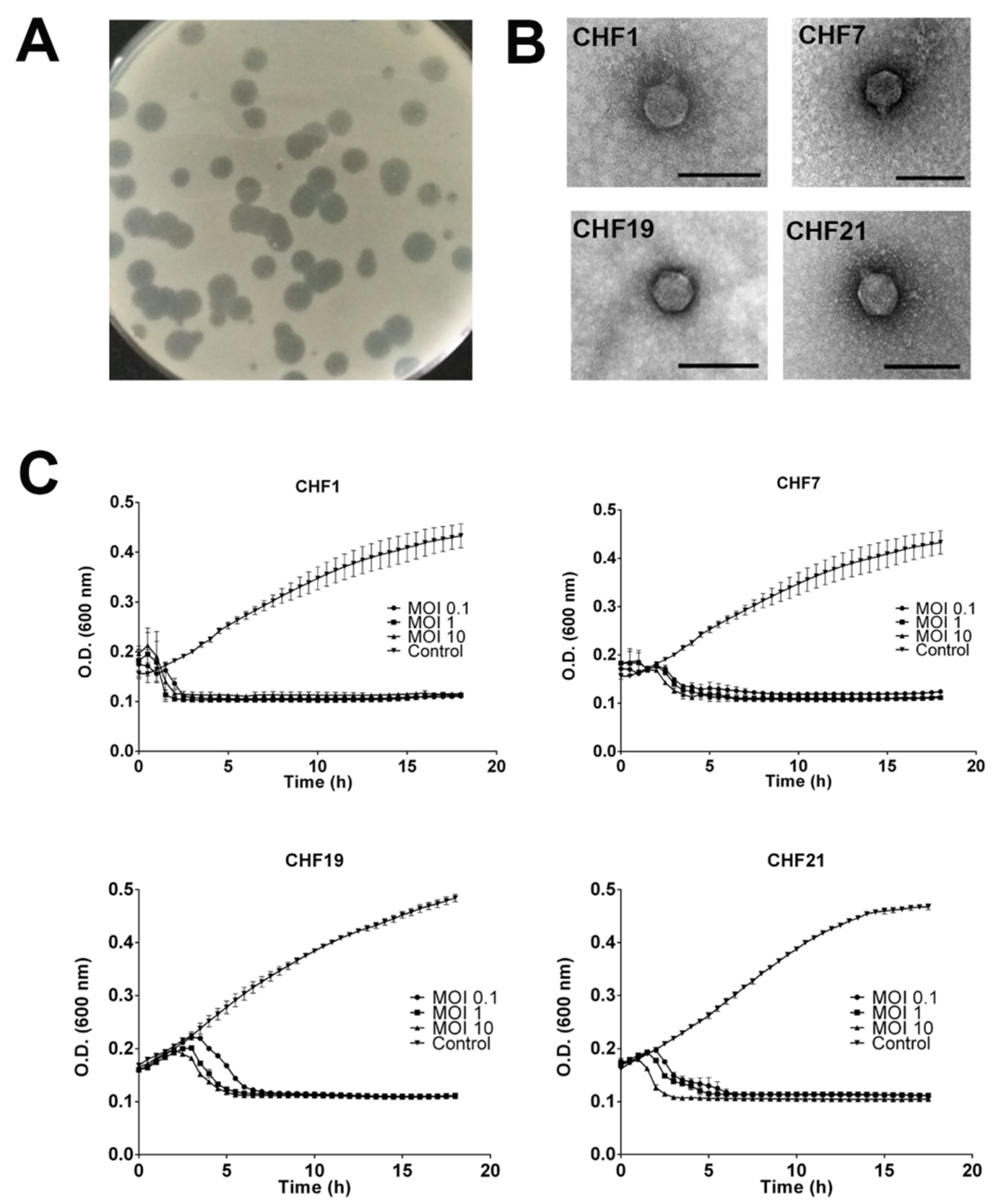
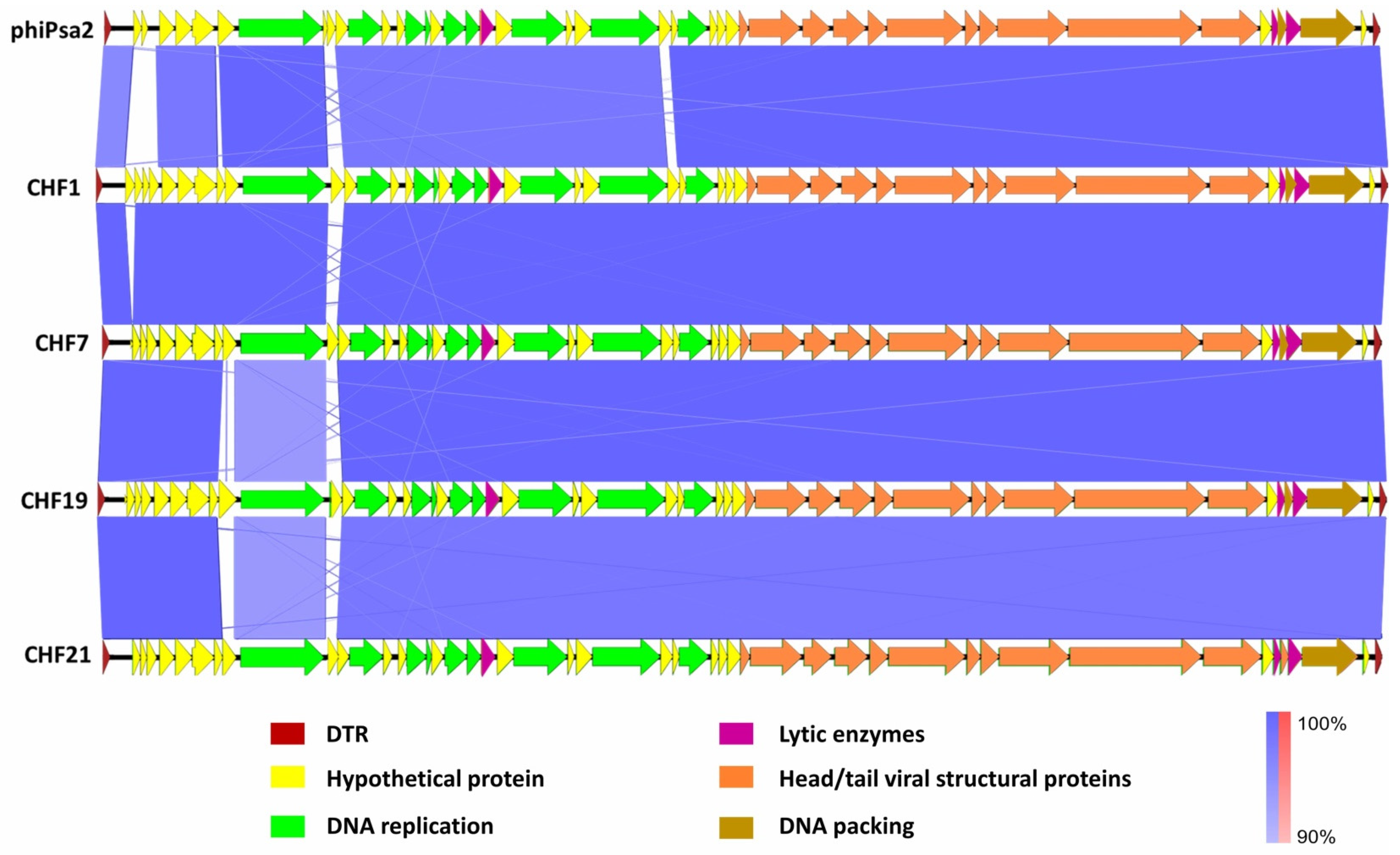
2.4. Molecular and Morphological Characterization of Bacteriophages
2.5. Infection Curves, Bacteriophage Host Range Analysis, and Resistance Frequency
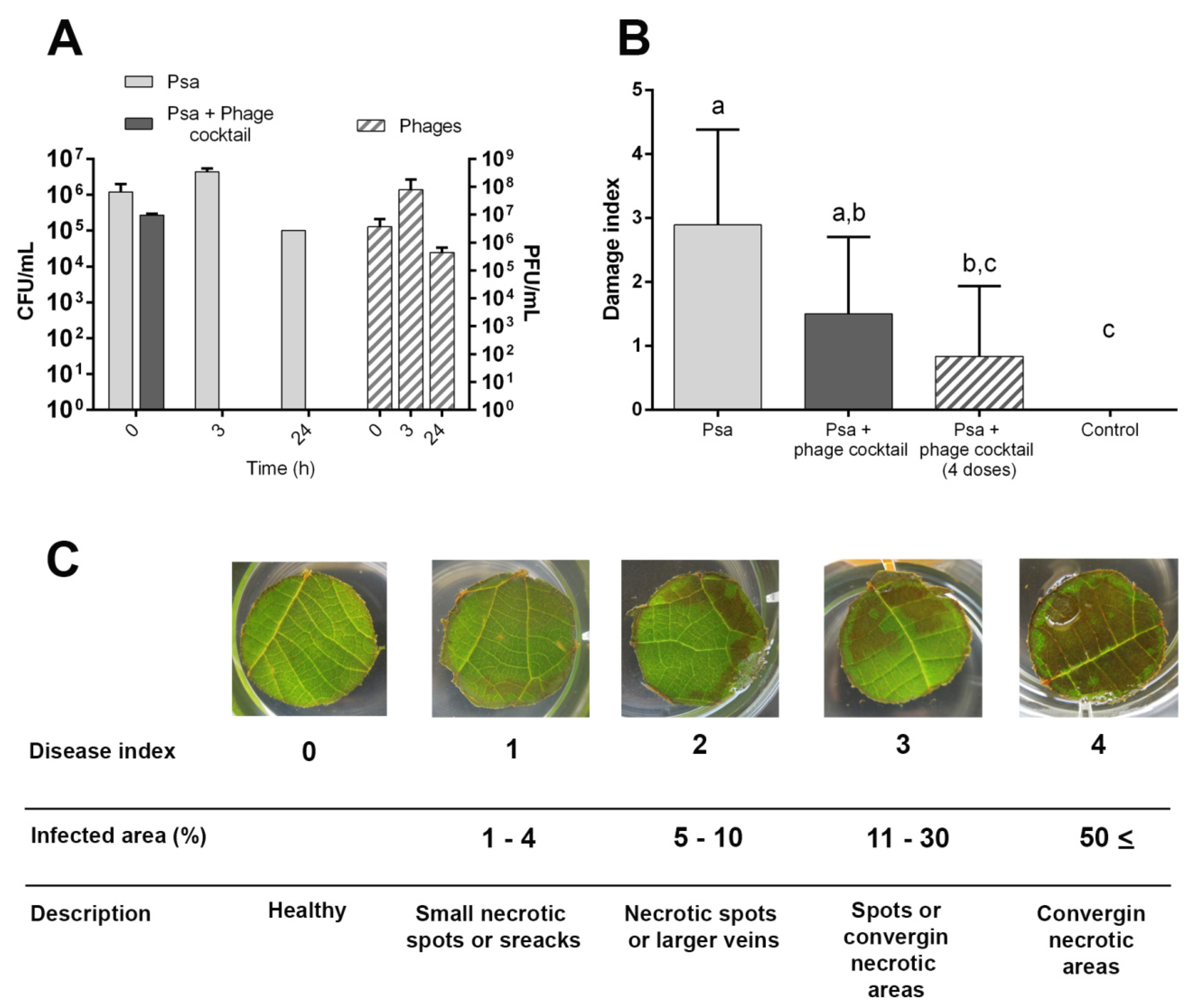
2.6. Efficacy of Bacteriophage Cocktail in Psa Control on Kiwifruit
3. Results
3.1. Isolation and Selection of Bacteriophages against Pseudomonas syringae pv. actinidiae
3.2. Genomic and Phenotypic Characterization of Selected Psa Bacteriophages
3.3. In Vitro Efficacy of Bacteriophages to Control Psa
3.4. Specificity of Chilean Psa Bacteriophages
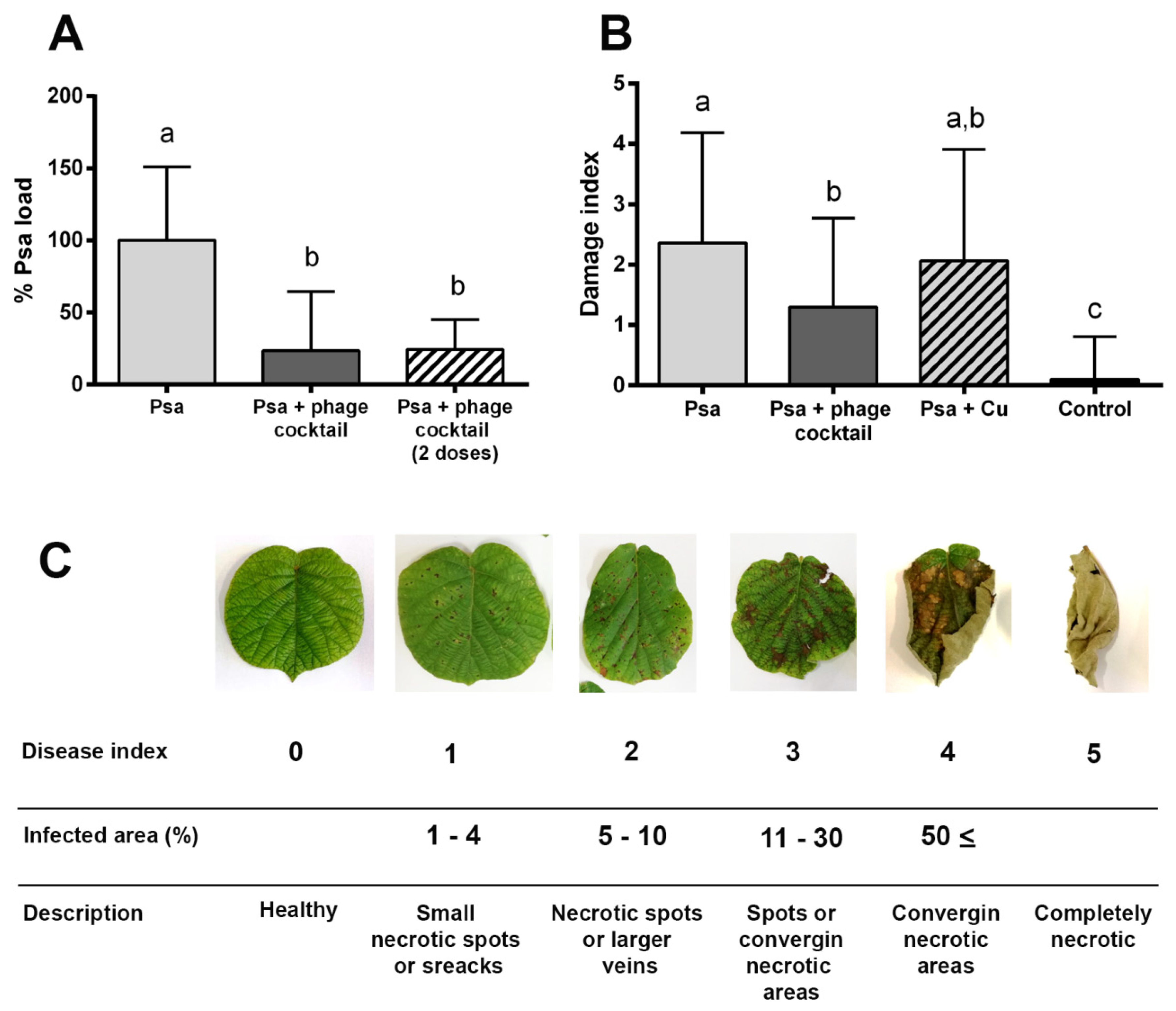
3.5. In Vivo Efficacy of Bacteriophages to Biocontrol Psa under Greenhouse Conditions
4. Discussion
4.1. Phages against Psa
4.2. Phages as Biological Control Agents against Psa
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ciacci, C.; Russo, I.; Bucci, C.; Iovino, P.; Pellegrini, L.; Giangrieco, I.; Tamburrini, M.; Ciardiello, M.A. The kiwi fruit peptide kissper displays anti-inflammatory and anti-oxidant effects in in-vitro and ex-vivo human intestinal models. Clin. Exp. Immunol. 2014, 175, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.N.; Meng, X.; Li, Y.; Li, S.; Liu, Q.; Tang, G.Y.; Li, H.B. Fruits for prevention and treatment of cardiovascular diseases. Nutrients 2017, 9, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guroo, I.; Wani, S.A.; Wani, S.M.; Ahmad, M.; Mir, S.A.; Masoodi, F.A. A Review of Production and Processing of Kiwifruit. J. Food Process. Technol. 2017, 8. [Google Scholar] [CrossRef]
- Wicaksono, W.A.; Jones, E.E.; Casonato, S.; Monk, J.; Ridgway, H.J. Biological control of Pseudomonas syringae pv. actinidiae (Psa), the causal agent of bacterial canker of kiwifruit, using endophytic bacteria recovered from a medicinal plant. Boil. Control. 2018, 116, 103–112. [Google Scholar] [CrossRef]
