
Probiotic Properties of Lactic Acid Bacteria with High Conjugated Linoleic Acid Converting Activity Isolated from Jeot-Gal, High-Salt Fermented Seafood


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Screening and Identification of LAB with LA to CLA Conversion Activity
2.3. Acid and Bile Tolerance of Isolated Strains
2.4. Antimicrobial Activity
2.5. Cell Adherence Assay
2.6. In Vivo Caenorhabditis Elegans Assay for Immune Response
2.7. qRT-PCR for Inflammatory Cytokine Assay
2.8. Statistical Analysis
3. Results
3.1. Screening of CLA-Producing LAB
3.2. Identification of CLA-Producing LAB
3.3. Isomeric Composition Analysis of CLA by GC
3.4. pH and Bile Salt Tolerance of CLA-Converting LAB
3.5. Antimicrobial Activity
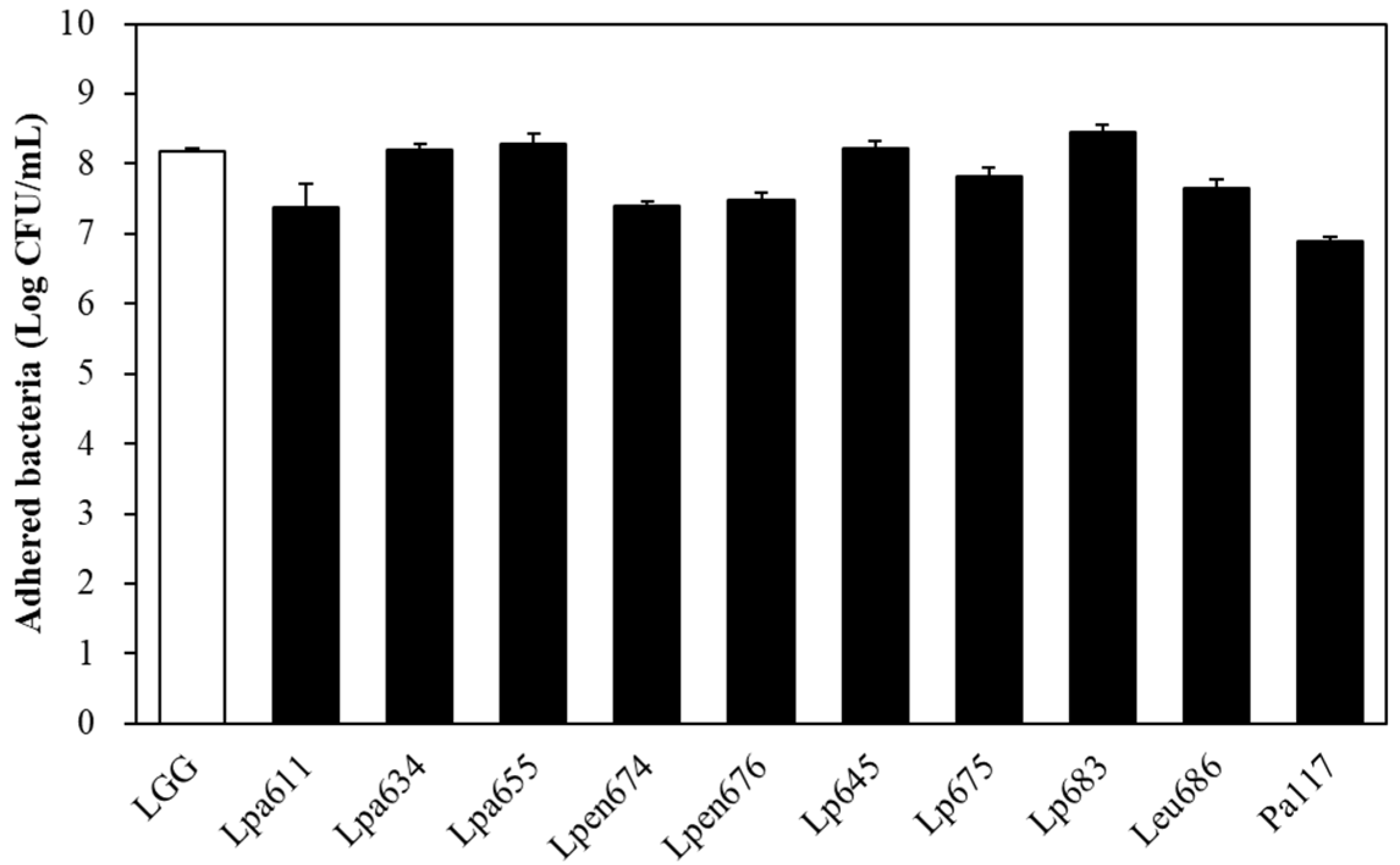
3.6. Intestinal Adhesion Ability of LAB Strains to Caco-2 Cells
3.7. Effects of LAB Strains on Growth and Cytokine Secretion in RAW 264.7 Cells
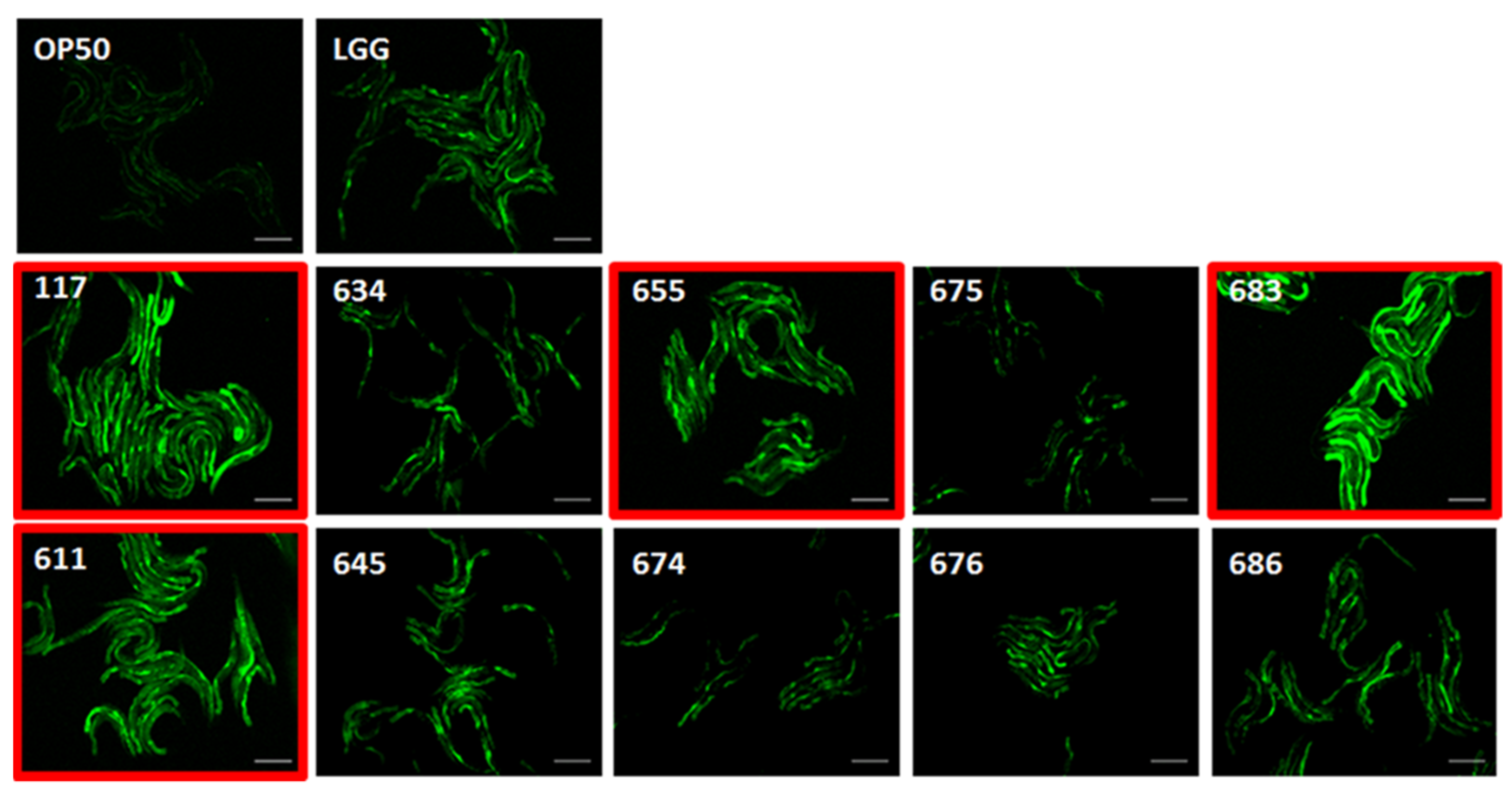
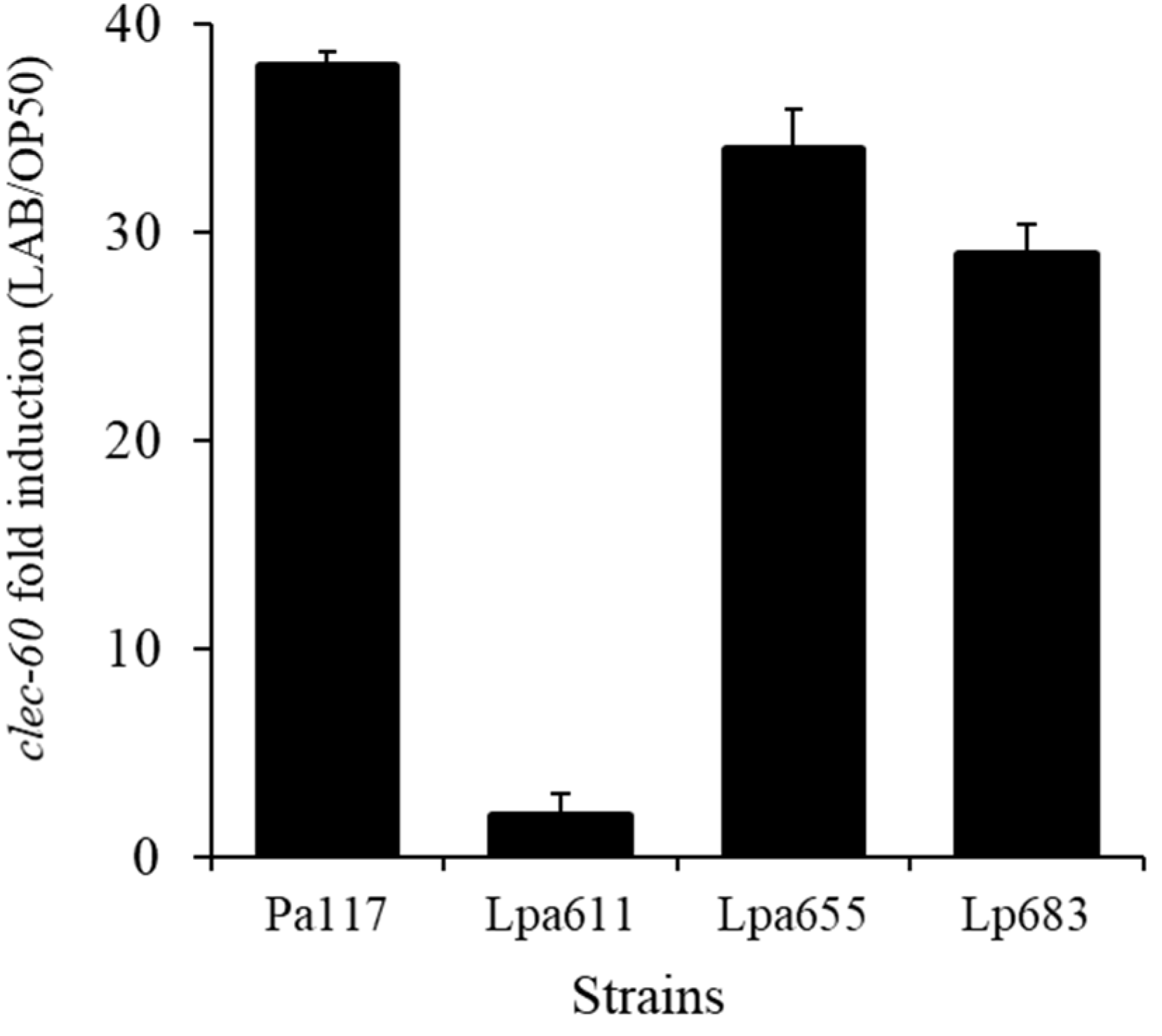
3.8. Effects of LAB Strains on In Vivo Caenorhabditis Elegans Assay for Immune Response
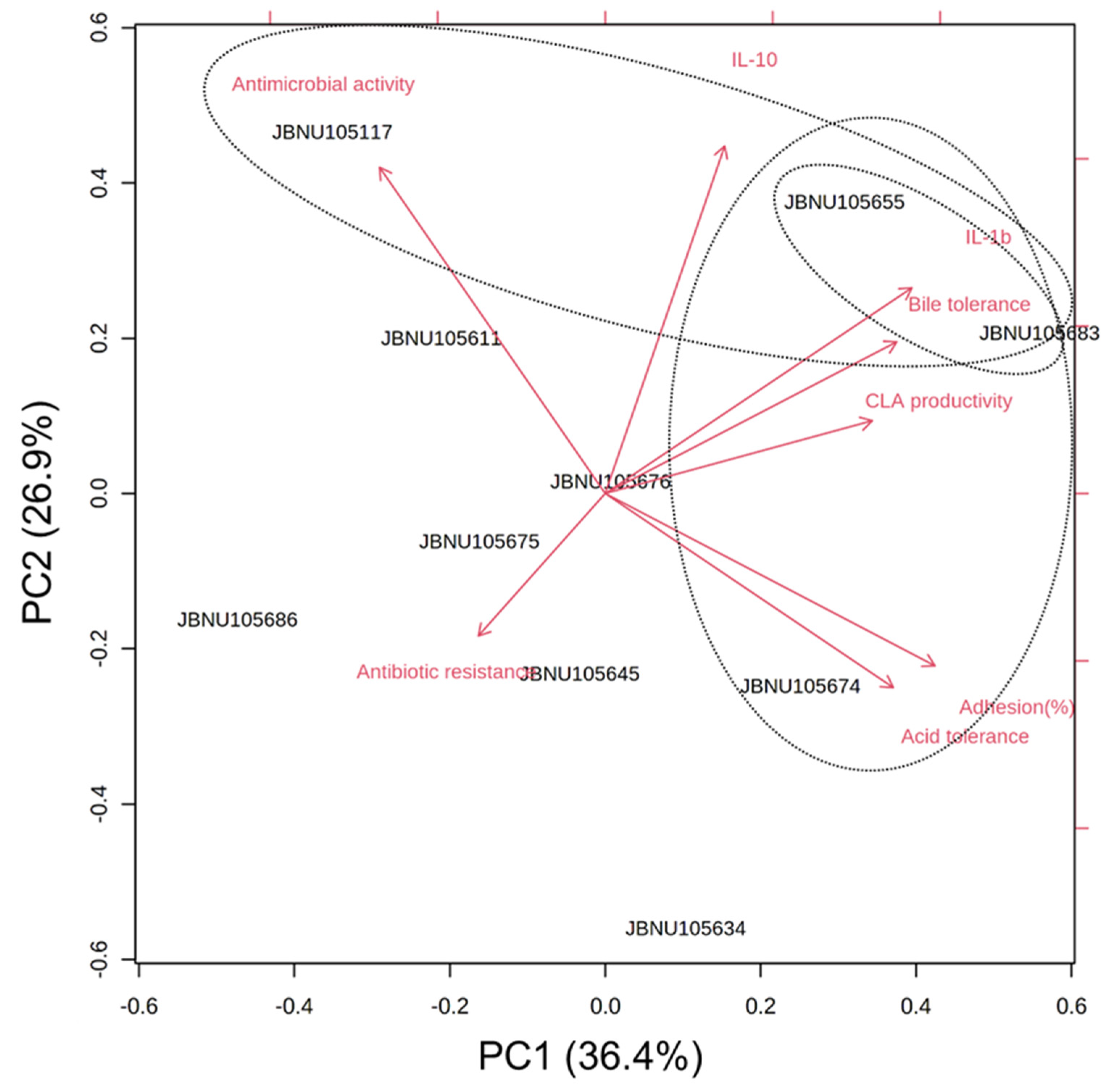
3.9. PCA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, K.W.; Park, J.Y.; Sa, H.D.; Jeong, J.H.; Jin, D.E.; Heo, H.J. Probiotic properties of Pediococcus strains isolated from jeotgals, salted and fermented Korean sea-food. Anaerobe 2014, 28, 199–206. [Google Scholar] [CrossRef]
- Um, M.N.; Lee, C.H. Isolation and identification of Staphylococcus sp. from Korean fermented fish products. J. Microbiol. Biotechnol. 1996, 6, 340–346. [Google Scholar]
- Kim, S.J.; Ma, S.J.; Kim, H.L. Probiotic properties of lactic acid bacteria and yeasts isolated from korean traditional food, Jeot-gal. Korean J. Food Preserv. 2005, 12, 184–189. [Google Scholar]
- Roh, S.W.; Kim, K.H.; Nam, Y.D.; Chang, H.W.; Park, E.J.; Bae, J.W. Investigation of archaeal and bacterial diversity in fermented seafood using barcoded pyrosequencing. ISME J. 2010, 4, 1–16. [Google Scholar] [CrossRef]
- Jeong, S.H.; Lee, J.H.; Jung, J.Y.; Lee, S.H.; Park, M.S.; Jeon, C.O. Halomonas cibimaris sp. nov., isolated from jeotgal, a traditional Korean fermented seafood. Antonie Leeuwenhoek 2013, 103, 503–512. [Google Scholar] [CrossRef]
- Jung, M.J.; Roh, S.W.; Kim, M.S.; Bae, J.W. Lentibacillus jeotgali sp. nov., a halophilic bacterium isolated from traditional Korean fermented seafood. Int. J. Syst. Evol. Microbiol. 2010, 60, 1017–1022. [Google Scholar] [CrossRef]
- Roh, S.W.; Nam, Y.-D.; Chang, H.-W.; Sung, Y.; Kim, K.-H.; Oh, H.-M.; Bae, J.-W. Halalkalicoccus jeotgali sp. nov., a halophilic archaeon from shrimp jeotgal, a traditional Korean fermented seafood. Int. J. Syst. Evol. Microbiol. 2007, 57, 2296–2298. [Google Scholar] [CrossRef]
