
The Impact of Temperature on the Sporogonic Development of the Tropical Avian Malaria Parasite Plasmodium relictum (Genetic Lineage pGRW4) in Culex pipiens Form molestus Mosquitoes

{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Site and Design of the Experiment
2.3. Experimental Infection of Vectors and Dissection of Female Mosquitoes
2.4. Molecular Analysis
2.5. Statistical Analysis
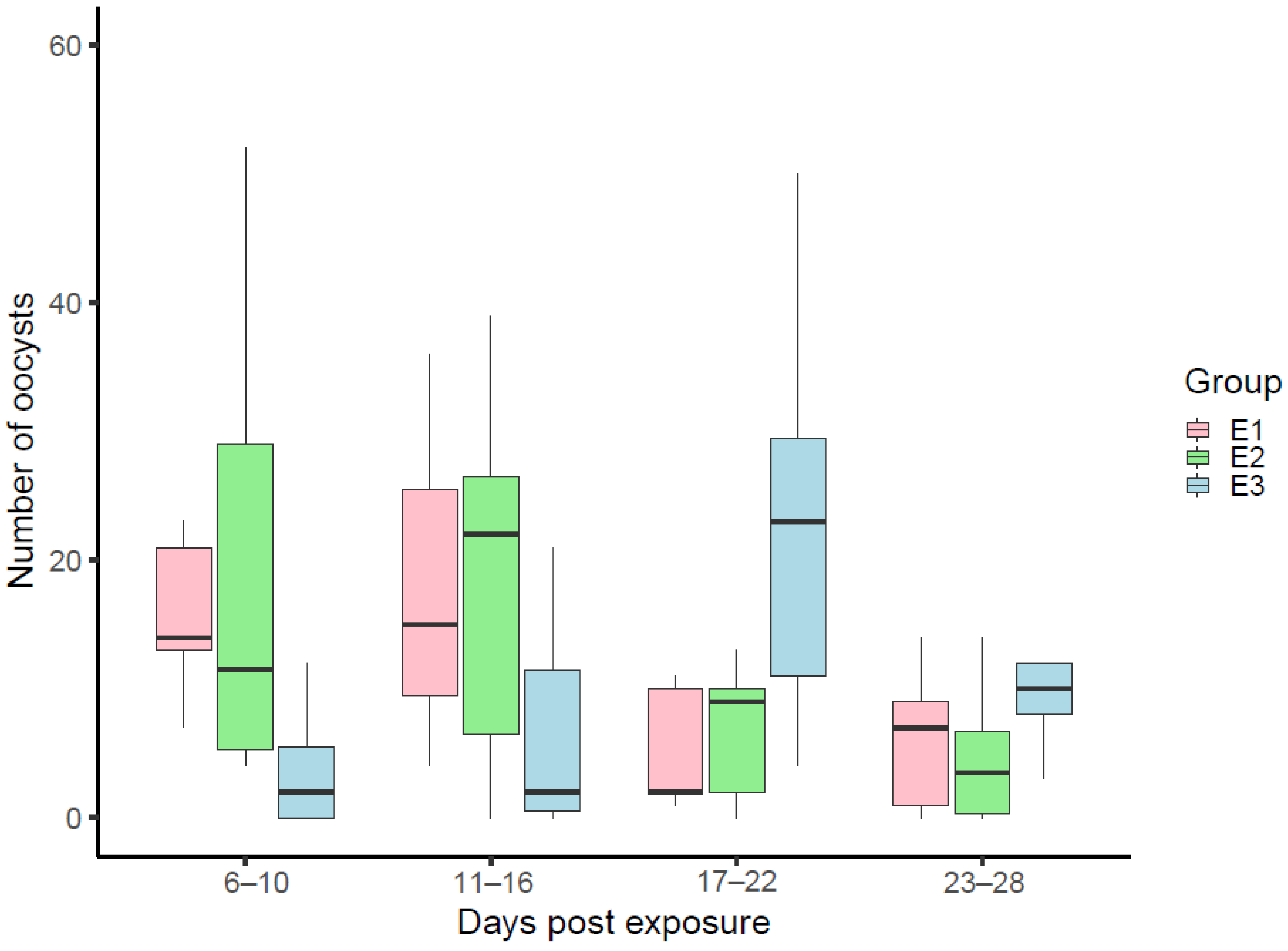
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- May, R.M.; Anderson, R.M. Parasite–host coevolution. Parasitology 1990, 100, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Leggett, H.C.; Buckling, A.; Long, G.H.; Boots, M. Generalism and the evolution of parasite virulence. Trends Ecol. Evol. 2009, 28, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Buczek, A.M.; Buczek, W.; Buczek, A.; Bartosik, K. The potential role of migratory birds in the rapid spread of ticks and tick-borne pathogens in the changing climatic and environmental conditions in Europe. Int. J. Environ. Res. Public Health 2020, 17, 2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, R.E. The role of introduced diseases in the extinction of the endemic Hawaiian avifauna. Condor 1968, 70, 101–120. [Google Scholar] [CrossRef]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2004; ISBN 978-0415300971. [Google Scholar] [CrossRef]
- Purse, B.V.; Mellor, P.S.; Rogers, D.J.; Samuel, A.R.; Mertens, P.P.; Baylis, M. Climate change and the recent emergence of bluetongue in Europe. Nat. Rev. Microbiol. 2005, 3, 171–181. [Google Scholar] [CrossRef]
- Dunn, A.M.; Hatcher, M.J. Parasites and biological invasions: Parallels, interactions, and control. Trends Parasitol. 2015, 35, 189–199. [Google Scholar] [CrossRef]
- Atkinson, C.T.; Thomas, N.J.; Hunter, D.B. Parasitic Diseases of Wild Birds; Wiley Blackwell: Hoboken, NJ, USA, 2008; ISBN 978-0-813-82081-1. [Google Scholar]
- Garnham, P.C.C. Malaria Parasites and Other Haemosproridia; Blackwell Scientific Publications: Oxford, UK, 1966. [Google Scholar]
- Beadell, J.S.; Gering, E.; Austin, J.; Dumbacher, J.P.; Peirce, M.A.; Pratt, T.K.; Atkinson, C.T.; Fleischer, R.C. Prevalence and differential host-specificity of two avian blood parasite genera in the Australo-Papuan region. Mol. Ecol. 2004, 13, 3829–3844. [Google Scholar] [CrossRef]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Van Riper, C.; Van Riper, S.G.; Goff, M.L.; Laird, M. The epizootiology and ecological significance of malaria in Hawaiian land birds. Ecol. Monogr. 1986, 56, 327–344. [Google Scholar] [CrossRef]
- Bennett, G.F.; Peirce, M.A.; Ashford, R.W. Avian haematozoa: Mortality and pathogenicity. J. Nat. Hist. 1993, 27, 993–1001. [Google Scholar] [CrossRef]