- McCann, H.C.; Li, L.; Liu, Y.; Li, D.; Pan, H.; Zhong, C.; Rikkerink, E.H.; Templeton, M.D.; Straub, C.; Colombi, E.; et al. Origin and Evolution of the Kiwifruit Canker Pandemic. Genome Boil. Evol. 2017, 9, 932–944. [Google Scholar] [CrossRef]
- Ferrante, P.; Scortichini, M. Redefining the global populations of Pseudomonas syringaepv. actinidiaebased on pathogenic, molecular and phenotypic characteristics. Plant Pathol. 2014, 64, 51–62. [Google Scholar] [CrossRef]
- Fujikawa, T.; Sawada, H. Genome analysis of Pseudomonas syringae pv. actinidiae biovar 6, which produces the phytotoxins, phaseolotoxin and coronatine. Sci. Rep. 2019, 9, 3836. [Google Scholar] [CrossRef]
- ODEPA. CHILE Estadísticas Productivas Sector Frutícola. Available online: https://www.odepa.gob.cl (accessed on 15 March 2020).
- Cameron, A.J.; Sarojini, V. Pseudomonas syringae pv. actinidiae: Chemical control, resistance mechanisms and possible alternatives. Plant Pathol. 2013, 63, 1–11. [Google Scholar] [CrossRef]
- Nakajima, M.; Goto, M.; Hibi, T. Similarity between Copper Resistance Genes from Pseudomonas syringae pv. actinidiae and P. syringae pv. tomato. J. Gen. Plant Pathol. 2002, 68, 68–74. [Google Scholar] [CrossRef]
- Vanneste, J.; Voyle, M. Genetic basis of copper resistance in New Zealand strains of Pseudomonas syringae. N. Z. Plant Prot. 2003, 56, 109–112. [Google Scholar] [CrossRef]
- Colombi, E.; Straub, C.; Künzel, S.; Templeton, M.D.; McCann, H.C.; Rainey, P.B. Evolution of copper resistance in the kiwifruit pathogenPseudomonas syringaepv.actinidiaethrough acquisition of integrative conjugative elements and plasmids. Environ. Microbiol. 