
Interactive Effects of Epichloë Endophyte, Dormancy-Breaking Treatments and Geographic Origin on Seed Germination of Achnatherum inebrians


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Materials
2.2. Breaking Dormancy and Germination Experiment
2.3. Determination of Germination Rate
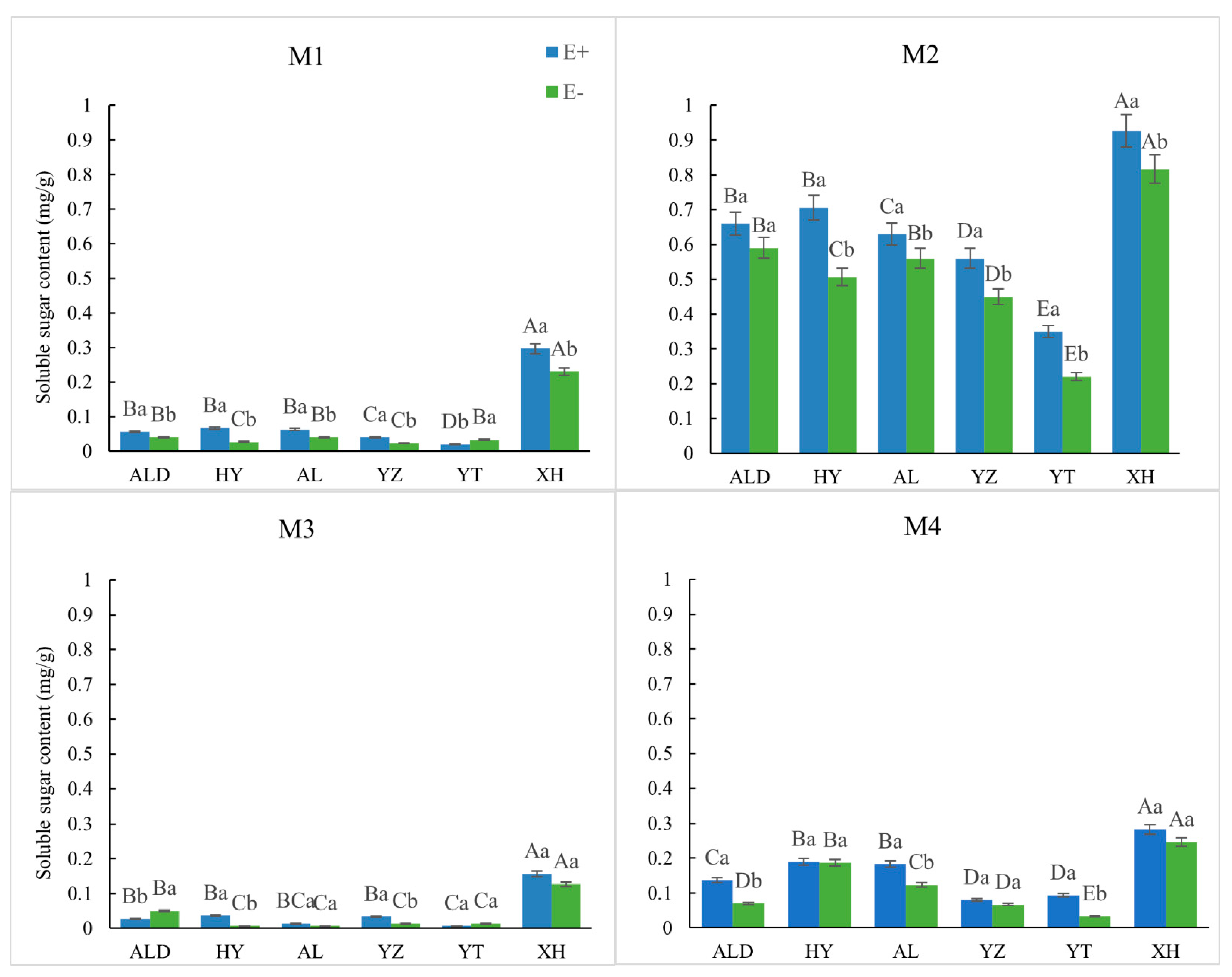
2.4. Determination of Soluble Sugar
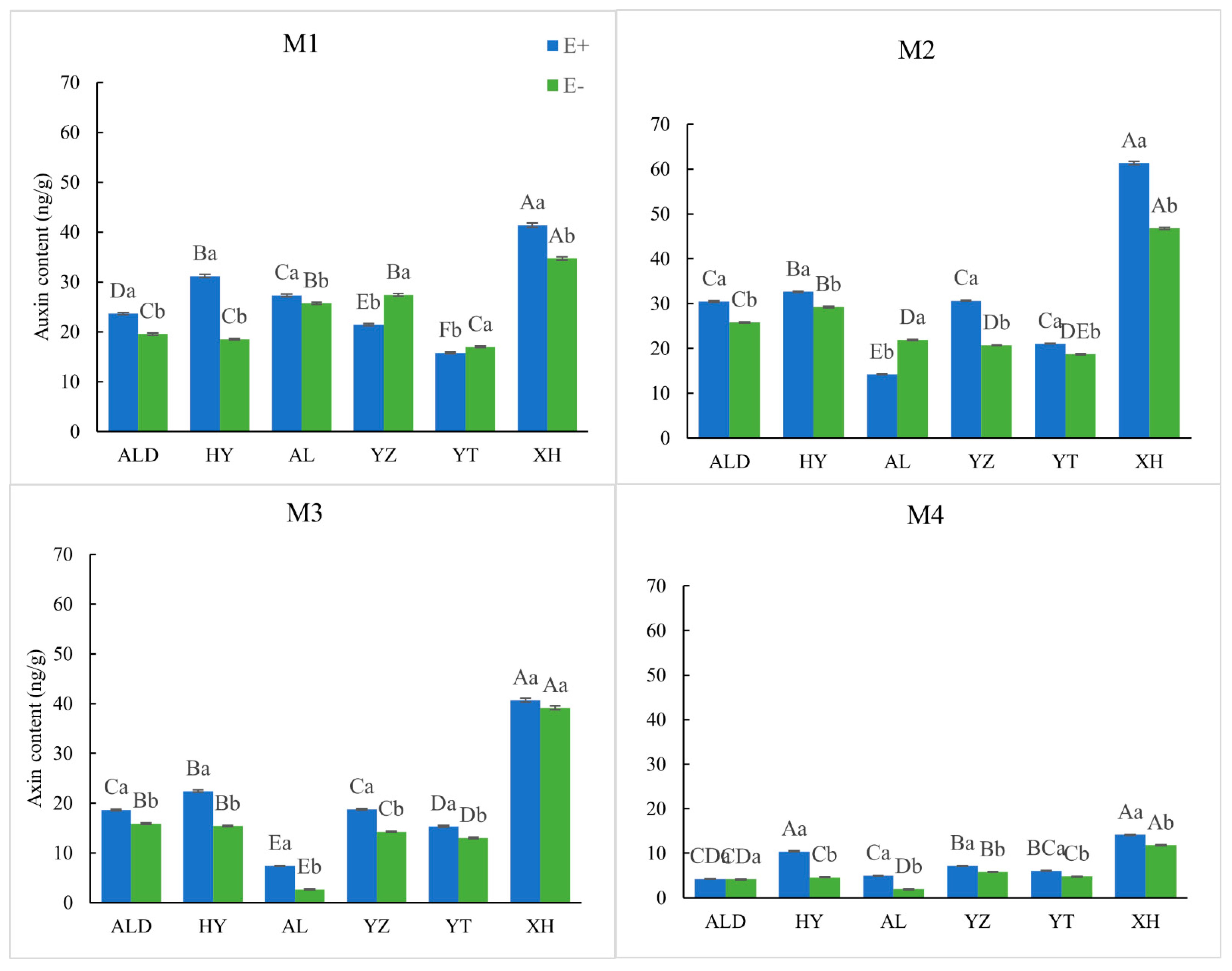
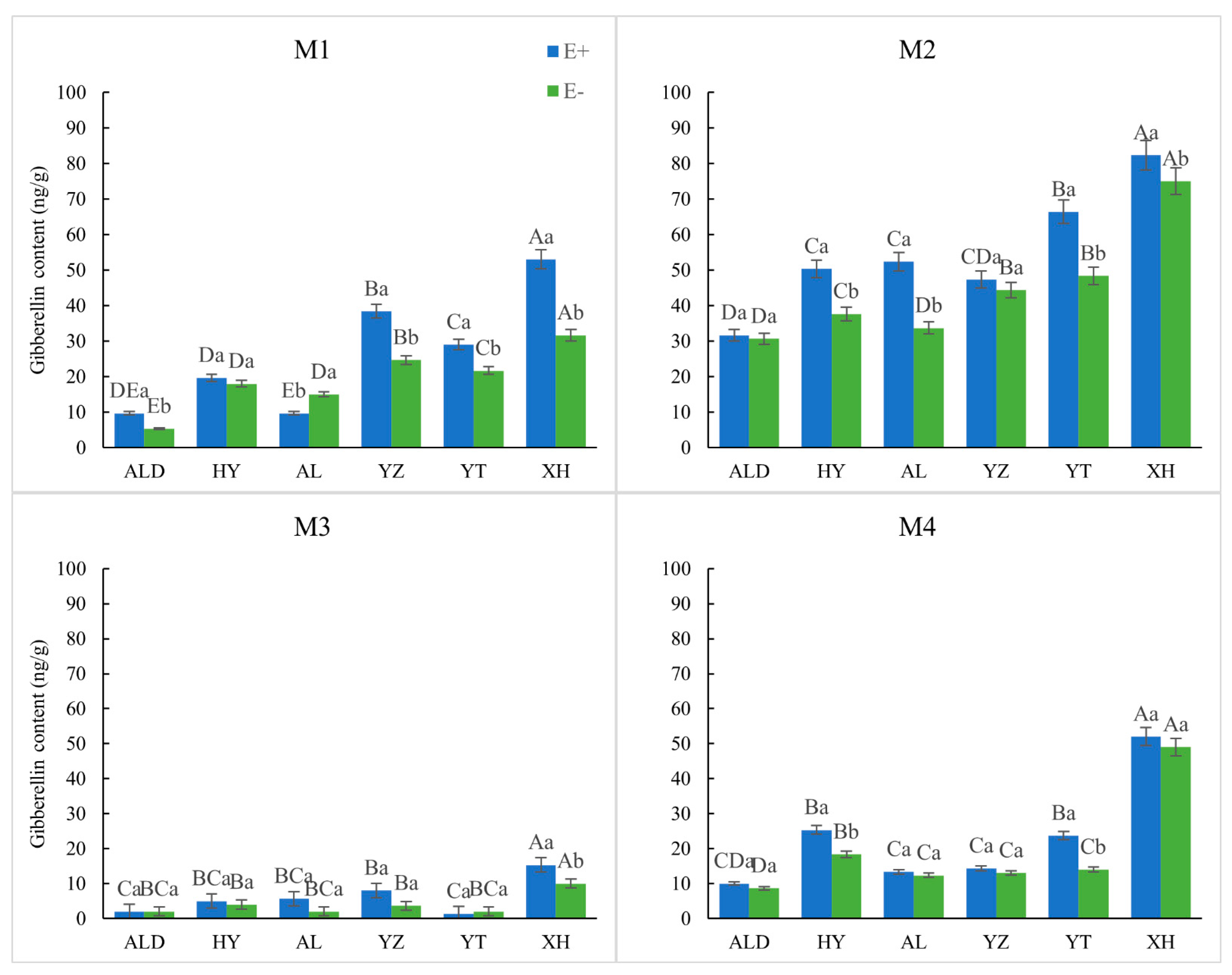
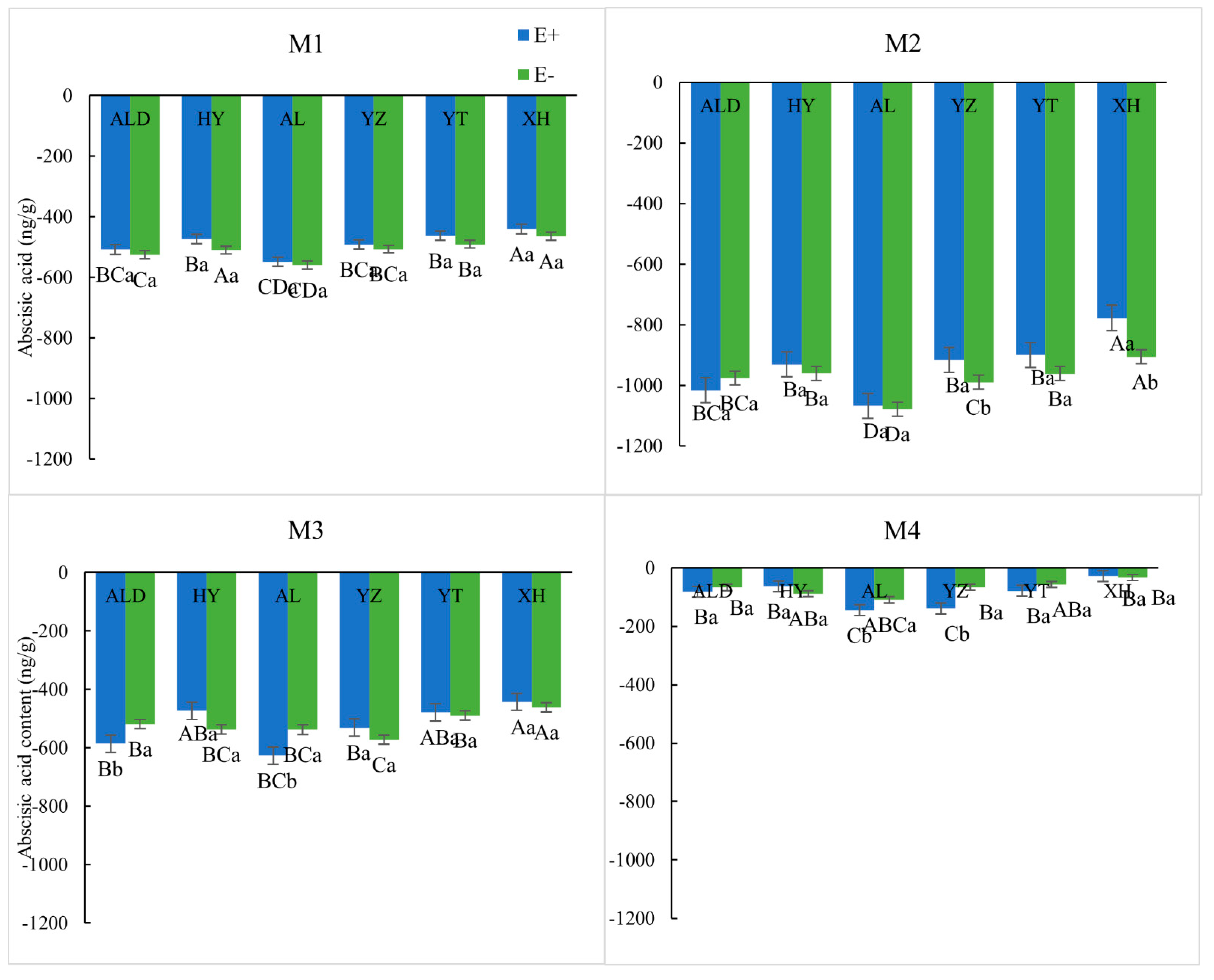
2.5. Determination of Internal Hormone Levels
2.6. Data Analysis
3. Results
3.1. Germination Rate
3.2. Physiological Elements
3.3. Interactive Effects
4. Discussion
4.1. Effects of Endophyte on Achnatherum inebrians Seed Germination
4.2. Effects of Geographic Origin on Seed Germination
4.3. Effects of Dormancy Breaking Treatments on Seed Germination
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 96, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, 874–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adkins, S.W.; Bellairs, S.M.; Loch, D.S. Seed dormancy mechanisms in warm season grass species. Euphytica 2020, 126, 13–20. [Google Scholar] [CrossRef]
- Abbasi, M.R. Breaking seed dormancy in some Iranian wild clovers (Trifolium spp.) selected as promising new germplasm. Genet. Res. Crop Evol. 2020, 67, 1577–1585. [Google Scholar] [CrossRef]
- Adkins, S.W.; Bellairs, S.M. Seed dormancy mechanisms in Australian native species. In Proceedings of the Third Native Seed Biology for Revegetation Workshop, Perth, Australia, 17–18 May 1995; Australian Centre for Minesite Rehabilitation Research: Kenmore, Australia, 2000. [Google Scholar]