- Atkinson, C.T.; LaPointe, D.A. Introduced avian diseases, climate change, and the future of Hawaiian honeycreepers. J. Avian Med. Surg. 2009, 23, 53–63. [Google Scholar] [CrossRef]
- Verwey, J.K.; Peters, A.; Monks, D.; Raidal, S.R. Spillover of avian haemosporidian parasites (Haemosporidia: Plasmodium) and death of captive psittacine species. Aust. Vet. J. 2018, 96, 93–97. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; Brunton, D.; Stidworthy, M.F.; Elsheikha, H.M.; Pennycott, T.; Schulze, C.; Braun, M.; Wink, M.; Gerlach, H.; Pendl, H.; et al. Haemoproteus minutus is highly virulent for Australasian and South American parrots. Parasit. Vectors 2019, 12, 40. [Google Scholar] [CrossRef]
- Huijben, S.; Schaftenaar, W.; Wijsman, A.; Paaijmans, K.; Takken, W. Avian Malaria in Europe: An Emerging Infectious Disease. In Emerging Pests and Vector-borne Diseases in Europe, 1st ed.; Takken, W., Knols, B.G., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; Volume 1, pp. 59–74. [Google Scholar]
- Hellgren, O.; Waldenström, J.; Peréz-Tris, J.; Szöll, E.; Si, Ö.; Hasselquist, D.; Krizanauskiene, A.; Ottosson, U.; Bensch, S. Detecting shifts of transmission areas in avian blood parasites—A phylogenetic approach. Mol. Ecol. 2007, 16, 1281–1290. [Google Scholar] [CrossRef]
- Marzal, A.; Ricklefs, R.E.; Valkiūnas, G.; Albayrak, T.; Arriero, E.; Bonneaud, C.; Czirják, G.A.; Ewen, J.; Hellgren, O.; Hořáková, D.; et al. Diversity, loss, and gain of malaria parasites in a globally invasive bird. PLoS ONE 2011, 6, e21905. [Google Scholar] [CrossRef] [Green Version]
- Waldenström, J.; Bensch, S.; Kiboi, S.; Hasselquist, D.; Ottosson, U. Cross-species infection of blood parasites between resident and migratory songbirds in Africa. Mol. Ecol. 2002, 11, 1545–1554. [Google Scholar] [CrossRef] [Green Version]
- Bensch, S.; Waldenström, J.; Jonzen, N.; Westerdahl, H.; Hansson, B.; Sejberg, D.; Hasselquist, D. Temporal dynamics and diversity of avian malaria parasites in a single host species. J. Anim. Ecol. 2007, 76, 112–122. [Google Scholar] [CrossRef]
- Dimitrov, D.; Zehtindjiev, P.; Bensch, S. Genetic diversity of avian blood parasites in SE Europe: Cytochrome b lineages of the genera Plasmodium and Haemoproteus (Haemosporida) from Bulgaria. Acta Parasitol. 2010, 55, 201–209. [Google Scholar] [CrossRef]
- Zehtindjiev, P.; Ilieva, M.; Westerdahl, H.; Hansson, B.; Valkiūnas, G.; Bensch, S. Dynamics of parasitemia of malaria parasites in a naturally and experimentally infected migratory songbird, the great reed warbler Acrocephalus arundinaceus. Exp. Parasitol. 2008, 119, 99–110. [Google Scholar] [CrossRef]