2017, 19, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.T.; Naidu, R.; Ming, H.; Megharaj, M. Copper phytotoxicity in native and agronomical plant species. Ecotoxicol. Environ. Saf. 2012, 85, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Donati, I.; Buriani, G.; Cellini, A.; Raule, N.; Spinelli, F. Screening of microbial biocoenosis of Actinidia chinensis for the isolation of candidate biological control agents against Pseudomonas syringae pv. actinidiae. Acta Hortic. 2018, 239–246. [Google Scholar] [CrossRef]
- Gould, E.; Black, M.; Clark, G.; Tanner, D.; Benge, J. Tools for managing the kiwifruit bacterial canker diseasePseudomonas syringaepvactinidiae(Psa). Acta Hortic. 2015, 39–46. [Google Scholar] [CrossRef]
- Tontou, R.; Giovanardi, D.; Ferrari, M.; Stefani, E. Isolation of bacterial endophytes from Actinidia chinensis and preliminary studies on their possible use as antagonists against Pseudomonas syringae pv. actinidiae. J. Berry Res. 2016, 6, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Daranas, N.; Roselló, G.; Cabrefiga, J.; Donati, I.; Francés, J.; Badosa, E.; Spinelli, F.; Montesinos, E.; Bonaterra, A. Biological control of bacterial plant diseases with Lactobacillus plantarum strains selected for their broad-spectrum activity. Ann. Appl. Boil. 2018, 174, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Bardin, M.; Ajouz, S.; Comby, M.; Lopez-Ferber, M.; Graillot, B.; Siegwart, M.; Nicot, P.C. Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Monchiero, M.; Gullino, M.; Pugliese, M.; Spadaro, D.; Garibaldi, A. Efficacy of different chemical and biological products in the control of Pseudomonas syringae pv. actinidiae on kiwifruit. Australas. Plant Pathol. 2014, 44, 13–23. [Google Scholar] [CrossRef]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and Bacterial Plant Diseases. Front. Microbiol. 2017, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Svircev, A.; Roach, D.R.; Castle, A. Framing the Future with Bacteriophages in Agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [Green Version]
