
The Exopolysaccharide Produced by Lactobacillus paracasei IJH-SONE68 Prevents and Ameliorates Inflammatory Responses in DSS–Induced Ulcerative Colitis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Purification of EPSs
2.3. IL-8 Release Inhibition Assay
2.4. Animals and Rearing Conditions
2.5. Preparation and Evaluation of the DSS-Induced UC Model in Mice
2.6. Myeloperoxidase (MPO) Activity Assay
2.7. Preparation, Manipulation, and qRT-PCR Analysis of RNA
2.8. Statistical Analyses
2.9. Histopathological Analysis
3. Results
3.1. Effect of the IJH-SONE68-Derived EPS on the Release of IL-8 from Caco-2 Cells
3.2. Preventive Effect of the IJH-SONE68-Derived EPS on DSS-Induced UC Model Mice
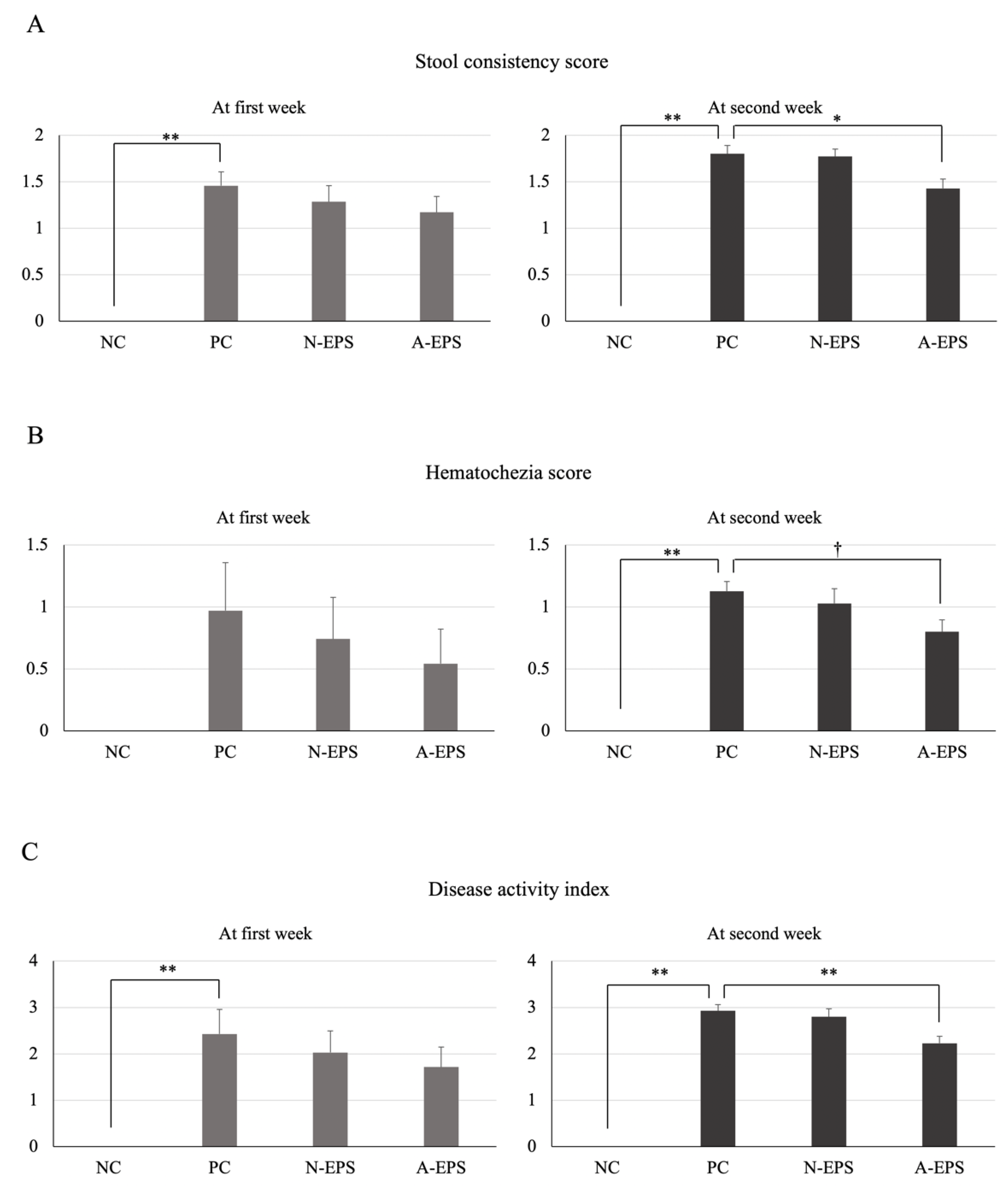
3.2.1. Disease Activity Index
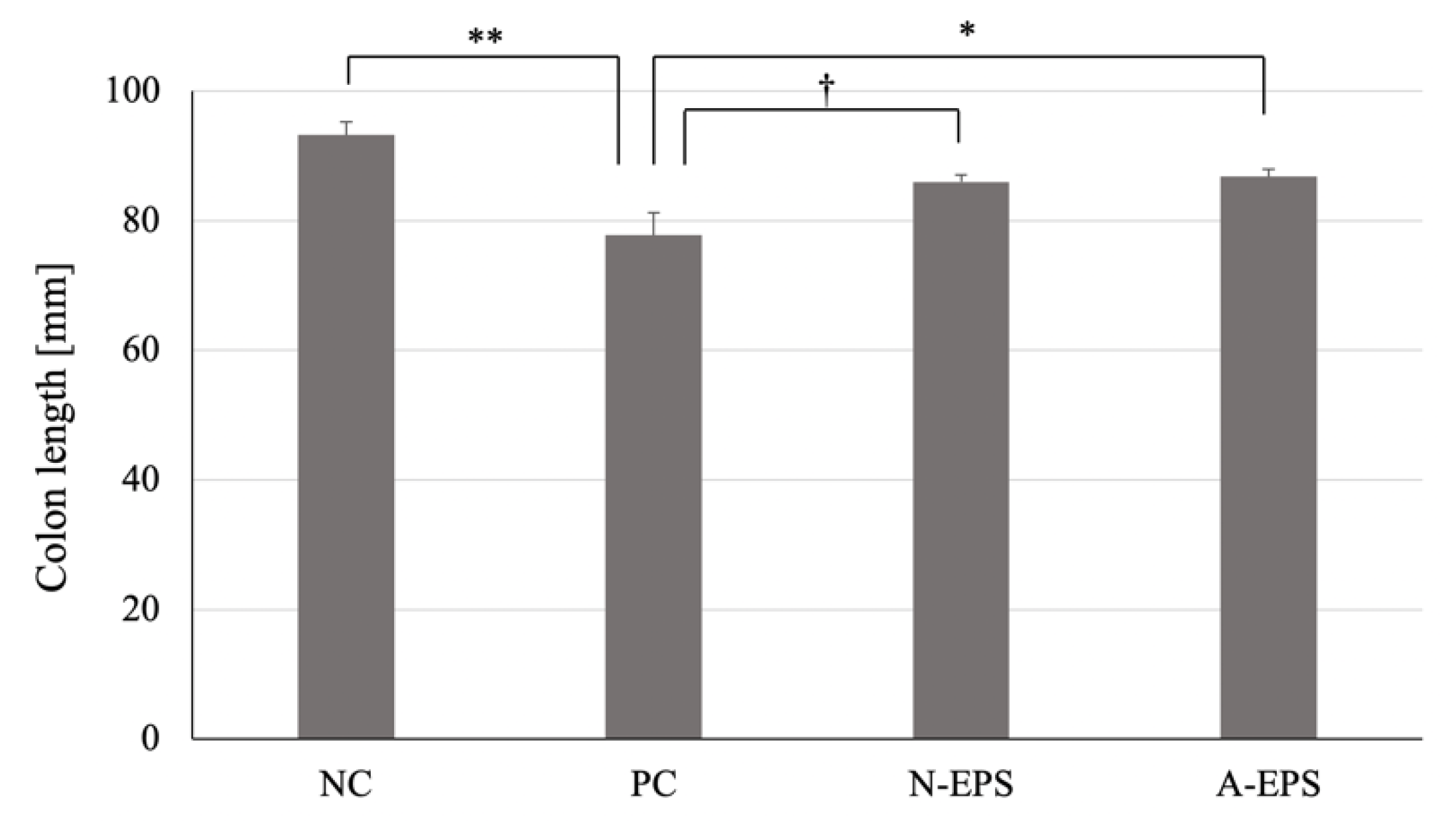
3.2.2. Colon Length
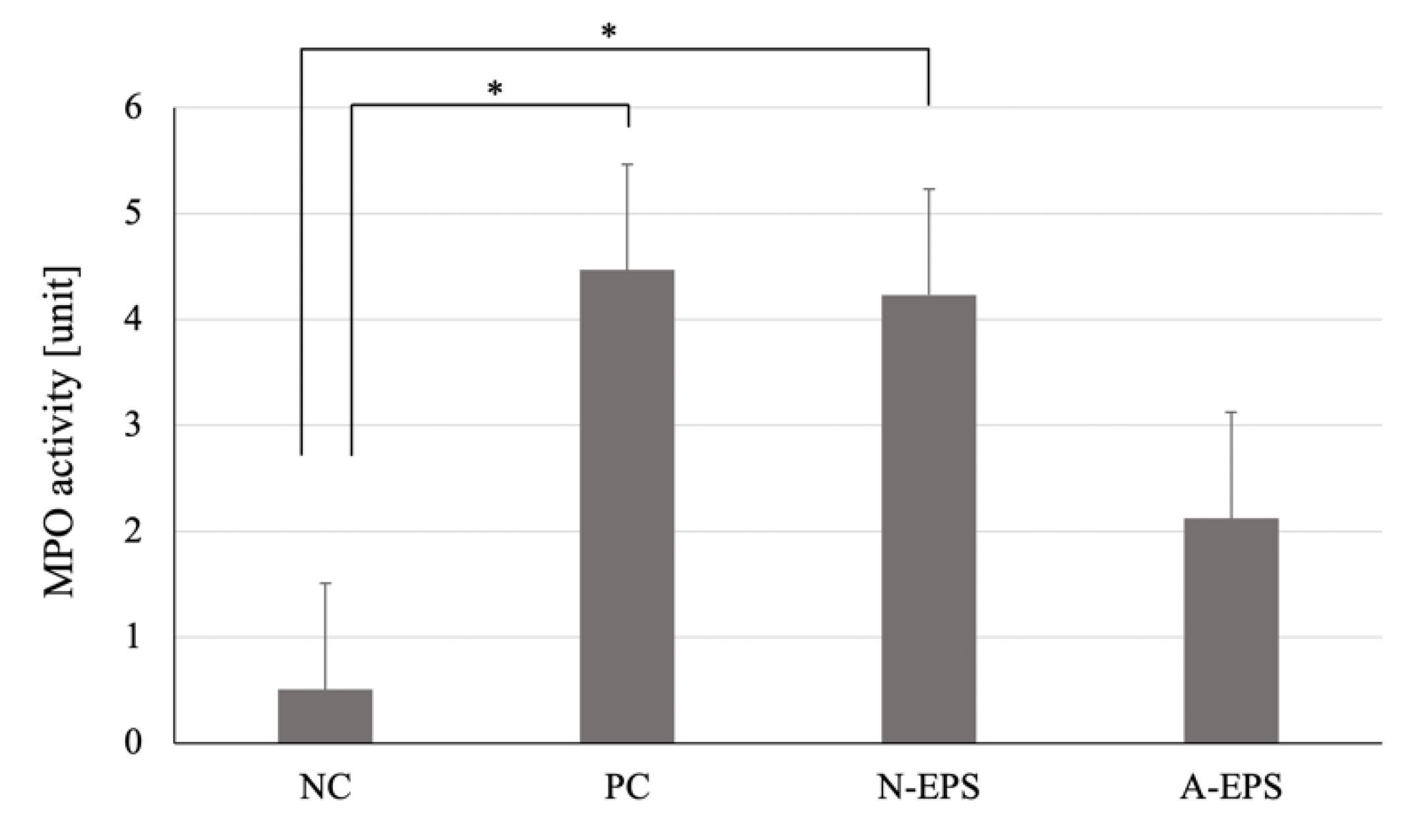
3.2.3. MPO Activity
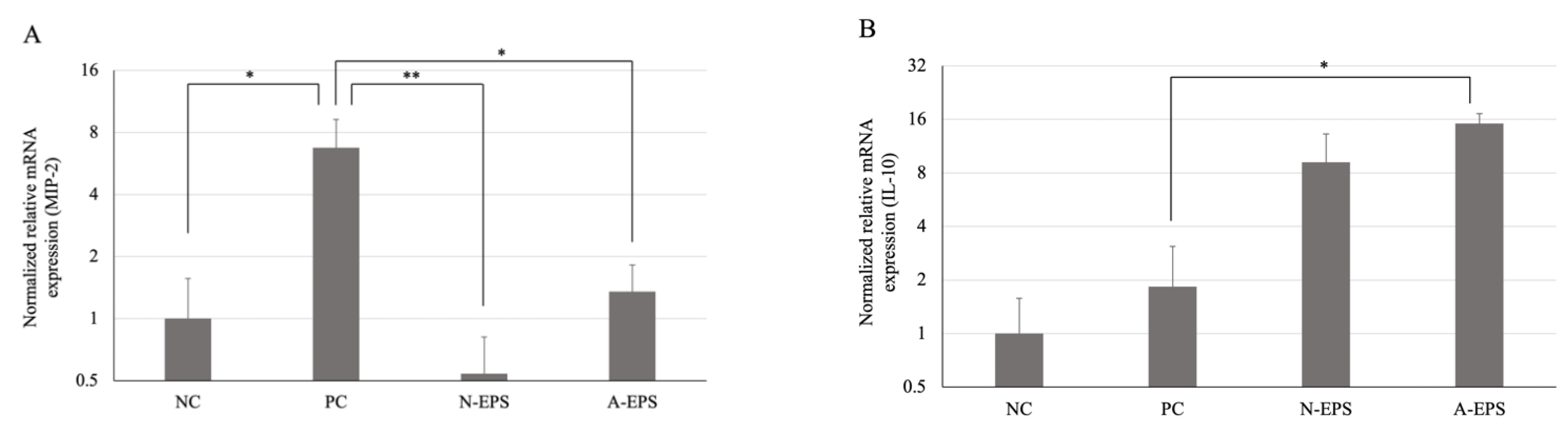
3.3. Difference in the Expression Level of Inflammatory-Related Cytokines
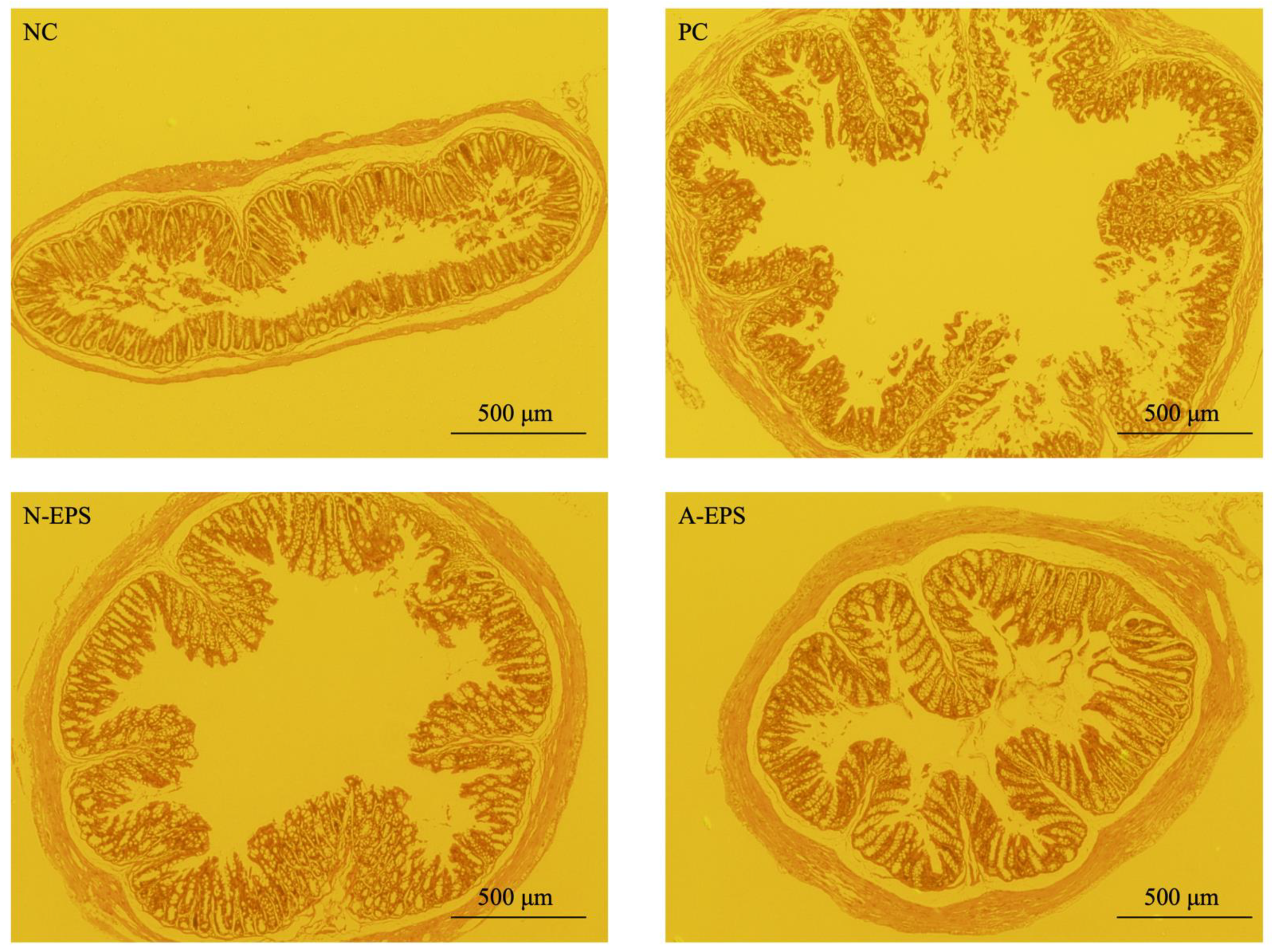
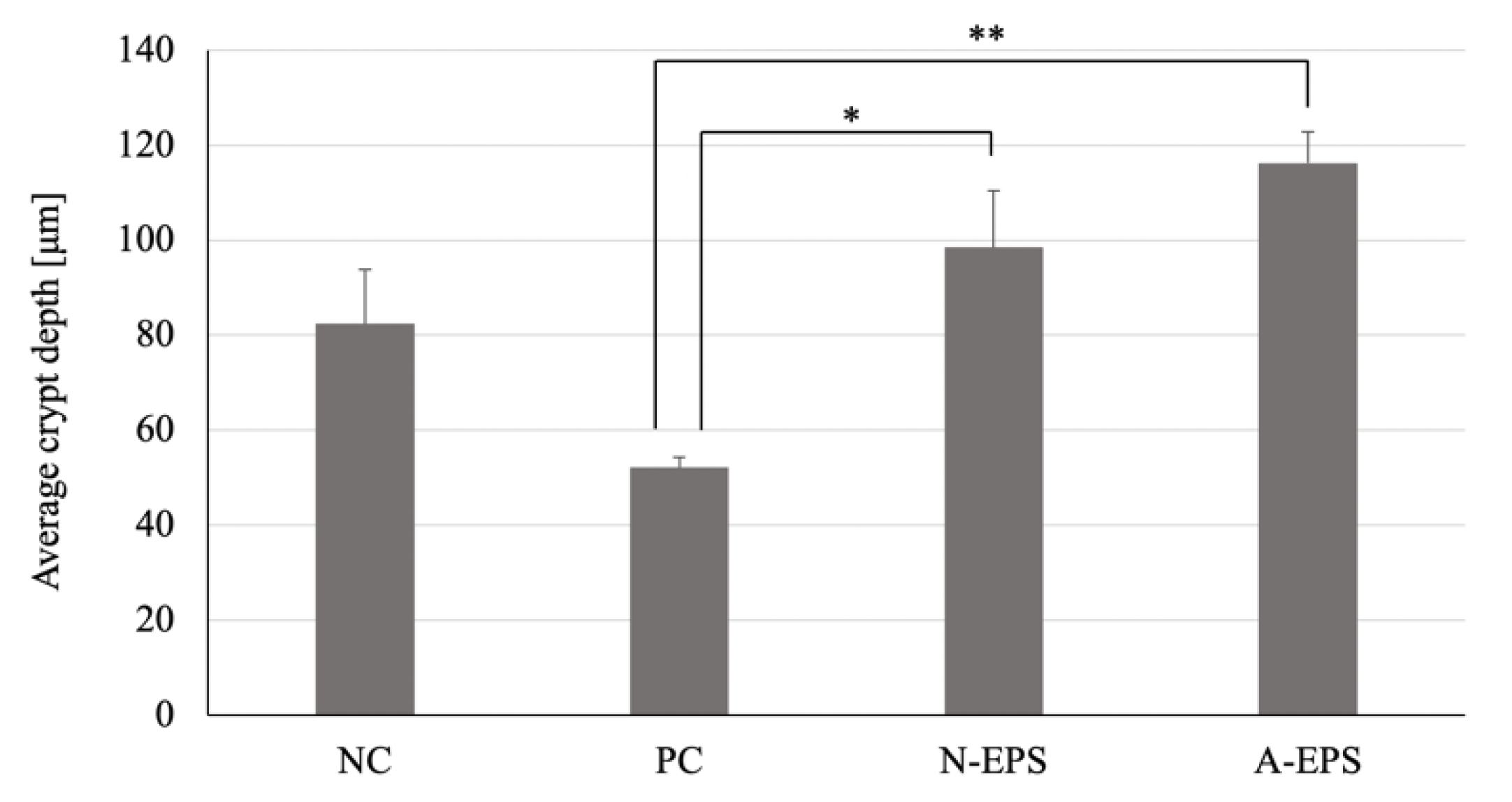
3.4. Histopathological Analysis of the Colon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2018, 390, 2769–2778. [Google Scholar] [CrossRef]
- GBD 2017 Inflammatory Bowel Disease Collaborators. The global, regional, and national burden of inflammatory bowel disease in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar] [CrossRef]
- Noth, R.; Stüber, E.; Häsler, R.; Nikolaus, S.; Kühbacher, T.; Hampe, J.; Bewig, B.; Schreiber, S.; Arlt, A. Anti-TNF-α antibodies improve intestinal barrier function in Crohn’s disease. J. Crohn’s Colitis 2012, 6, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salim, S.Y.; Söderholm, J.D. Importance of disrupted intestinal barrier in inflammatory bowel diseases. Inflamm. Bowel Dis. 2011, 17, 362–381. [Google Scholar] [CrossRef] [PubMed]
- Mowat, C.; Cole, A.; Windsor, A.; Ahmad, T.; Arnott, I.; Driscoll, R.; Mitton, S.; Orchard, T.; Rutter, M.; Younge, L.; et al. Guidelines for the management of inflammatory bowel disease in adults. Gut 2011, 60, 571–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triantafillidis, J.K.; Merikas, E.; Georgopoulos, F. Current and emerging drugs for the treatment of inflammatory bowel disease. Drug Des. Dev. Ther. 2011, 5, 185–210. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.E. Probiotics: Definition, sources, selection, and uses. Clin. Infect. Dis. 2008, 46, S58–S61. [Google Scholar] [CrossRef] [Green Version]