- Palinauskas, V.; Žiegytė, R.; Ilgūnas, M.; Iezhova, T.A.; Bernotienė, R.; Bolshakov, C.; Valkiūnas, G. Description of the first cryptic avian malaria parasite, Plasmodium homocircumflexum n. sp., with experimental data on its virulence and development in avian hosts and mosquitoes. Int. J. Parasitol. 2015, 45, 51–62. [Google Scholar] [CrossRef]
- Ilgūnas, M.; Palinauskas, V.; Platonova, E.; Iezhova, T.; Valkiūnas, G. The experimental study on susceptibility of common European songbirds to Plasmodium elongatum (lineage pGRW6), a widespread avian malaria parasite. Malar. J. 2019, 18, 290. [Google Scholar] [CrossRef] [PubMed]
- Platonova, E.; Aželytė, J.; Iezhova, T.; Ilgūnas, M.; Mukhin, A.; Palinauskas, V. Experimental study of newly described avian malaria parasite Plasmodium (Novyella) collidatum n. sp., genetic lineage pFANTAIL01 obtained from South Asian migrant bird. Malar. J. 2021, 20, 82. [Google Scholar] [CrossRef] [PubMed]
- Žiegytė, R.; Platonova, E.; Bernotienė, R.; Valkiūnas, G.; Palinauskas, V. Complete sporogony of the blood parasite Haemoproteus nucleocondensus in common biting midges: Why is its transmission interrupted in Europe? Parasitology 2020, 147, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Žiegytė, R.; Palinauskas, V.; Bernotienė, R.; Bukauskaitė, D.; Ilgūnas, M.; Dimitrov, D.; Iezhova, T.A. Complete sporogony of Plasmodium relictum (lineage pGRW4) in mosquitoes Culex pipiens pipiens, with implications on avian malaria epidemiology. Parasitol. Res. 2015, 114, 3075–3085. [Google Scholar] [CrossRef]
- Dimitrov, D.; Palinauskas, V.; Iezhova, T.A.; Bernotienė, R.; Ilgūnas, M.; Bukauskaitė, D.; Zehtindjiev, P.; Ilieva, M.; Shapoval, A.P.; Bolshakov, C.V.; et al. Plasmodium spp.: An experimental study on vertebrate host susceptibility to avian malaria. Exp. Parasitol. 2015, 148, 1–16. [Google Scholar] [CrossRef]
- Chagas, C.R.F.; Harl, J.; Valkiūnas, G. Co-infections of Plasmodium relictum lineages pSGS1 and pGRW04 are readily distinguishable by broadly used PCR-based protocols, with remarks on global distribution of these malaria parasites. Acta Trop. 2021, 217, 105860. [Google Scholar] [CrossRef]
- Atkinson, C.T.; Utzurrum, R.B.; LaPointe, D.A.; Camp, R.J.; Crampton, L.H.; Foster, J.T.; Giambelluca, T.W. Changing climate and the altitudinal range of avian malaria in the Hawaiian Islands—An ongoing conservation crisis on the island of Kaua’i. Glob. Chang. Biol. 2014, 20, 2426–2436. [Google Scholar] [CrossRef]
- Soares, L.; Marra, P.; Gray, L.; Ricklefs, R.E. The malaria parasite Plasmodium relictum in the endemic avifauna of eastern Cuba. Conserv. Biol. 2017, 31, 1477–1482. [Google Scholar] [CrossRef]
- Westerdahl, H.; Waldenström, J.; Hansson, B.; Hasselquist, D.; von Schantz, T.; Bensch, S. Associations between malaria and MHC genes in a migratory songbird. Proc. R. Soc. B. 2005, 272, 1511–1518. [Google Scholar] [CrossRef]