- Baskin, J.M.; Schank, S.C.; West, S.H. Seed dormancy in 2 species of Digitaria from Africa. Crop Sci. 1969, 9, 584. [Google Scholar] [CrossRef]
- Keshtkar, A.; Keshtkar, H.; Razavi, S.; Dalfardi, S. Methods to break seed dormancy of Astragalus cyclophyllon. Afr. J. Biotechnol. 2010, 7, 3874–3877. [Google Scholar]
- Li, X.; Chen, L.; Gao, H.D. Endogenous screening of Torreya grandis seeds inhibiting their germination and growth. J. Southwest For. Univ. (Nat. Sci.) 2018, 38, 28–33. [Google Scholar]
- Kucera, B.; Cohn, M.A.; Leubner-Metzger, G. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 2005, 15, 218–307. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2013, 99, 110–121. [Google Scholar] [CrossRef]
- Fennimore, S.A.; Foley, M.E. Genetic and physiological evidence for the role of gibberellic acid in the germination of dormant Avena fatua seeds. J. Exp. Bot. 1998, 49, 89–94. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Z.H.; Hou, Q.Y.; Yu, F.Y. Investigation on dormancy characteristics of Styrax tonkinensis seeds. J. Cent. South Univ. For. Technol. 2019, 39, 45–51. [Google Scholar]
- Zhang, J.B. Variation of Storage Substances and Hormones during Cold Stratification for Seeds of Acer mono; Northeast Forestry University: Harbin, China, 2008. [Google Scholar]
- Zhao, Y.Y.; Song, C.C.; Brummell, D.A.; Qi, S.N.; Lin, Q.; Bi, J.F.; Duan, Y.Q. Salicylic acid treatment mitigates chilling injury in peach fruit by regulation of sucrose metabolism and soluble sugar content. Food Chem. 2021, 358, 129867. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Song, C.C.; Brummell, D.A.; Qi, S.N.; Lin, Q.; Bi, J.F.; Duan, Y.Q. Jasmonic acid treatment alleviates chilling injury in peach fruit by promoting sugar and ethylene metabolism. Food Chem. 2021, 338, 128005. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.S.; Song, M.L.; Wang, Y.Q.; Saikkonen, K.; Wang, H.S. Interactive effects of Epichloë fungal and host origins on the seed germination of Achnatherum inebrians. Symbiosis 2019, 79, 49–58. [Google Scholar] [CrossRef]
- Li, C.J.; Gao, J.H.; Nan, Z.B. Interactions of Neotyphodium gansuense, Achnatherum inebrians, and plant-pathogenic fungi. Mycol. Res. 2007, 111, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Li, C.J.; Nan, Z.B.; Matthew, C. Neotyphodium endophyte increases Achnatherum inebrians (drunken horse grass) resistance to herbivores and seed predators. Weed Res. 2012, 52, 70–78. [Google Scholar] [CrossRef]
- Ren, J.Z. Common poisonous plants in north-western grasslands of China. J. Gansu Agric. Univ. 1959, 1, 9–16. [Google Scholar]
- Bruehl, G.W.; Kaiser, W.J.; Klein, R.E. An endophyte of Achnatherum inebrians, an intoxicating grass of northwest China. Mycologia 1994, 86, 773–776. [Google Scholar] [CrossRef]
- Li, C.J.; Nan, Z.B.; Paul, V.H.; Dapprich, P.D.; Liu, Y. A new Neotyphodium species symbiotic with drunken horse grass (Achnatherum inebrians) in China. Mycotaxon 2005, 90, 141–147. [Google Scholar]
- Wang, J.F.; Hou, W.P.; Christensen, M.J.; Xia, C.; Chen, T.; Zhang, Z.X.; Nan, Z.B. The fungal endophyte Epichloe gansuensis increase NaCl-tolerance in Achnatherum inebrians through enhancing the activity of plasma membrane H+-ATPase and glucose-6-phosphate dehydrogenase. Sci. China Life Sci. 2021, 64, 452–465. [Google Scholar] [CrossRef]