- Panthavee, W.; Noda, M.; Danshiitsoodol, N.; Kumagai, T.; Sugiyama, M. Characterization of exopolysaccharides produced by thermophilic lactic acid bacteria isolated from tropical fruits of Thailand. Biol. Pharm. Bull. 2017, 40, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Noda, M.; Sugimoto, S.; Hayashi, I.; Danshiitsoodol, N.; Fukamachi, M.; Sugiyama, M. A novel structure of exopolysaccharide produced by a plant-derived lactic acid bacterium Lactobacillus paracasei IJH-SONE68. J. Biochem. 2018, 164, 87–92. [Google Scholar] [CrossRef]
- Noda, M.; Shiraga, M.; Kumagai, T.; Danshiitsoodol, N.; Sugiyama, M. Characterization of the SN35N strain-specific exopolysaccharide encoded in the whole circular genome of a plant-derived Lactobacillus plantarum. Biol. Pharm. Bull. 2018, 41, 536–545. [Google Scholar] [CrossRef] [Green Version]
- Kakegawa, H.; Matsumoto, H.; Satoh, T. Activation of hyaluronidase by metallic salts and compound 48/80, and inhibitory effect of anti-allergic agents on hyaluronidase. Chem. Pharm. Bull. 1985, 33, 642–646. [Google Scholar] [CrossRef] [Green Version]
- Fujitani, N.; Sakaki, S.; Yamaguchi, Y.; Takenaka, H. Inhibitory effects of microalgae on the activation of hyaluronidase. J. Appl. Phycol. 2001, 13, 489–492. [Google Scholar] [CrossRef]
- Maeda, Y.; Yamamoto, M.; Masui, T.; Sugiyama, K.; Yokota, M.; Nakagomi, K.; Tanaka, H.; Takahashi, I.; Kobayashi, T.; Kobayashi, E. Inhibitory effect of tea extracts on hyaluronidase. Shokuhin Eiseigaku Zashi 1990, 31, 233–237. [Google Scholar] [CrossRef]
- Noda, M.; Sultana, N.; Hayashi, I.; Fukamachi, M.; Sugiyama, M. Exopolysaccharide produced by Lactobacillus paracasei IJH-SONE68 prevents and improves the picryl chloride-induced contact dermatitis. Molecules 2019, 24, 2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmel, S.A.; Roberts, R.F. Development of a growth medium suitable for exopolysaccharide production by Lactobacillus delbrueckii ssp. bulgaricus RR. Int. J. Food Microbiol. 1998, 40, 87–92. [Google Scholar] [CrossRef]
- Kets, E.P.W.; Galinski, E.A.; de Bont, J.A.M. Carnitine: A novel compatible solute in Lactobacillus plantarum. Arch. Microbiol. 1994, 162, 243–248. [Google Scholar] [CrossRef]
- Li, P.; Yu, Q.; Ye, X.; Wang, Z.; Yang, Q. Lactobacillus S-layer protein inhibition of Salmonella-induced reorganization of the cytoskeleton and activation of MAPK signaling pathways in Caco-2 cells. Microbiology 2011, 157, 2639–2646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Corredig, M.; Morales-Rayas, R.; Hassan, A.; Griffiths, M.W.; LaPointe, G. Effect of fermented milk from Lactococcus lactis ssp. cremoris strain JFR1 on Salmonella invasion of intestinal epithelial cells. J. Dairy Sci. 2019, 102, 6802–6819. [Google Scholar] [PubMed]