- Asghar, M.; Hasselquist, D.; Bensch, S. Are chronic avian haemosporidian infections costly in wild birds? J. Avian Biol. 2011, 42, 530–537. [Google Scholar] [CrossRef]
- Lemke, H.W.; Tarka, M.; Klaassen, R.H.; Åkesson, M.; Bensch, S.; Hasselquist, D.; Hansson, B. Annual cycle and migration strategies of a trans-Saharan migratory songbird: A geolocator study in the great reed warbler. PLoS ONE 2013, 8, e79209. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Tsuda, Y. Avian Plasmodium lineages found in spot surveys of mosquitoes from 2007 to 2010 at Sakata wetland, Japan: Do dominant lineages persist for multiple years? Mol. Ecol. 2012, 21, 5374–5385. [Google Scholar] [CrossRef]
- Vinogradova, E.B. Mosquitoes Culex Pipiens Pipiens: Taxonomy, Distribution, Ecology, Physiology, Genetics and Control; PenSoft: Sofia, Bulgaria, 2004. [Google Scholar]
- Vanderberg, J.P.; Yoeli, M. Effects of temperature on sporogonic development of Plasmodium berghei. J. Parasitol. 1966, 52, 559–564. [Google Scholar] [CrossRef]
- Fortini, L.B.; Kaiser, L.R.; LaPointe, D.A. Fostering real-time climate adaptation: Analyzing past, current, and forecast temperature to understand the dynamic risk to Hawaiian honeycreepers from avian malaria. Glob. Ecol. Conserv. 2020, 23, e01069. [Google Scholar] [CrossRef]
- Beier, J.C. Malaria parasite development in mosquitoes. Annu. Rev. Entomol. 1998, 43, 519–543. [Google Scholar] [CrossRef]
- LaPointe, D.A.; Goff, M.L.; Atkinson, C.T. Thermal constraints to the sporogonic development and altitudinal distribution of avian malaria in Hawai’i. J. Parasitol. 2010, 96, 318–324. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Križanauskienė, A.; Palinauskas, V.; Sehgal, R.N.M.; Bensch, S. A comparative analysis of microscopy and PCR-based detection methods for blood parasites. J. Parasitol. 2008, 94, 1395–1401. [Google Scholar] [CrossRef]
- Palinauskas, V.; Valkiūnas, G.; Bolshakov, C.V.; Bensch, S. Plasmodium relictum (lineage P-SGS1): Effects on experimentally infected passerine birds. Exp. Parasitol. 2008, 120, 372–380. [Google Scholar] [CrossRef]
- Žiegytė, R.; Bernotienė, R.; Bukauskaitė, D.; Palinauskas, V.; Iezhova, T.; Valkiūnas, G. Complete sporogony of Plasmodium relictum (lineages pSGS1 and pGRW11) in mosquito Culex pipiens pipiens form molestus, with implications to avian malaria epidemiology. J. Parasitol. 2014, 100, 878–882. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Mamiatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; ISBN 0879693096. [Google Scholar]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef]
- Hall, A. BioEdit: A user-friendly biological sequence alignment editor and analysis program of Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 April 2020).