- Doss, J.; Culbertson, K.; Hahn, D.; Camacho, J.; Barekzi, N. A Review of Phage Therapy against Bacterial Pathogens of Aquatic and Terrestrial Organisms. Viruses 2017, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rombouts, S.; Volckaert, A.; Venneman, S.; Declercq, B.; Vandenheuvel, D.; Allonsius, C.N.; Van Malderghem, C.; Jang, H.B.; Briers, Y.; Noben, J.-P.; et al. Characterization of Novel Bacteriophages for Biocontrol of Bacterial Blight in Leek Caused by Pseudomonas syringae pv. porri. Front. Microbiol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuppels, D.A. Isolation and Characterization of Phages Useful for Identifying Pseudomonas syringae pv. tomato. Phytopathol. 1983, 73, 1376. [Google Scholar] [CrossRef]
- Smith, A.R.W.; Zamze, S.E.; Hignett, R.C. Morphology and hydrolytic activity of A7, a typing phage of Pseudomonas syringae pv. morsprunorum. Microbiol. 1994, 140, 905–913. [Google Scholar] [CrossRef] [Green Version]
- Di Lallo, G.; Evangelisti, M.; Mancuso, F.; Ferrante, P.; Marcelletti, S.; Tinari, A.; Superti, F.; Migliore, L.; D’Addabbo, P.; Frezza, D.; et al. Isolation and partial characterization of bacteriophages infecting Pseudomonas syringae pv. actinidiae, causal agent of kiwifruit bacterial canker. J. Basic Microbiol. 2014, 54, 1210–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frampton, R.A.; Taylor, C.; Moreno, A.V.H.; Visnovsky, S.B.; Petty, N.K.; Pitman, A.R.; Fineran, P.C. Identification of Bacteriophages for Biocontrol of the Kiwifruit Canker Phytopathogen Pseudomonas syringae pv. actinidiae. Appl. Environ. Microbiol. 2014, 80, 2216–2228. [Google Scholar] [CrossRef] [Green Version]