- Kim, J.J.; Shajib, M.S.; Manocha, M.M.; Khan, W.I. Investigating intestinal inflammation in DSS-induced model of IBD. J. Vis. Exp. 2012, 60, 3678. [Google Scholar] [CrossRef] [Green Version]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15.25.1–15.25.14. [Google Scholar] [CrossRef] [PubMed]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Smith, J.W.; Castro, G.A. Relation of peroxidase activity in gut mucosa to inflammation. Am. J. Physiol. 1978, 234, R72–R79. [Google Scholar] [CrossRef]
- Khan, A.K.A.; Piris, J.; Truelove, S.C. An experiment to determine the active therapeutic moiety of sulphasalazine. Lancet 1977, 2, 892–895. [Google Scholar] [CrossRef]
- Murray, A.; Nguyen, T.M.; Parker, C.E.; Feagan, B.G.; MacDonald, J.K. Oral 5-aminosalicylic acid for induction of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2020, 8, CD000543. [Google Scholar] [PubMed]
- Murray, A.; Nguyen, T.M.; Parker, C.E.; Feagan, B.G.; MacDonald, J.K. Oral 5-aminosalicylic acid for maintenance of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2020, 8, CD000544. [Google Scholar] [PubMed]
- Feuerstein, J.D.; Isaacs, K.L.; Schneider, Y.; Siddique, S.M.; Falck-Ytter, Y.; Singh, S.; AGA Institute Clinical Guidelines Committee. AGA clinical practice guidelines on the management of moderate to severe ulcerative colitis. Gastroenterology 2020, 158, 1450–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelrad, J.E.; Lichtiger, S.; Yajnik, V. Inflammatory bowel disease and cancer: The role of inflammation, immunosuppression, and cancer treatment. World J. Gastroenterol. 2016, 22, 4794–4801. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Lin, O.; Sinha, S.R. Use of tumor necrosis factor alpha inhibitors for inflammatory bowel disease patients with concurrent heart failure. Dig. Dis. Sci. 2017, 62, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyama, K.; Toyonaga, A.; Sasaki, E.; Watanabe, K.; Tateishi, H.; Nishiyama, T.; Saiki, T.; Ikeda, H.; Tsuruta, O.; Tanikawa, K. IL-8 as an important chemoattractant for neutrophils in ulcerative colitis and Crohn’s disease. Clin. Exp. Immunol. 1994, 96, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Waugh, D.J.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, J.C.; O’Loghlen, A.; Banito, A.; Guijarro, M.V.; Augert, A.; Raguz, S.; Fumagalli, M.; Da Costa, M.; Brown, C.; Popov, N.; et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell 2008, 133, 1006–1018. [Google Scholar] [CrossRef] [Green Version]