- Ball, G.H.; Chao, J. Temperature stresses on the mosquito phase of Plasmodium relictum. J. Parasitol. 1964, 50, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Ball, G.H. The effect of low temperature on Plasmodium relictum in Culex tarsalis. J. Parasitol. 1962, 48, 252–254. [Google Scholar] [CrossRef] [PubMed]
- Ragab, H.A. Effect of humidities and temperatures on the size and number of oocysts of Plasmodium gallinaceum transmitted by a mosquito. Nature 1949, 163, 643–644. [Google Scholar] [CrossRef]
- Hunninen, A.V. Comparative development of Plasmodium relictum oocysts in Anopheles quadrimaculatus, A. albimanus, and Culex pipiens. J. Parasitol. 1953, 39, 28–32. [Google Scholar] [CrossRef]
- Huff, C.G. Further infectivity experiments with mosquitoes and bird malaria. Am. J. Trop. Med. Hyg. 1932, 15, 751–754. [Google Scholar] [CrossRef]
- Huff, C.G. Quantitative studies on size, variability, and growth rates of oocysts of different strains of avian malaria. Am. J. Trop. Med. Hyg. 1940, 32, 71–80. [Google Scholar]
- Rutledge, L.C.; Wand, R.A.; Buckwalter, R.M. Plasmodium spp; Dispersion of malarial oocysts populations in Anopheline and Culicine mosquitos. Exp. Parasitol. 1973, 34, 132–147. [Google Scholar] [CrossRef]
- Sergent, E.T. Influence du froid sur le developpement du Plasmodium relictum chez le moustique. Bull. Soc. Path. Exot. 1919, 12, 174–176. [Google Scholar]
- Goritzkaya, V. Some observations on P. praecox in experiments with artificial infection of C. pipiens. Parazitologicheski Sbornik 1934, 4, 85–92. [Google Scholar]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Fragner, K.; Weissenböck, H.; Atkinson, C.T.; Iezhova, T. Characterization of Plasmodium relictum, a cosmopolitan agent of avian malaria. Malar. J. 2018, 17, 184. [Google Scholar] [CrossRef] [Green Version]
- Martínez-de la Puente, J.; Santiago-Alarcon, D.; Palinauskas, V.; Bensch, S. Plasmodium relictum. Trends Parasitol. 2021, 37, 355–356. [Google Scholar] [CrossRef]
- Hellgren, O.; Atkinson, C.T.; Bensch, S.; Albayrak, T.; Dimitrov, D.; Ewen, J.G.; Kim, K.S.; Lima, M.R.; Martin, L.; Palinauskas, V.; et al. Global phylogeography of the avian malaria pathogen Plasmodium relictum based on MSP1 allelic diversity. Ecography 2015, 38, 842–850. [Google Scholar] [CrossRef]
- Beadel, J.S.; Ishitag, F.; Covas, R.; Melo, M.; Warnen, B.H.; Atkinson, C.T.; Bensch, S.; Graves, G.R.; Ihala, Y.V.; Peirce, M.A.; et al. Global phylogeographic limits of Hawaii’s avian malaria. Proc. Royal Soc. B Biol. Sci. 2015, 273, 2935–2944. [Google Scholar] [CrossRef] [Green Version]
- Van Riper III, C. The impact of introduced vectors and avian malaria on insular passeriform bird population in Hawaii. Bull. Soc. Vector Ecol. 1991, 16, 59–83. [Google Scholar]
- Dunn, J.C.; Stockdale, J.E.; Bradford, E.; Mccubbin, A.; Morris, A.J.; Grice, P.V.; Goodman, S.J.; Hamer, K.C. High rates of infection by blood parasites during the nestling phase in UK Columbids with notes on ecological associations. Parasitology 2017, 145, 622–628. [Google Scholar] [CrossRef] [Green Version]