- Yoon, J.H.; Lee, K.C.; Kho, Y.H.; Kang, K.H.; Kim, C.J.; Park, Y.H. Halomonas alimentaria sp. nov., isolated from jeotgal, a traditional Korean fermented seafood. Int. J. Syst. Evol. Microbiol. 2002, 52, 123–130. [Google Scholar] [CrossRef]
- Jeun, J.; Kim, H.; Lee, H.; Ryu, B. Isolation and identification of Lactobacillus sp. produced γ-aminobutyric acid (GABA) from traditional salt fermented anchovy. Korean J. Food Nutr. 2004, 17, 72–79. [Google Scholar]
- Lee, N.K.; Kim, T.H.; Choi, S.Y.; Lee, S.K.; Paik, H.D. Identification and Probiotic Properties of Lactococcus lactis NK24 Isolated from Jeot-gal, a Korean Fermented Food. Food Sci. Biotechnol. 2004, 13, 411–416. [Google Scholar]
- McCrorie, T.A.; Keaveney, E.M.; Wallace, J.M.; Binns, N.; Livingstone, M.B.E. Human health effects of conjugated linoleic acid from milk and supplements. Nutr. Res. Rev. 2011, 24, 206–227. [Google Scholar] [CrossRef] [Green Version]
- Koba, K.; Yanagita, T. Health benefits of conjugated linoleic acid (CLA). Obes. Res. Clin. Pract. 2014, 8, e525–e532. [Google Scholar] [CrossRef]
- Albers, R.; Van der Wielen, R.; Brink, E.; Hendriks, H.; Dorovska-Taran, V.; Mohede, I. Effects of cis-9, trans-11 and trans-10, cis-12 conjugated linoleic acid (CLA) isomers on immune function in healthy men. Eur. J. Clin. Nutr. 2003, 57, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Chin, S.; Liu, W.; Storkson, J.; Ha, Y.; Pariza, M. Dietary sources of conjugated dienoic isomers of linoleic acid, a newly recognized class of anticarcinogens. J. Food Compos. Anal. 1992, 5, 185–197. [Google Scholar] [CrossRef]
- Kritchevsky, D.; Tepper, S.A.; Wright, S.; Czarnecki, S.K.; Wilson, T.A.; Nicolosi, R.J. Conjugated linoleic acid isomer effects in atherosclerosis: Growth and regression of lesions. Lipids 2004, 39, 611–616. [Google Scholar] [CrossRef]
- Brown, J.M.; McIntosh, M.K. Conjugated linoleic acid in humans: Regulation of adiposity and insulin sensitivity. J. Nutr. 2003, 133, 3041–3046. [Google Scholar] [CrossRef] [Green Version]
- Azain, M.J.; Hausman, D.B.; Sisk, M.B.; Flatt, W.P.; Jewell, D.E. Dietary conjugated linoleic acid reduces rat adipose tissue cell size rather than cell number. J. Nutr. 2000, 130, 1548–1554. [Google Scholar] [CrossRef]
- Lee, K.; Paek, K.; Lee, H.Y.; Park, J.H.; Lee, Y. Antiobesity effect of trans-10, cis-12-conjugated linoleic acid-producing Lactobacillus plantarum PL62 on diet-induced obese mice. J. Appl. Microbiol. 2007, 103, 1140–1146. [Google Scholar] [CrossRef]
- Li, C.; Wang, J.; Wang, H.; Cai, S.; Mu, G.; Zou, W.; Zhang, X.; Tian, L.; Chen, H. Effect of trans10, cis12-conjugated linoleic acid on glucose consumption of insulin resistance chang liver cells. J. Food Biochem. 2011, 35, 1593–1602. [Google Scholar] [CrossRef]