- Cotton, J.A.; Platnich, J.M.; Muruve, D.A.; Jijon, H.B.; Buret, A.G.; Beck, P.L. Interleukin-8 in gastrointestinal inflammation and malignancy: Induction and clinical consequences. Int. J. Interferon Cytokine Mediat. Res. 2016, 8, 13–34. [Google Scholar]
- Li, M.C.; He, S.H. IL-10 and its related cytokines for treatment of inflammatory bowel disease. World J. Gastroenterol. 2004, 10, 620–625. [Google Scholar] [CrossRef]
- Franke, A.; Balschun, T.; Karlsen, T.H.; Sventoraityte, J.; Nikolaus, S.; Mayr, G.; Domingues, F.S.; Albrecht, M.; Nothnagel, M.; Ellinghaus, D.; et al. Sequence variants in IL10, ARPC2 and multiple other loci contribute to ulcerative colitis susceptibility. Nat. Genet. 2008, 40, 1319–1323. [Google Scholar] [CrossRef]
- Franke, A.; McGovern, D.P.B.; Barrett, J.C.; Wang, K.; Radford-Smith, G.L.; Ahmad, T.; Lees, C.W.; Balschun, T.; Lee, J.; Roberts, R.; et al. Genome-wide meta-analysis increases to 71 the number of confirmed Crohn’s disease susceptibility loci. Nat. Genet. 2010, 42, 1118–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühn, R.; Löhler, J.; Rennick, D.; Rajewsky, K.; Müller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar] [CrossRef]
- Berg, D.J.; Davidson, N.; Kühn, R.; Müller, W.; Menon, S.; Holland, G.; Thompson-Snipes, L.; Leach, M.W.; Rennick, D. Enterocolitis and colon cancer in interleukin-10- deficient mice are associated with aberrant cytokine production and CD4(+) TH1-like responses. J. Clin. Investig. 1996, 98, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Yen, D.; Cheung, J.; Scheerens, H.; Poulet, F.; McClanahan, T.; Mckenzie, B.; Kleinschek, M.A.; Owyang, A.; Mattson, J.; Blumenschein, W.; et al. IL-23 is essential for T cell–mediated colitis and promotes inflammation via IL-17 and IL-6. J. Clin. Investig. 2006, 116, 1310–1316. [Google Scholar] [CrossRef]
- Kullberg, M.C.; Jankovic, D.; Feng, C.G.; Hue, S.; Gorelick, P.L.; McKenzie, B.S.; Cua, D.J.; Powrie, F.; Cheever, A.W.; Maloy, K.J.; et al. IL-23 plays a key role in Helicobacter hepaticus-induced T cell-dependent colitis. J. Exp. Med. 2006, 203, 2485–2494. [Google Scholar] [CrossRef] [Green Version]
- Ogino, H.; Nakamura, K.; Ihara, E.; Akiho, H.; Takayanagi, R. CD4+CD25+ regulatory T cells suppress Th17-responses in an experimental colitis model. Dig. Dis. Sci. 2011, 56, 376–386. [Google Scholar] [CrossRef]