- Zehtindjiev, P.; Križanauskienė, A.; Scebba, S.; Dimitrov, D.; Valkiūnas, G.; Hegemann, A.; Tieleman, B.I.; Bensch, S. Haemosporidian infections in skylarks (Alauda arvensis): A comparative PCR-based and microscopy study on the parasite diversity and prevalence in southern Italy and the Netherlands. Eur. J. Wildl. Res. 2011, 58, 335–344. [Google Scholar] [CrossRef]
- Calero-Riestra, M.; García, J.T. Sex-dependent differences in avian malaria prevalence and consequences of infections on nestling growth and adult condition in the Tawny pipit, Anthus campestris. Malar. J. 2016, 15, 178. [Google Scholar] [CrossRef] [Green Version]
- Miholcsa, T.; Tóth, A.; Csörgő, T. Change of timing of autumn migration in Acrocephalus and Locustella genus. Acta Zool. Hung. 2009, 55, 175–185. [Google Scholar]
- Koskimies, P.; Saurola, P. Autumn migration strategy of the sedge warbler Acrocephalus schoenobaenus in Finland: A preliminary report. Oris. Fenn. 1985, 62, 145–152. [Google Scholar]
- Ferraguti, M.; Martínez-de la Puente, J.; Muñoz, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Avian Plasmodium in Culex and Ochlerotatus mosquitoes from Southern Spain: Effects of season and host-feeding source on parasite dynamics. PLoS ONE 2013, 8, e66237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalubin, F.; Delédevant, A.; Glaizot, O.; Christe, P. Temporal changes in mosquito abundance (Culex pipiens), avian malaria prevalence and lineage composition. Parasit. Vectors 2013, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernotienė, R. The fauna and seasonal activity of mosquitoes (Diptera: Culicidae) in the Curonian Spit (Russia, Lithuania). Euro. Mosq. Bull. 2012, 30, 72–78. [Google Scholar]
- Ferrer, E.S.; García-Navas, V.; Sanz, J.J.; Ortego, J. Molecular characterization of avian malaria parasites in three Mediterranean blue tit (Cyanistes caeruleus) populations. Parasitol. Res. 2012, 111, 2137–2142. [Google Scholar] [CrossRef]
- Ferraguti, M.; Martínez-de la Puente, J.; Ruiz, S.; Soriguer., R.; Figuerola, J. On the study of the transmission networks of blood parasites from SW Spain: Diversity of avian haemosporidians in the biting midge Culicoides circumscriptus and wild birds. Parasit. Vectors 2013, 6, 208. [Google Scholar] [CrossRef] [Green Version]
- Videvall, E.; Palinauskas, V.; Valkiūnas, G.; Hellgren, O. Host transcriptional responses to high- and low-virulent avian malaria parasites. Am. Nat. 2020, 195, 1070–1084. [Google Scholar] [CrossRef]
- Aželytė, J.; Platonova, E.; Bensch, S.; Hellgren, O.; Palinauskas, V. A comparative analysis of the dynamics of Plasmodium relictum (GRW4) development in the blood during single and mixed infections. Acta Trop. 2021. submit. [Google Scholar]
- Palinauskas, V.; Žiegytė, R.; Šengaut, J.; Bernotienė, R. Different paths—the same virulence: Experimental study on avian single and co-infections with Plasmodium relictum and Plasmodium elongatum. Int. J. Parasitol. 2018, 48, 1089–1096. [Google Scholar] [CrossRef]
- Ferguson, H.M.; Read, A.F. Why is the effect of malaria parasites on mosquito survival still unresolved? Trends Parasitol. 2002, 18, 256–261. [Google Scholar] [CrossRef]
- Martínez-De la Puente, J.; Gutierrez-López, R.; Figuerola, J. Do avian malaria parasites reduce vector longevity? Curr. Opin. Insect Sci. 2018, 28, 113–117. [Google Scholar] [CrossRef]