- Stringer, D.M.; Zahradka, P.; DeClercq, V.C.; Ryz, N.R.; Diakiw, R.; Burr, L.L.; Xie, X.; Taylor, C.G. Modulation of lipid droplet size and lipid droplet proteins by trans-10, cis-12 conjugated linoleic acid parallels improvements in hepatic steatosis in obese, insulin-resistant rats. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2010, 1801, 1375–1385. [Google Scholar] [CrossRef]
- Liu, X.; Li, H.; Chen, Y.; Cao, Y. Method for screening of bacterial strains biosynthesizing specific conjugated linoleic acid isomers. J. Agric. Food Chem. 2012, 60, 9705–9710. [Google Scholar] [CrossRef]
- Steven, W.; Belury, M.A. Impact of conjugated linoleic acid on bone physiology: Proposed mechanism involving inhibition of adipogenesis. Nutr. Rev. 2011, 69, 123–131. [Google Scholar]
- Chin, S.F.; Storkson, J.M.; Albright, K.J.; Cook, M.E.; Pariza, M.W. Conjugated linoleic acid is a growth factor for rats as shown by enhanced weight gain and improved feed efficiency. J. Nutr. 1994, 124, 2344–2349. [Google Scholar] [CrossRef]
- Schmid, A.; Collomb, M.; Sieber, R.; Bee, G. Conjugated linoleic acid in meat and meat products: A review. Meat Sci. 2006, 73, 29–41. [Google Scholar] [CrossRef]
- Andrade, J.C.; Ascencao, K.; Gullon, P.; Henriques, S.; Pinto, J.; Rocha-Santos, T.A.; Freitas, A.C.; Gomes, A.M. Production of conjugated linoleic acid by food-grade bacteria: A review. Int. J. Dairy Technol. 2012, 65, 467–481. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Stanton, C.; Yang, B.; Ross, R.P.; Silva, C.C. Conjugated linoleic acid production and probiotic assessment of Lactobacillus plantarum isolated from Pico cheese. LWT Food Sci. Technol. 2018, 90, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Di Cerbo, A.; Palmieri, B.; Aponte, M.; Morales-Medina, J.C.; Iannitti, T. Mechanisms and therapeutic effectiveness of lactobacilli. J. Clin. Pathol. 2016, 69, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Riedel, C.U.; Foata, F.; Philippe, D.; Adolfsson, O.; Eikmanns, B.J.; Blum, S. Anti-inflammatory effects of bifidobacteria by inhibition of LPS-induced NF-κB activation. World J. Gastroenterol. 2006, 12, 3729–3735. [Google Scholar] [CrossRef]
- Ikeda, T.; Yasui, C.; Hoshino, K.; Arikawa, K.; Nishikawa, Y. Influence of lactic acid bacteria on longevity of Caenorhabditis elegans and host defense against Salmonella enterica serovar enteritidis. Appl. Environ. Microbiol. 2007, 73, 6404–6409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’Halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C. In vitro selection criteria for probiotic bacteria of human origin: Correlation with in vivo findings. Am. J. Clin. Nutr. 2001, 73, 386s–392s. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Choe, J.; Kim, J.; Oh, S.; Park, S.; Kim, S.; Kim, Y. Heat-killed Lactobacillus spp. cells enhance survivals of Caenorhabditis elegans against Salmonella and Yersinia infections. Lett. Appl. Microbiol. 2015, 61, 523–530. [Google Scholar] [CrossRef]