- Kamada, N.; Hisamatsu, T.; Okamoto, S.; Sato, T.; Matsuoka, K.; Arai, K.; Nakai, T.; Hasegawa, A.; Inoue, N.; Watanabe, N.; et al. Abnormally differentiated subsets of intestinal macrophage play a key role in Th1-dominant chronic colitis through excess production of IL-12 and IL-23 in response to bacteria. J. Immunol. 2005, 175, 6900–6908. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Sato, A.; Goto, A.; Nakamura, M.; Ogawa, M.; Chiba, Y.; Hemmi, J.; Kano, H.; Takeda, K.; Okumura, K.; et al. Enhanced natural killer cell activation by exopolysaccharides derived from yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. J. Dairy Sci. 2016, 99, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Vinderola, G.; Perdigón, G.; Duarte, J.; Farnworth, E.; Matar, C. Effect of the oral administration of the exopolysaccharide produce by Lactobacillus kefiranofaciens on the gut mucosal immunity. Cytokine 2006, 36, 254–260. [Google Scholar] [CrossRef]
- Sasaki, S.; Yoneyama, H.; Suzuki, K.; Suriki, H.; Aiba, T.; Watanabe, S.; Kawauchi, Y.; Kawachi, H.; Shimizu, F.; Matsushima, K.; et al. Blockade of CXCL10 protects mice from acute colitis and enhances crypt cell survival. Eur. J. Immunol. 2002, 32, 3197–3205. [Google Scholar] [CrossRef]
- Potten, C.S. Stem cells in gastrointestinal epithelium: Numbers, characteristics and death. Philos. Trans. R. Soc. Lond. B. 1998, 353, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Snippert, H.J. Colonic Crypts: Safe Haven from Microbial Products. Cell 2016, 165, 1564–1566. [Google Scholar] [CrossRef] [Green Version]
- Hermiston, M.L.; Gordon, J.I. Inflammatory bowel diseases and adenomas in mice expressing a dominant negative N-cadherin. Science 1995, 270, 1203–1207. [Google Scholar] [CrossRef]
- Hermiston, M.L.; Wong, M.H.; Gordon, J.I. Forced expression of E-cadherin in the mouse intestinal epithelium slows cell migration and provides evidence for nonautonomous regulation of cell fate in a self-renewing system. Genes Dev. 1996, 10, 985–996. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; de Vera, J.; Narushima, S.; Beck, E.X.; Palencia, S.; Shinkawa, P.; Kim, K.A.; Liu, Y.; Levy, M.D.; Berg, D.J.; et al. R-spondin1, a novel intestinotrophic mitogen, ameliorates experimental colitis in mice. Gastroenterology 2007, 132, 1331–1343. [Google Scholar] [CrossRef]
- Qi, K.; Lv, J.; Wu, J.; Xu, Z. Therapeutic effects of different doses of polyethylene glycosylated porcine glucagon-like peptide-2 on ulcerative colitis in male rats. BMC Gastroenterol. 2017, 17, 34. [Google Scholar] [CrossRef] [Green Version]