- Huff, C.G.; Marchbank, D.F.; Shiroishi, T. Changes in infectiousness of malarial gametocytes. II. Analysis of the possible causative factors. Exp. Parasitol. 1958, 7, 399–417. [Google Scholar] [CrossRef]
- Da, D.F.; Churcher, T.S.; Yerbanga, R.S.; Yaméogo, B.; Sangaré, I.; Ouedraogo, J.B.; Cohuet, A. Experimental study of the relationship between Plasmodium gametocyte density and infection success in mosquitoes; implications for the evaluation of malaria transmission-reducing interventions. Exp. Parasitol. 2015, 149, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Dawes, E.J.; Zhuang, S.; Sinden, R.E.; Basáñez, M.G. The temporal dynamics of Plasmodium density through the sporogonic cycle within Anopheles mosquitoes. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 1197–1198. [Google Scholar] [CrossRef]
- Sinden, R.E.; Dawes, E.J.; Alavi, Y.; Waldock, J.; Finney, O.; Mendoza, J.; Butcher, G.A.; Andrews, L.; Hill, A.V.; Gilbert, S.C.; et al. Progression of Plasmodium berghei through Anopheles stephensi is density-dependent. PLoS Pathog. 2007, 3, e195. [Google Scholar] [CrossRef] [Green Version]
- Schneider, P.; Rund, S.S.C.; Smith, N.L.; Prior, K.F.; O’Donnell, A.J.; Reece, S.E. Adaptive periodicity in the infectivity of malaria gametocytes to mosquitoes. Proc Soc Biol. 2018, 285, 20181876. [Google Scholar] [CrossRef] [Green Version]
- Pigeault, R.; Vézilier, J.; Cornet, S.; Zélé, F.; Nicot, A.; Perret, P.; Gandon, S.; Rivero, A. Avian malaria: A new lease of life for an old experimental model to study the evolutionary ecology of Plasmodium. Phil. Trans. R. Soc. B 2015, 370, 20140300. [Google Scholar] [CrossRef] [Green Version]
- Cornet, S.; Nicot, A.; Rivero, A.; Gandon, S. Evolution of plastic transmission strategies in avian malaria. PLoS Pathog. 2014, 10, e1004308. [Google Scholar] [CrossRef] [Green Version]
- Lensen, A.; Bril, A.; van de Vegte, M.; van Gemert, G.J.; Eling, W.; Sauerwein, R. Plasmodium falciparum: Infectivity of cultured, synchronized gametocytes to mosquitoes. Exp. Parasitol. 1999, 91, 101–103. [Google Scholar] [CrossRef]
- Paul, R.E.; Brey, P.T.; Robert, V. Plasmodium sex determination and transmission to mosquitoes. Trends Parasitol. 2002, 18, 32–38. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Platonova, E.; Palinauskas, V. The Impact of Temperature on the Sporogonic Development of the Tropical Avian Malaria Parasite Plasmodium relictum (Genetic Lineage pGRW4) in Culex pipiens Form molestus Mosquitoes. Microorganisms 2021, 9, 2240. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112240
Platonova E, Palinauskas V. The Impact of Temperature on the Sporogonic Development of the Tropical Avian Malaria Parasite Plasmodium relictum (Genetic Lineage pGRW4) in Culex pipiens Form molestus Mosquitoes. Microorganisms. 2021; 9(11):2240. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112240
Chicago/Turabian StylePlatonova, Elena, and Vaidas Palinauskas. 2021. "The Impact of Temperature on the Sporogonic Development of the Tropical Avian Malaria Parasite Plasmodium relictum (Genetic Lineage pGRW4) in Culex pipiens Form molestus Mosquitoes" Microorganisms 9, no. 11: 2240. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112240