- Jang, S.Y.; Heo, J.; Park, M.R.; Song, M.H.; Kim, J.N.; Jo, S.H.; Jeong, D.Y.; Lee, H.K.; Kim, Y.; Oh, S. Genome characteristics of Lactobacillus fermentum strain JDFM216 for application as probiotic bacteria. J. Microbiol. Biotechnol. 2017, 27, 1266–1271. [Google Scholar] [CrossRef]
- Romero-Perez, G.A.; Inoue, R.; Ushida, K.; Yajima, T. A Rapid Method of Screening Lactic Acid Bacterial Strains for Conjugated Linoleic Acid Production. Biosci. Biotechnol. Biochem. 2013, 77, 648–650. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Baik, S.H. Probiotic properties of Lactobacillus strains with high cinnamoyl esterase activity isolated from jeot-gal, a high-salt fermented seafood. Ann. Microbiol. 2019, 69, 407–417. [Google Scholar] [CrossRef]
- Beppu, F.; Hosokawa, M.; Tanaka, L.; Kohno, H.; Tanaka, T.; Miyashita, K. Potent inhibitory effect of trans 9, trans 11 isomer of conjugated linoleic acid on the growth of human colon cancer cells. J. Nutr. Biochem. 2006, 17, 830–836. [Google Scholar] [CrossRef] [Green Version]
- Roach, J.A.; Mossoba, M.M.; Yurawecz, M.P.; Kramer, J.K. Chromatographic separation and identification of conjugated linoleic acid isomers. Anal. Chim. Acta 2002, 465, 207–226. [Google Scholar] [CrossRef]
- Liu, P.; Shen, S.R.; Ruan, H.; Zhou, Q.; Ma, L.L.; He, G.Q. Production of conjugated linoleic acids by Lactobacillus plantarum strains isolated from naturally fermented Chinese pickles. J. Zhejiang Univ. Sci. B 2011, 12, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lievin, V.; Peiffer, I.; Hudault, S.; Rochat, F.; Brassart, D.; Neeser, J.; Servin, A. Bifidobacterium strains from resident infant human gastrointestinal microflora exert antimicrobial activity. Gut 2000, 47, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.K.; Courtney, P.D. Lactobacillus growth and membrane composition in the presence of linoleic or conjugated linoleic acid. Can. J. Microbiol. 2003, 49, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C. The effects of fatty acids on pure cultures of rumen bacteria. J. Agric. Sci. 1973, 81, 107–112. [Google Scholar] [CrossRef]
- Jiang, J.; Björck, L.; Fonden, R. Production of conjugated linoleic acid by dairy starter cultures. J. Appl. Microbiol. 1998, 85, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Alonso, L.; Cuesta, E.P.; Gilliland, S.E. Production of free conjugated linoleic acid by Lactobacillus acidophilus and Lactobacillus casei of human intestinal origin. Int. J. Dairy Sci. 2003, 86, 1941–1946. [Google Scholar] [CrossRef] [Green Version]