- L’Heureux, M.C.; Brubaker, P.L. GLP-2 and DSS glucagon-like peptide-2 and common therapeutics in a murine model of ulcerative colitis. J. Pharmacol. Exp. Ther. 2003, 306, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Tafazoli, F.; Magnusson, K.E.; Zheng, L. Disruption of epithelial barrier integrity by Salmonella enterica serovar Typhimurium requires geranylgeranylated proteins. Infect. Immun. 2003, 71, 872–881. [Google Scholar] [CrossRef] [Green Version]
- Harrer, A.; Bücker, R.; Boehm, M.; Zarzecka, U.; Tegtmeyer, N.; Sticht, H.; Schulzke, J.D.; Backert, S. Campylobacter jejuni enters gut epithelial cells and impairs intestinal barrier function through cleavage of occludin by serine protease HtrA. Gut Pathog. 2019, 11, 4. [Google Scholar] [CrossRef]
- Freudenberg, M.A.; Merlin, T.; Gumenscheimer, M.; Kalis, C.; Landmann, R.; Galanos, C. Role of lipopolysaccharide susceptibility in the innate immune response to Salmonella typhimurium infection: LPS, a primary target for recognition of Gram-negative bacteria. Microb. Infect. 2001, 3, 1213–1222. [Google Scholar] [CrossRef]
- Rathinam, V.A.K.; Appledorn, D.M.; Hoag, K.A.; Amalfitano, A.; Mansfield, L.S. Campylobacter jejuni-induced activation of dendritic cells involves cooperative signaling through Toll-like receptor 4 (TLR4)-MyD88 and TLR4-TRIF axes. Infect. Immun. 2009, 77, 2499–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, M.J. Living in the danger zone: Innate immunity to Salmonella. Curr. Opin. Microb. 2004, 7, 51–57. [Google Scholar] [CrossRef] [PubMed]
- De Zoete, M.R.; Keestra, A.M.; Wagenaar, J.A.; van Putten, J.P.M. Reconstitution of a functional toll-like receptor 5 binding site in Campylobacter jejuni flagellin. J. Biol. Chem. 2010, 285, 12149–12158. [Google Scholar] [CrossRef] [Green Version]
- Deguine, J.; Barton, G.M. MyD88: A central player in innate immune signaling. F1000Prime Rep. 2014, 6, 97. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Bowie, A.G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signaling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef]
- Tohno, M.; Shimizu, T.; Ueda, W.; Anzawa, D.H.; Aso, H.; Nishimura, J.; Kawai, Y.; Saito, Y.; Saito, T.; Kitazawa, H. Molecular cloning of porcine RP105/MD-1 involved in recognition of extracellular phosphopolysaccharides from Lactococcus lactis ssp. cremoris. Mol. Immunol. 2007, 44, 2566–2577. [Google Scholar] [CrossRef]
- Makino, S.; Ikegami, S.; Kano, H.; Sashihara, T.; Sugano, H.; Horiuchi, H.; Saito, T.; Oda, M. Immunomodulatory effects of polysaccharides produced by Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. J. Dairy Sci. 2006, 89, 2873–2881. [Google Scholar] [CrossRef] [Green Version]