- Moon, C.D.; Scott, B.; Schardl, C.L.; Christensen, M.J. The Evolutionary Origins of Epichloë Endophytes from Annual Ryegrasses. Mycologia 2000, 92, 1103–1118. [Google Scholar]
- Chen, N.; He, R.L.; Chai, Q.; Li, C.J.; Nan, Z.B. Transcriptomic analyses giving insights into molecular regulation mechanisms involved in cold tolerance by Epichloë endophyte in seed germination of Achnatherum inebrians. Plant Growth Regul. 2016, 80, 367–375. [Google Scholar] [CrossRef]
- Zhang, X.X.; Li, C.J.; Nan, Z.B. Effects of cadmium stress on growth and anti-oxidative systems in Achnatherum inebrians symbiotic with Neotyphodium gansuense. J. Hazard Mater. 2010, 175, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, 99–127. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Bacon, C.W.; Schardl, C.L.; White, J.F.; Tadych, J.M. Nomenclatural realignment of Neotyphodium species with genus Epichloë. Mycologia 2014, 106, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Simpson, W.R.; Faville, M.J.; Moraga, R.A.; Williams, W.M.; Mcmanus, M.T.; Johnson, R.D. Epichloë fungal endophytes and the formation of synthetic symbioses in Hordeeae (1⁄4 Triticeae) grasses. J. Syst. Evol. 2014, 52, 794–806. [Google Scholar] [CrossRef]
- Thom, E.R.; Popayt, A.J.; Waugh, C.D.; Minnee, E.M.K. Impact of novel endophytes in perennial ryegrass on herbage production and insect pests from pastures under dairy cow grazing in northern New Zealand. Grass Forage Sci. 2013, 69, 191–204. [Google Scholar] [CrossRef]
- Bacon, C.W. Abiotic stress tolerances (moisture, nutrients) and photosynthesis in endophyte-infected tall fescue. Agric. Ecosyst. Environ. 1993, 44, 123–142. [Google Scholar] [CrossRef]