- Vela Gurovic, M.S.; Gentili, A.R.; Olivera, N.L.; Rodriguez, M.S. Lactic acid bacteria isolated from fish gut produce conjugated linoleic acid without the addition of exogenous substrate. Process Biochem. 2014, 49, 1071–1077. [Google Scholar] [CrossRef]
- Kishino, S.; Takeuchi, M.; Park, S.B.; Hirata, A.; Kitamura, N.; Kunisawa, J.; Kiyono, H.; Iwamoto, R.; Isobe, Y.; Arita, M.; et al. Polyunsaturated fatty acid saturation by gut lactic acid bacteria affecting host lipid composition. Proc. Natl. Acad. Sci. USA 2013, 110, 17808–17813. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Chen, H.; Gu, Z.; Tian, F.; Ross, R.P.; Stanton, C.; Chen, Y.Q.; Chen, W.; Zhang, H. Synthesis of conjugated linoleic acid by the linoleate isomerase complex in food-derived lactobacilli. J. Appl. Microbiol. 2014, 117, 430–439. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Anaya, J.; Hernández-Santoyo, A. Production of bioactive conjugated linoleic acid by the multifunctional enolase from Lactobacillus plantarum. Int. J. Biol. Macromol. 2016, 91, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Coakley, M.; Ross, R.; Nordgren, M.; Fitzgerald, G.; Devery, R.; Stanton, C. Conjugated linoleic acid biosynthesis by human-derived Bifidobacterium species. J. Appl. Microbiol. 2003, 94, 138–145. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Haberer, P.; Snel, J.; Schillinger, U.; Veld, J.H.H.I. Overview of gut flora and probiotics. Int. J. Food Microbiol. 1998, 41, 85–101. [Google Scholar] [CrossRef]
- Singh, T.P.; Kaur, G.; Malik, R.K.; Schillinger, U.; Guigas, C.; Kapila, S. Characterization of intestinal Lactobacillus reuteri strains as potential probiotics. Probiotics Antimicrob. Proteins 2012, 4, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Mathara, J.M.; Schillinger, U.; Kutima, P.M.; Mbugua, S.K.; Guigas, C.; Franz, C.; Holzapfel, W.H. Functional properties of Lactobacillus plantarum strains isolated from Maasai traditional fermented milk products in Kenya. Curr. Microbiol. 2008, 56, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Shah, N. Acid, bile, and heat tolerance of free and microencapsulated probiotic bacteria. J. Food Sci. 2007, 72, M446–M450. [Google Scholar] [CrossRef] [PubMed]
- Perdigon, G.; Galdeano, C.M.; Valdez, J.C.; Medici, M. Interaction of lactic acid bacteria with the gut immune system. Eur. J. Clin. Nutr. 2002, 56, S21. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, T.; Yamazaki, R.; Hashimoto, S.; Yokokura, T. The effect of oral feeding of Lactobacillus casei strain Shirota on immunoglobulin E production in mice. J. Dairy Sci. 1998, 81, 48–53. [Google Scholar] [CrossRef]