- Wojtus, J.; Frampton, R.A.; Warring, S.; Hendrickson, H.L.; Fineran, P.C. Genome Sequence of a Jumbo Bacteriophage That Infects the Kiwifruit Phytopathogen Pseudomonas syringae pv. actinidiae. Microbiol. Resour. Announc. 2019. [Google Scholar] [CrossRef] [Green Version]
- Frampton, R.A.; López-Acedo, E.; Young, V.L.; Chen, D.; Tong, B.; Taylor, C.; Easingwood, R.A.; Pitman, A.R.; Kleffmann, T.; Bostina, M.; et al. Genome, Proteome and Structure of a T7-Like Bacteriophage of the Kiwifruit Canker Phytopathogen Pseudomonas syringae pv. actinidiae. Viruses 2015, 7, 3361–3379. [Google Scholar] [CrossRef]
- Yu, J.-G.; Lim, J.-A.; Song, Y.-R.; Heu, S.; Kim, G.H.; Koh, Y.J.; Oh, C.-S. Isolation and Characterization of Bacteriophages Against Pseudomonas syringae pv. actinidiae Causing Bacterial Canker Disease in Kiwifruit. J. Microbiol. Biotechnol. 2016, 26, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Ni, P.; Deng, B.; Wang, S.; Xu, W.; Wang, D. Isolation and characterisation of phages against Pseudomonas syringae pv. actinidiae. Acta Agric. Scand. Sect. B Plant Soil Sci. 2018, 69, 199–208. [Google Scholar] [CrossRef]
- Pinheiro, L.A.M.; Pereira, C.; Barreal, M.E.; Gallego, P.P.; Balcão, V.M.C.F.; Almeida, A. Use of phage ϕ6 to inactivate Pseudomonas syringae pv. actinidiae in kiwifruit plants: In vitro and ex vivo experiments. Appl. Microbiol. Biotechnol. 2019, 104, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Flores, O.; Prince, C.; Nuñez, M.; Vallejos, A.; Mardones, C.; Yañez, C.; Besoain, X.; Bastias, R. Genetic and Phenotypic Characterization of Indole-Producing Isolates of Pseudomonas syringae pv. actinidiae Obtained from Chilean Kiwifruit Orchards. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Kutter, E. Phage Host Range and Efficiency of Plating. Adv. Struct. Saf. Stud. 2009, 501, 141–149. [Google Scholar] [CrossRef]