- Kitazawa, H.; Harata, T.; Uemura, J.; Saito, T.; Kaneko, T.; Itoh, T. Phosphate group requirement for mitogenic activation of lymphocytes by an extracellular phosphopolysaccharide from Lactobacillus delbrueckii ssp. bulgaricus. Int. J. Food Microbiol. 1998, 40, 169–175. [Google Scholar] [CrossRef]
- Kitazawa, H.; Itoh, T.; Tomioka, Y.; Mizugaki, M.; Yamaguchi, T. Induction of IFN-γ and IL-1α production in macrophages stimulated with phosphopolysaccharide produced by Lactococcus lactis ssp. cremoris. Int. J. Food Microbiol. 1996, 31, 99–106. [Google Scholar] [CrossRef]
- Hashiguchi, K.; Nagata, Y.; Yoshida, M.; Murohushi, Y.; Kitazawa, H. Chemical and immunological characterization of extracellular polysaccharides produced by Lactobacillus plantarum No 14. Jpn. J. Lact. Acid Bact. 2011, 22, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Narula, N.; Kassam, Z.; Yuan, Y.; Colombel, J.F.; Ponsioen, C.; Reinisch, W.; Moayyedi, P. Systematic review and meta-analysis: Fecal microbiota transplantation for treatment of active ulcerative colitis. Inflamm. Bowel Dis. 2017, 23, 1702–1709. [Google Scholar] [CrossRef]
- Ianiro, G.; Maida, M.; Burisch, J.; Simonelli, C.; Hold, G.; Ventimiglia, M.; Gasbarrini, A.; Cammarota, G. Efficacy of different faecal microbiota transplantation protocols for Clostridium difficile infection: A systematic review and meta-analysis. United Eur. Gastroenterol. J. 2018, 6, 1232–1244. [Google Scholar] [CrossRef] [Green Version]
- Cammarota, G.; Ianiro, G.; Tilg, H.; Rajilić-Stojanović, M.; Kump, P.; Satokari, R.; Sokol, H.; Arkkila, P.; Pintus, C.; Hart, A.; et al. European consensus conference on faecal microbiota transplantation in clinical practice. Gut 2017, 66, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Kootte, R.S.; Levin, E.; Salojärvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of insulin sensitivity after lean donor feces in metabolic syndrome is driven by baseline intestinal microbiota composition. Cell Metab. 2017, 26, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Xu, M.; Wang, W.; Cao, X.; Piao, M.; Khan, S.; Yan, F.; Cao, H.; Wang, B. Systematic review: Adverse events of fecal microbiota transplantation. PLoS ONE 2016, 11, e0161174. [Google Scholar] [CrossRef] [Green Version]
- FDA. Important Safety Alert Regarding Use of Fecal Microbiota for Transplantation and Risk of Serious Adverse Reactions Due to Transmission of Multi-Drug Resistant Organisms; U.S. Food & Drug Administration: Hampton, VA, USA, 2019.
