
Extensive Genome Exploration of Clostridium botulinum Group III Field Strains

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Growth Condition, DNA Isolation and WGS
2.3. Genome Assembly
2.4. Phylogenetic Analysis
2.5. Bont and FliC Characterization
2.6. Plasmidome Analysis
2.7. Recombination Analysis
2.8. Flagellin Locus Sequencing
2.9. Nucleotide Sequence Accession Number
3. Results
3.1. Sequenced Genomes
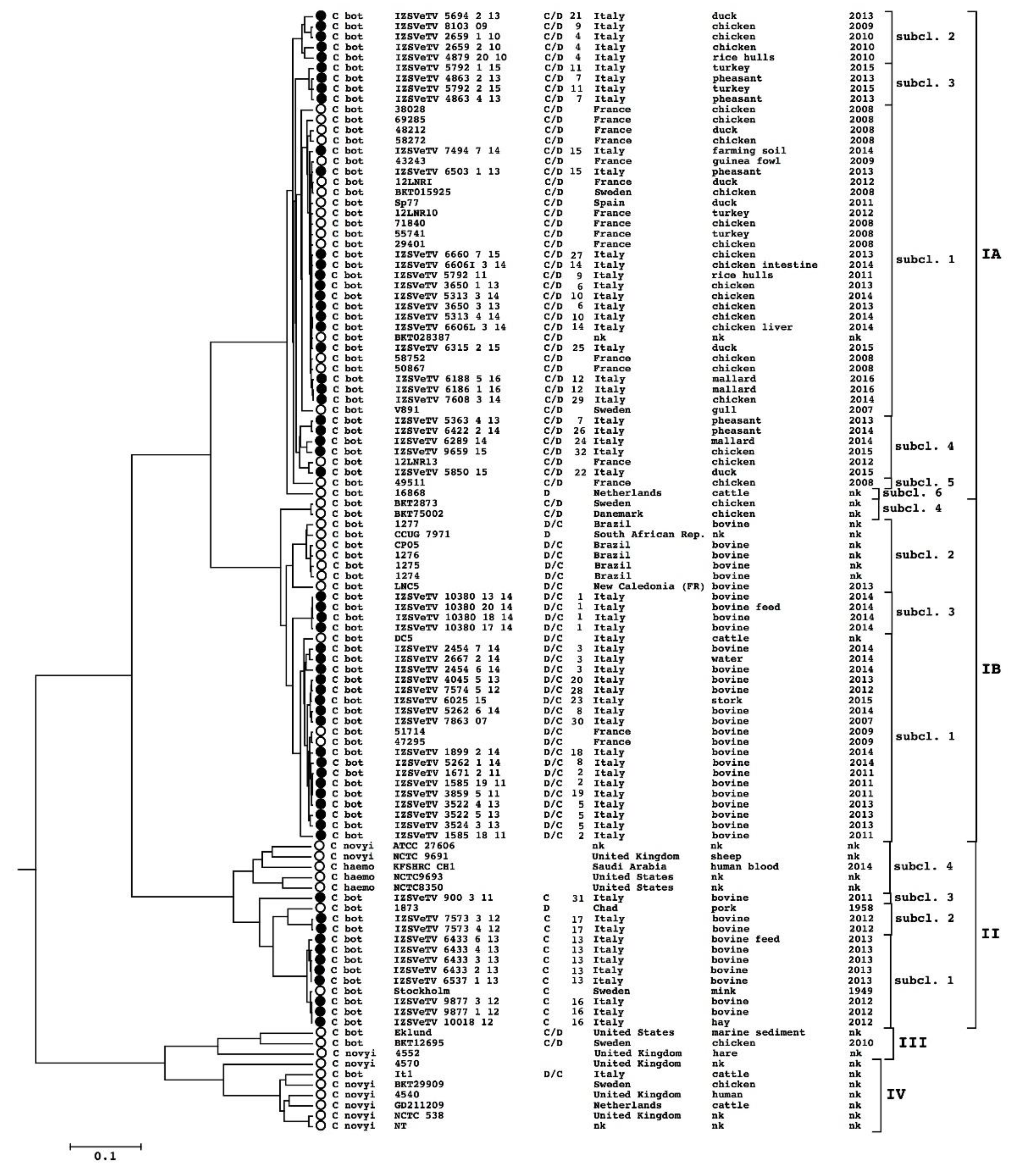
3.2. Genome Clustering
3.3. BoNT-Gene-Carrying Plasmid Clustering
3.4. BoNT Gene Sequence Comparison
3.5. Plasmidome Comparison
3.6. Recombination Analysis
3.7. fliC Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandez-Garayzabal, J.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A. The phylogeny of the genus Clostridium: Proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.; Williamson, C.H.D.; Hill, K.; Sahl, J.; Keim, P. Botulinum Neurotoxin-Producing Bacteria. Isn’t It Time that We Called a Species a Species? mBio 2018, 9, e01469-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatheway, C.L. Toxigenic Clostridia. Clin. Microbiol. Rev. 1990, 3, 66–98. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.R. Ecology of neurotoxigenic strains of clostridia. Curr. Top Microbiol. Immunol. 1995, 195, 1–29. [Google Scholar] [PubMed]
- Popoff, M.R.; Bouvet, P. Genetic characteristics of toxigenic Clostridia and toxin gene evolution. Toxicon 2013, 75, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Arnon, S.S.; Schechter, R.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Hauer, J.; Layton, M.; et al. Botulinum toxin as a biological weapon: Medical and public health management. JAMA 2001, 285, 1059–1070. [Google Scholar] [CrossRef]
- Pirazzini, M.; Azarnia Tehran, D.; Zanetti, G.; Megighian, A.; Scorzeto, M.; Fillo, S.; Shone, C.C.; Binz, T.; Rossetto, O.; Lista F Montecucco, C. Thioredoxin and its reductase are present on synaptic vesicles, and their inhibition prevents the paralysis induced by botulinum neurotoxins. Cell Rep. 2014, 8, 1870–1878. [Google Scholar] [CrossRef]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef]
- Hill, K.K.; Xie, G.; Foley, B.T.; Smith, T.J.; Munk, A.C.; Bruce, D.; Smith, L.A.; Brettin, T.S.; Detter, J.C. Recombination and insertion events involving the botulinum neurotoxin complex genes in Clostridium botulinum type A, B, E and F and Clostridium butyricum type E strains. BMC Biol. 2009, 7, 66. [Google Scholar] [CrossRef]
- Peck, M.W. Biology and genomic analysis of Clostridium botulinum. Adv. Microb. Physiol. 2009, 55, 183–265. [Google Scholar]