- Yao, X. Effect of Achnatherum inebrians—Epichloe gansuensis Symbiont on Plant Composition and Soil Microbial Community Diversity in Rangelands; Lanzhou University: Lanzhou, China, 2019. [Google Scholar]
- Li, N.N.; Zhao, Y.F.; Xia, C.; Zhong, R.; Zhang, X.X. Effects of thiophanate methyl on seed borne Epichole fungal endophyte of Achnatherum inerbians. Pratacult. Sci. 2016, 10, 1306–1314. [Google Scholar]
- Bacon, C.W.; White, J.F. Microbial Endophytes; Marcel Dekker: New York, NY, USA, 1994; pp. 341–388. [Google Scholar]
- Carruggio, F.; Onofri, A.; Impelluso, C.; Giusso del Galdo, G.; Scopece, G.; Cristaudo, A. Seed Dormancy Breaking and Germination in Bituminaria basaltica and B. bituminosa (Fabaceae). Plants 2020, 9, 1110. [Google Scholar] [CrossRef]
- Grappin, P.; Bouinot, D.; Sotta, B.; Miginiac, E.; Jullien, M. Control of seed dormancy in Nicotiana plumbaginifolia: Post-imbibition abscisic acid synthesis imposes dormancy maintenance. Planta 2000, 210, 279–285. [Google Scholar] [CrossRef]
- Bano, C.; Amist, N.; Singh, N.B. UV-B radiation escalate allelopathic effect of benzoic acid on Solanum lycopersicum L. Sci. Hortic. 2017, 220, 199–205. [Google Scholar] [CrossRef]
- Chen, X.P.; Wang, X.F.; Sun, X.L.; Zhao, Z.Z. Determination of gibberellin abbeymycin and abscisic acid in cucumber by high-performance liquid chromatography. Shandong Agric. Sci. 2005, 226, 526–532. [Google Scholar]