- Pavel, A.B.; Vasile, C.I. PyElph—A software tool for gel images analysis and phylogenetics. BMC Bioinform. 2012, 13, 9. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- León, M.; Kokkari, C.; García, K.; Castillo, D.; Katharios, P.; Bastias, R. Diversification of Vibrio anguillarum Driven by the Bacteriophage CHOED. Front. Microbiol. 2019, 10, 1396. [Google Scholar] [CrossRef] [Green Version]
- Prencipe, S.; Gullino, M.; Spadaro, D. Pseudomonas syringae pv. actinidiae isolated from Actinidia chinensis Var. deliciosa in Northern Italy: Genetic diversity and virulence. Eur. J. Plant Pathol. 2017, 150, 191–204. [Google Scholar] [CrossRef]
- Collina, M.; Donati, I.; Bertacchini, E.; Brunelli, A.; Spinelli, F. Greenhouse assays on the control of the bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). J. Berry Res. 2016, 6, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.M.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunty, A.; Cesbron, S.; Poliakoff, F.; Jacques, M.-A.; Manceau, C. Origin of the Outbreak in France of Pseudomonas syringae pv. actinidiae Biovar 3, the Causal Agent of Bacterial Canker of Kiwifruit, Revealed by a Multilocus Variable-Number Tandem-Repeat Analysis. Appl. Environ. Microbiol. 2015, 81, 6773–6789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Lim, J.-A.; Yu, J.-G.; Oh, C.-S. Genomic Features and Lytic Activity of the Bacteriophage PPPL-1 Effective against Pseudomonas syringae pv. actinidiae, a Cause of Bacterial Canker in Kiwifruit. J. Microbiol. Biotechnol. 2018, 28, 1542–1546. [Google Scholar] [CrossRef]