- Pochard, P.; Gosset, P.; Grangette, C.; Andre, C.; Tonnel, A.B. Lactic acid bacteria inhibit Th2 cytokine production by mononuclear cells from allergic patients. J. Allergy Clin. Immunol. 2002, 110, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Von der Weid, T.; Bulliard, C.; Schiffrin, E.J. Induction by a lactic acid bacterium of a population of CD4+ T cells with low proliferative capacity that produce transforming growth factor β and interleukin-10. Clin. Diagn. Lab. Immunol. 2001, 8, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boirivant, M.; Strober, W. The mechanism of action of probiotics. Curr. Opin. Gastroenterol. 2007, 23, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Won, T.J.; Kim, B.; Song, D.S.; Lim, Y.T.; Oh, E.S.; Lee, D.I.; Park, E.S.; Min, H.; Park, S.Y.; Hwang, K.W. Modulation of Th1/Th2 balance by Lactobacillus strains isolated from Kimchi via stimulation of macrophage cell line J774A. 1 in vitro. J. Food Sci. 2011, 76, H55–H61. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | LA (μg/mL) | CLA Productivity | Conversion Rate (%) | Isomer Ratio | ||||
|---|---|---|---|---|---|---|---|---|
| cis9, trans11-CLA (μg/mL) | Conversion Rate (%) | trans10, cis12-CLA (μg/mL) | Conversion Rate (%) | Total CLA (μg/mL) | ||||
| L. paraplantarum JBNU105611 | 100 | 91.5 ± 1.9 | 91.5 | 91.1 ± 3.6 | 91.1 | 182.6 ± 5.5 | 182.6 | 50:50 |
| 200 | 87.4 ± 3.8 | 43.7 | 87.3 ± 1.8 | 43.6 | 174.7 ± 5.6 | 87.3 | 50:50 | |
| 600 | 80.6 ± 5.8 | 13.4 | 77.1 ± 5.4 | 12.9 | 157.7 ± 11.1 | 26.3 | 51:49 | |
| 1000 | 75.2 ± 5.8 | 7.5 | 73.3 ± 0.0 | 7.3 | 148.5 ± 5.8 | 14.8 | 51:49 | |
| L. paraplantarum JBNU105634 | 100 | 73.8 ± 0.0 | 73.8 | 75.8 ± 0.0 | 75.9 | 149.7 ± 0.0 | 149.7 | 49:51 |
| 200 | 77.9 ± 1.9 | 39.0 | 77.1 ± 1.8 | 38.6 | 155.0 ± 3.7 | 77.5 | 50:50 | |
| 600 | 76.5 ± 0.0 | 12.8 | 67.5 ± 1.8 | 11.3 | 146.0 ± 1.8 | 24.3 | 52:48 | |
| 1000 | 76.5 ± 3.8 | 7.7 | 74.6 ± 5.4 | 7.5 | 151.1 ± 9.2 | 15.1 | 51:49 | |
| L. paraplantarum JBNU105655 | 100 | 163.4 ± 3.8 | 163.4 | 86.0 ± 0.0 | 86.0 | 249.4 ± 3.8 | 249.4 | 66:34 |
| 200 | 308.6 ± 1.9 | 154.3 | 89.8 ± 1.8 | 44.9 | 398.4 ± 0.1 | 199.2 | 77:23 | |
| 600 | 228.5 ± 3.8 | 38.1 | 80.9 ± 3.6 | 13.5 | 309.5 ± 7.4 | 51.6 | 74:26 | |
| 1000 | 140.3 ± 5.8 | 14.0 | 82.2 ± 5.4 | 8.2 | 222.5 ± 11.1 | 22.3 | 63:37 | |
| L. pentosus JBNU105674 | 100 | 231.3 ± 7.7 | 231.3 | 73.3 ± 0.0 | 73.3 | 304.6 ± 7.7 | 304.6 | 76:24 |
| 200 | 354.8 ± 9.6 | 177.4 | 80.9 ± 3.6 | 40.5 | 435.7 ± 13.2 | 217.8 | 81:19 | |
| 600 | 516.3 ± 7.7 | 86.0 | 80.9 ± 0.0 | 13.5 | 597.2 ± 7.7 | 99.5 | 86:14 | |
| 1000 | 182.4 ± 11.5 | 18.2 | 51.7 ± 1.8 | 5.2 | 234.1 ± 13.3 | 23.4 | 78:22 | |
| L. pentosus JBNU105676 | 100 | 261.1 ± 11.5 | 261.1 | 75.8 ± 3.6 | 75.9 | 337.0 ± 15.1 | 337.0 | 77:23 |
| 200 | 519.0 ± 11.5 | 259.5 | 92.4 ± 1.8 | 46.2 | 611.3 ± 13.3 | 305.7 | 85:15 | |
| 600 | 638.4 ± 3.8 | 106.4 | 88.6 ± 0.0 | 14.8 | 727.0 ± 3.8 | 121.2 | 88:12 | |