- Fu, N.; Song, H.; Wang, S.J.; Xie, H.F.; Wei, M.H.; Xing, L.; Zhang, Y.; Liu, J.R. Research progress on seed dormancy and breaking methods. J. Anhui Agric. Sci. 2018, 46, 10–12. [Google Scholar]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.Z.; Li, C.J.; Swoboda, G.A.; Young, C.A.; Sugawara, K.; Leuchtmann, A.; Schardl, C.L. Two distinct Epichloë species symbiotic with Achnatherum inebrians, drunken horse grass. Mycologia 2015, 107, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Gundel, P.E.; Maseda, P.H.; Ghersa, C.M.; Benech-Arnolo, R. Effects of the Neotyphodium endophyte fungus on dormancy and germination rate of Lolium multiflorum seeds. Aust. Ecol. 2006, 31, 767–775. [Google Scholar] [CrossRef]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis. Fungal Ecol. 2017, 29, 52–58. [Google Scholar] [CrossRef]
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed dormancy and germination. Curr. Opin. Plant Biol. 2002, 5, 33–36. [Google Scholar] [CrossRef] [Green Version]
- White, C.N.; Rivin, C.J. Gibberellins and seed development in maize. II. Gibberellin synthesis inhibition enhances abscisic acid signaling in cultured embryos. Plant Physiol. 2000, 122, 1089–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soitamo, A.J.; Piippo, M.; Allahverdiyeva, Y.; Battchikova, N.; Aro, E.M. Light has a specific role in modulating Arabidopsis gene expression at low temperature. BMC Plant Biol. 2008, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethke, P.C.; Libourel, I.G.; Aoyama, N.; Chung, Y.Y.; Still, D.W.; Jones, R.L. The Arabidopsis aleurone layer responds to nitric oxide, gibberellin, and abscisic acid and is sufficient and necessary for seed dormancy. Plant Physiol. 2007, 1436, 1173–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, J.P.; Zhang, S.L. Changes in germination, storage materials and antioxidant enzyme activities in pear (Pyrus betulaefolia Bge. and Pyrus calleryana Dcne.) stock seeds during cold stratification. Seed Sci. Technol. 2011, 39, 655–659. [Google Scholar] [CrossRef]