- Liu, W.; Lin, Y.-R.; Lu, M.-W.; Sung, P.-J.; Wang, W.-H.; Lin, C.-S. Genome sequences characterizing five mutations in RNA polymerase and major capsid of phages ϕA318 and ϕAs51 of Vibrio alginolyticus with different burst efficiencies. BMC Genom. 2014, 15, 505. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.B.; Vallad, G.E.; Iriarte, F.B.; Obradović, A.; Wernsing, M.H.; Jackson, L.E.; Balogh, B.; Hong, J.C.; Momol, M. Considerations for using bacteriophages for plant disease control. Bacteriophage 2012, 2, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Gasic, K.; Kuzmanović, N.; Ivanović, M.; Prokić, A.; Šević, M.; Obradoviċ, A. Complete Genome of the Xanthomonas euvesicatoria Specific Bacteriophage KΦ1, Its Survival and Potential in Control of Pepper Bacterial Spot. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Jones, J.B.; Jackson, L.; Balogh, B.; Obradovic, A.; Iriarte, F.; Momol, M. Bacteriophages for Plant Disease Control. Annu. Rev. Phytopathol. 2007, 45, 245–262. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores, O.; Retamales, J.; Núñez, M.; León, M.; Salinas, P.; Besoain, X.; Yañez, C.; Bastías, R. Characterization of Bacteriophages against Pseudomonas Syringae pv. Actinidiae with Potential Use as Natural Antimicrobials in Kiwifruit Plants. Microorganisms 2020, 8, 974. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8070974
Flores O, Retamales J, Núñez M, León M, Salinas P, Besoain X, Yañez C, Bastías R. Characterization of Bacteriophages against Pseudomonas Syringae pv. Actinidiae with Potential Use as Natural Antimicrobials in Kiwifruit Plants. Microorganisms. 2020; 8(7):974. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8070974
Chicago/Turabian StyleFlores, Oriana, Julio Retamales, Mauricio Núñez, Marcela León, Paula Salinas, Ximena Besoain, Carolina Yañez, and Roberto Bastías. 2020. "Characterization of Bacteriophages against Pseudomonas Syringae pv. Actinidiae with Potential Use as Natural Antimicrobials in Kiwifruit Plants" Microorganisms 8, no. 7: 974. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8070974