| 1000 | 357.5 ± 5.8 | 35.8 | 72.0 ± 1.8 | 7.2 | 429.5 ± 7.6 | 43.0 | 83:17 | |
| L. plantarum JBNU105645 | 100 | 117.3 ± 7.7 | 117.3 | 110.1 ± 5.4 | 110.2 | 227.4 ± 13.1 | 227.4 | 52:48 |
| 200 | 96.9 ±61.9 | 48.5 | 88.6 ± 3.6 | 44.3 | 185.4 ± 5.5 | 92.7 | 52:48 | |
| 600 | 101.0 ± 0.0 | 16.8 | 91.1 ± 0.0 | 15.2 | 192.1 ± 0.0 | 32.0 | 53:47 | |
| 1000 | 90.1 ± 7.7 | 9.0 | 77.1 ± 5.4 | 7.7 | 167.2 ± 13.1 | 16.7 | 54:46 | |
| L. plantarum JBNU105675 | 100 | 266.5 ± 7.7 | 266.5 | 89.8 ± 1.8 | 89.8 | 356.4 ± 9.5 | 356.4 | 75:25 |
| 200 | 361.5 ± 11.5 | 180.8 | 65.7 ± 3.6 | 32.8 | 427.2 ± 15.1 | 213.6 | 85:15 | |
| 600 | 185.1 ± 11.5 | 30.9 | 32.7 ± 0.0 | 5.4 | 217.8 ± 11.5 | 36.3 | 85:15 | |
| 1000 | 42.6 ± 1.9 | 4.3 | 27.6 ± 0.0 | 2.8 | 70.2 ± 2.0 | 7.0 | 61:39 | |
| L. plantarum JBNU105683 | 100 | 242.1 ± 0.0 | 242.1 | 92.4 ± 1.8 | 92.4 | 334.5 ± 1.8 | 334.5 | 72:28 |
| 200 | 443.0 ± 3.8 | 221.5 | 110.2 ± 1.8 | 55.1 | 553.1 ± 5.6 | 276.6 | 80:20 | |
| 600 | 642.5 ± 1.9 | 107.1 | 106.3 ± 0.0 | 17.7 | 748.8 ± 2.0 | 124.8 | 86:14 | |
| 1000 | 316.8 ± 1.9 | 31.7 | 96.2 ± 0.0 | 9.6 | 412.9 ± 2.0 | 41.3 | 77:23 | |
| L.mesenteroides JBNU105686 | 100 | 101.0 ± 0.0 | 101.0 | 73.3 ± 0.0 | 73.3 | 174.3 ± 0.0 | 174.3 | 58:42 |
| 200 | 90.1 ± 3.8 | 45.1 | 67.0 ± 1.8 | 33.5 | 157.1 ± 5.6 | 78.5 | 57:43 | |
| 600 | 83.3 ± 5.8 | 13.9 | 63.1 ± 3.6 | 10.5 | 146.5 ± 9.4 | 24.4 | 57:43 | |
| 1000 | 75.1 ± 9.6 | 7.5 | 60.6 ± 7.2 | 6.1 | 135.8 ± 16.8 | 13.6 | 55:45 | |
| P. acidilactici JBNU105117 | 100 | 73.8 ± 0.0 | 73.8 | 64.4 ± 1.8 | 64.4 | 138.2 ± 1.8 | 138.2 | 53:47 |
| 200 | 80.6 ± 1.9 | 40.3 | 65.2 ± 3.6 | 32.6 | 148.8 ± 5.5 | 74.4 | 54:46 | |
| 600 | 73.8 ± 0.0 | 12.3 | 63.1 ± 0.0 | 10.5 | 137.0 ± 0.0 | 22.8 | 54:46 | |
| 1000 | 76.5 ± 0.0 | 7.7 | 64.4 ± 1.8 | 6.4 | 141.0 ± 1.8 | 14.1 | 54:46 | |
| Strain | Viable Cell Counts (log CFU/mL) | |
|---|---|---|
| Control | 2 h | |
| L. paraplantarum JBCC105611 | 8.91 ± 0.10 | 4.68 ± 0.24 |
| P. acidilactici JBCC105117 | 9.12 ± 0.10 | 3.61 ± 0.16 |
| L. plantarum JBCC105675 | 9.01 ± 0.04 | 4.02 ± 0.15 |
| L. paraplantarum JBCC105634 | 7.88 ± 0.10 | 5.01 ± 0.53 |
| L. pentosus JBCC105676 | 9.11 ± 0.21 | 4.31 ± 0.19 |
| L. pentosus JBCC105674 | 9.05 ± 0.26 | 5.37 ± 0.22 |
| L. plantarum JBCC105683 | 9.05 ± 0.04 | 5.03 ± 0.44 |
| L. paraplantarum JBCC105655 | 8.97 ± 0.18 | 5.01 ± 0.51 |
| L. plantarum JBCC105645 | 9.08 ± 0.04 | 4.99 ± 0.40 |
| L. mesenteroides JBCC105686 | 8.77 ± 0.37 | 3.75 ± 0.19 |
| Strain | Bile Concentration (%) | ||||
|---|---|---|---|---|---|
| 0 | 0.3 | 1 | 3 | 5 | |
| L. paraplantarum JBNU105611 | 8.91 ± 0.10 a | 8.89 ± 0.02 a | 8.02 ± 0.18 b | 7.16 ± 0.22 c | 7.08 ± 0.05 c |
| P. acidilactici JBNU105117 | 9.12 ± 0.10 a | 9.03 ± 0.12 ab | 9.0 ± 0.11 ab | 8.19 ± 0.05 c | 7.89 ± 0.37 c |