- Valenzuela-Núez, L.M.; Briceo-Contreras, E.A.; Esparza-Rivera, J.R.; Pea, G.; Núez-Colima, J.A. Seasonal changes in soluble sugar concentration in pecan perennial organs (Carya Illinoiensis (Wangenh) Koch). Acta Univ. 2019, 29, 1–13. [Google Scholar]
- Wen, B.B.; Zhang, X.H.; Chen, X.D.; Gao, D.S.; Zhu, C.Y.; Xiao, W. Physiological changes of peach cultivars with three different chilling requirement during dormancy induction. J. Shandong Agric. Univ. (Nat. Sci. Ed.) 2018, 50, 1–5 + 74. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Longitude and Latitude | Elevation (m) | Mean Temperature (°C) | Mean Precipitation (mm) |
|---|---|---|---|---|
| Gansu-Yuzhong (YZ) | 35°56′26.7″ N | 1850 | 6.6 | 320 |
| 104°02′48.9″ E | ||||
| Gansu-Yingtai (YT) | 36°52′42.7″ N | 2279 | 5.3 | 331 |
| 103°47′19.8″ E | ||||
| Gansu-Xiahe (XH) | 35°32′16.4″ N | 3500 | 2.6 | 516 |
| 102°25′36.7″ E | ||||
| Ningxia-Haiyuan (HY) | 36°36′9″ N | 1732 | 7 | 286 |
| 105°39′46″ E | ||||
| Alxa League (AL) | 38°39′15″ N | 1830 | 8 | 113 |
| 105°44′21″ E | ||||
| Alxa Desert (ALD) | 38°56′16″ N | 1660 | 7.7 | 39 |
| 105°47′39″ E |
| Break Dormancy Method | Immersion Time | Temperature |
|---|---|---|
| Needle pin (M1) | / | 22 ± 2 °C |
| Water soaking (M2) | 30 min | 25/20 °C |
| 98% H2SO4 soaking (M3) | 20 min | 22 ± 2 °C |
| Exogenous GA soaking(M4) | 30 min | 22 ± 2 °C |
| Control (CK) | / | 22 ± 2 °C |
| Source of Variation | df | Germination Rate | Soluble Sugar | Gibberellin | Auxin | Abscisic Acid | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | ||