- Pabón-Carrasco, M.; Ramirez-Baena, L.; Vilar-Palomo, S.; Castro-Méndez, A.; Martos-García, R.; Rodríguez-Gallego, I. Probiotics as a coadjuvant factor in active or quiescent inflammatory bowel disease of adults—A meta-analytical study. Nutrients 2020, 12, 2628. [Google Scholar] [CrossRef]
- Emamie, A.D.; Rajabpour, M.; Ghanavati, R.; Asadolahi, P.; Farzi, S.; Sobouti, B.; Darbandi, A. The effects of probiotics, prebiotics and synbiotics on the reduction of IBD complications, a periodic review during 2009–2020. J. Appl. Microbiol. 2020, 130, 1823–1838. [Google Scholar] [CrossRef]
- Furukawa, N.; Matsuoka, A.; Takahashi, T.; Yamanaka, Y. Anti-metastatic effect of kefir grain components on Lewis lung carcinoma and highly metastatic B16 melanoma in mice. J. Agric. Sci. Tokyo Univ. Agric. 2020, 45, 62–70. [Google Scholar]
- Sengül, N.; Işık, S.; Aslım, B.; Uçar, G.; Demirbağ, A.E. The effect of exopolysaccharide-producing probiotic strains on gut oxidative damage in experimental colitis. Dig. Dis. Sci. 2011, 56, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Cantabrana, C.; López, P.; Gueimonde, M.; de Los Reyes-Gavilán, C.G.; Suárez, A.; Margolles, A.; Ruas-Madiedo, P. Immune modulation capability of exopolysaccharides synthesised by lactic acid bacteria and bifidobacteria. Probiotics Antimicrob. Proteins 2012, 4, 227–237. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Stool Consistency | Hematochezia |
|---|---|---|
| 0 | Normal stools | Normal stools |
| 1 | Soft pellets; can be picked up by tweezers | Pale red to red-colored stools; blood spreads only surface on the stools |
| 2 | Loose stools; cannot be picked up by tweezers | Red to deep red-colored stools; blood spreads in the stools |
| 3 | Diarrhea | Deep red to dark red-colored stools; gross bleeding |
| Name | Sequence (5′ → 3′) | Target Products |
|---|---|---|
| mβ-actin-F | GGGACGACATGGAGAAGA | β-actin |
| mβ-actin-R | CATACAGGGACAGCACAG | |
| mCOX-2-F | TCAGTAGGTTTTTGCTGTGAGG | COX-2 |
| mCOX-2-R | GTTCAATGGGCTGGAAGACA | |
| miNOS-F | GAAGAAAACCCCTTGTGCTG | iNOS |
| miNOS-R | TTCTGTGCTGTCCCAGTGAG | |
| mTNF-α-F | CATCTTCTCAAAATTCGAGTGACAA | TNF-α |
| mTNF-α-R | TGGGAGTAGACAAGGTACAACCC | |
| mIL-6-F | GAGGATACCACTCCCAACAGACC | IL-6 |
| mIL-6-R | AAGTGCATCATCGTTGTTCATACA | |
| mMIP2-F | AGTGAACTGCGCTGTCAATG | MIP-2 |
| mMIP2-R | CAAGGCAAACTTTTTGACCG | |
| mIL-10-F | GGTTGCCAAGCCTTATCGGA | IL-10 |
| mIL-10-R | ACCTGCTCCACTGCCTTGCT | |
| mIL-12-F | AGGACTTGAAGATGTACCAG | IL-12 |
| mIL-12-R | CTATCTGTGTGAGGAGGG | |
| mIL-17-F | GCTCCAGAAGGCCCTCAGA | IL-17 |
| mIL-17-R | AGCTTTCCCTCCGCATTGA | |
| mIL-18-F | CAGGCCTGACATCTTCTGCAA | IL-18 |
| mIL-18-R | TCTGACATGGCAGCCATTGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noda, M.; Danshiitsoodol, N.; Kanno, K.; Uchida, T.; Sugiyama, M. The Exopolysaccharide Produced by Lactobacillus paracasei IJH-SONE68 Prevents and Ameliorates Inflammatory Responses in DSS–Induced Ulcerative Colitis. Microorganisms 2021, 9, 2243. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112243
Noda M, Danshiitsoodol N, Kanno K, Uchida T, Sugiyama M. The Exopolysaccharide Produced by Lactobacillus paracasei IJH-SONE68 Prevents and Ameliorates Inflammatory Responses in DSS–Induced Ulcerative Colitis. Microorganisms. 2021; 9(11):2243. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112243
Chicago/Turabian StyleNoda, Masafumi, Narandalai Danshiitsoodol, Keishi Kanno, Tomoyuki Uchida, and Masanori Sugiyama. 2021. "The Exopolysaccharide Produced by Lactobacillus paracasei IJH-SONE68 Prevents and Ameliorates Inflammatory Responses in DSS–Induced Ulcerative Colitis" Microorganisms 9, no. 11: 2243. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112243