| L. plantarum JBNU105675 | 9.01 ± 0.04 a | 8.99 ± 0.06 b | 8.83 ± 0.04 c | 6.70 ± 0.13 d | 6.62 ± 0.07 d |
| L. paraplantarum JBNU105634 | 7.88 ± 0.10 a | 7.81 ± 0.28 ab | 7.77 ± 0.13 b | 7.00 ± 0.18 c | 5.97 ± 0.21 d |
| L. pentosus JBNU105676 | 9.11 ± 0.21 a | 8.93 ± 0.07 ab | 8.76 ± 0.01 c | 8.66 ± 0.11 c | 7.51 ± 0.22 d |
| L. pentosus JBNU105674 | 9.05 ± 0.26 a | 8.89 ± 0.13 b | 8.17 ± 0.23 c | 5.74 ± 0.17 d | 5.35 ± 0.09 e |
| L. plantarum JBNU105683 | 9.05 ± 0.04 a | 9.01 ± 0.08 a | 8.37 ± 0.21 b | 6.44 ± 0.29 c | 6.15 ± 0.29 c |
| L. paraplantarum JBNU105655 | 8.97 ± 0.18 a | 8.07 ± 0.09 b | 6.68 ± 0.27 c | 6.13 ± 0.05 d | 5.17 ± 0.15 e |
| L. plantarum JBNU105645 | 9.08 ± 0.04 a | 7.9 ± 0.23 b | 7.03 ± 0.08 c | 7.12 ± 0.08 c | 6.69 ± 0.17 c |
| L. mesenteroides JBNU105686 | 8.77 ± 0.37 a | 8.55 ± 0.05 a | 7.61 ± 0.11 b | 7.02 ± 0.6 c | 6.218 ± 0.09 d |
| Strain | Pathogenic Microorganisms | ||||||
|---|---|---|---|---|---|---|---|
| S. aureus | S. epidermidis | S. xylosus | P. aeruginosa | P. putida | B. cereus | E. coli | |
| L. rhamnosus GG * | ++++ | + | +++ | ++ | ++ | ++ | + |
| L. paraplantarum JBNU105611 | ++++ | ++ | +++ | +++ | +++ | ++++ | ++++ |
| P. acidilactici JBNU105117 | ++++ | +++ | +++ | +++ | +++ | ++++ | ++++ |
| L. plantarum JBNU105675 | ++++ | + | +++ | ++ | +++ | ++++ | +++ |
| L. paraplantarum JBNU105634 | ++++ | ++ | +++ | ++ | ++ | +++ | +++ |
| L. pentosus JBNU105676 | ++++ | ++ | ++++ | ++ | +++ | ++++ | ++++ |
| L. pentosus JBNU105674 | ++++ | ++ | +++ | ++ | +++ | ++++ | ++ |
| L. plantarum JBNU105683 | ++++ | + | +++ | ++ | +++ | ++++ | +++ |
| L. paraplantarum JBNU105655 | ++++ | + | +++ | ++ | +++ | ++++ | ++++ |
| L. plantarum JBNU105645 | ++++ | + | +++ | ++ | +++ | ++++ | +++ |
| L. mesenteroides JBNU105686 | ++++ | ++ | +++ | +++ | ++ | +++ | ++++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, N.-E.; Kim, N.-J.; Kim, Y.-H.; Baik, S.-H. Probiotic Properties of Lactic Acid Bacteria with High Conjugated Linoleic Acid Converting Activity Isolated from Jeot-Gal, High-Salt Fermented Seafood. Microorganisms 2021, 9, 2247. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112247
Song N-E, Kim N-J, Kim Y-H, Baik S-H. Probiotic Properties of Lactic Acid Bacteria with High Conjugated Linoleic Acid Converting Activity Isolated from Jeot-Gal, High-Salt Fermented Seafood. Microorganisms. 2021; 9(11):2247. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112247
Chicago/Turabian StyleSong, Nho-Eul, Na-Jeong Kim, Young-Hun Kim, and Sang-Ho Baik. 2021. "Probiotic Properties of Lactic Acid Bacteria with High Conjugated Linoleic Acid Converting Activity Isolated from Jeot-Gal, High-Salt Fermented Seafood" Microorganisms 9, no. 11: 2247. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112247