| Origin | 5 | 53.01 | <0.001 | 992.18 | <0.001 | 455.01 | <0.001 | 59.37 | <0.001 | 19.18 | <0.001 |
| Method | 4 | 438.88 | <0.001 | 193.03 | <0.001 | 1510.96 | <0.001 | 35,819.20 | <0.001 | 3909.35 | <0.001 |
| Endophyte | 1 | 563.25 | <0.001 | 62.07 | <0.001 | 2.63 | <0.05 | 1.55 | <0.05 | 10.42 | <0.001 |
| Origin*Method | 20 | 4.38 | <0.001 | 3.13 | <0.001 | 71.71 | <0.001 | 1.67 | 0.22 | 2.51 | <0.001 |
| Origin*Endophyte | 5 | 2.09 | 0.07 | 16.27 | <0.001 | 0.99 | 0.42 | 1.11 | 0.36 | 3.95 | <0.001 |
| Method*Endophyte | 4 | 10.68 | <0.001 | 1.40 | 0.24 | 4.72 | <0.001 | 0.63 | 0.65 | 2.06 | 0.09 |
| Origin*Method*Endophyte | 20 | 2.24 | <0.001 | 0.77 | 0.74 | 1.70 | <0.05 | 1.68 | 0.05 | 1.52 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Su, K.; Li, C.; White, J.F. Interactive Effects of Epichloë Endophyte, Dormancy-Breaking Treatments and Geographic Origin on Seed Germination of Achnatherum inebrians. Microorganisms 2021, 9, 2183. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112183
Chen Y, Su K, Li C, White JF. Interactive Effects of Epichloë Endophyte, Dormancy-Breaking Treatments and Geographic Origin on Seed Germination of Achnatherum inebrians. Microorganisms. 2021; 9(11):2183. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112183
Chicago/Turabian StyleChen, Yaqi, Kaiqi Su, Chunjie Li, and James F. White. 2021. "Interactive Effects of Epichloë Endophyte, Dormancy-Breaking Treatments and Geographic Origin on Seed Germination of Achnatherum inebrians" Microorganisms 9, no. 11: 2183. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112183