- Peck, M.W.; Stringer, S.C.; Carter, A.T. Clostridium botulinum in post-genomic era. Food Microbiol. 2011, 28, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Hill, K.K.; Raphael, B.H. Historical and current perspectives on Clostridium botulinum diversity. Res. Microbiol. 2015, 166, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Woudstra, C.; Le Maréchal, C.; Souillard, R.; Bayon-Auboyer, M.H.; Anniballi, F.; Auricchio, B.; De Medici, D.; Bano, L.; Koene, M.; Sansonetti, M.H.; et al. Molecular gene profiling of Clostridium botulinum group III and its detection in naturally contaminated samples originating from various European countries. Appl. Environ. Microbiol. 2015, 81, 2495–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, L.; Drigo, I.; Tonon, E.; Pascoletti, S.; Puiatti, C.; Anniballi, F.; Auricchio, B.; Lista, F.; Montecucco, C.; Agnoletti, F. Identification and characterization of Clostridium botulinum group III field strains by matrix-assisted laser desorption-ionization time-of-flight mass spectrometry (MALDI-TOF MS). Anaerobe 2017, 48, 126–134. [Google Scholar] [CrossRef]
- Skarin, H.; Hafstrom, T.; Westerberg, J.; Segerman, B. Clostridium botulinum group III: A group with dual identity shaped by plasmids, phages and mobile elements. BMC Genom. 2011, 12, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skarin, H.; Segerman, B. Plasmidome interchange between Clostridium botulinum, Clostridium novyi and Clostridium haemolyticum converts strains of independent lineages into distinctly different pathogens. PLoS ONE 2014, 9, e107777. [Google Scholar]
- Moriishi, K.; Koura, M.; Abe, N.; Fujii, N.; Fujinaga, Y.; Inoue, K.; Ogumad, K. Mosaic structures of neurotoxins produced from Clostridium botulinum types C and D organisms. Biochim. Biophys. Acta 1996, 1307, 123–126. [Google Scholar] [CrossRef]
- Nakamura, K.; Kohda, T.; Umeda, K.; Yamamoto, H.; Mukamoto, M.; Kozaki, S. Characterization of the D/C mosaic neurotoxin produced by Clostridium botulinum associated with bovine botulism in Japan. Vet. Microbiol. 2010, 140, 147–154. [Google Scholar] [CrossRef]
- Takeda, M.; Tsukamoto, K.; Kohda, T.; Matsui, M.; Mukamoto, M.; Kozaki, S. Characterization of the neurotoxin produced by isolates associated with avian botulism. Avian Dis. 2005, 49, 376–381. [Google Scholar] [CrossRef]
- Holdeman, L.V. The ecology and natural history of Clostridium botulinum. J. Wildl. Dis. 1970, 6, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Martrenchar, A.; Djossou, F.; Stagnetto, C.; Dupuy, C.; Brulez, E.; Attica, C.; Egmann, G.; Gruenfeld, J.; Fontanella, J.; Popoff, M. Is botulism type C transmissible to human by consumption of contaminated poultry meat? Analysis of a suspect outbreak in French Guyana. Anaerobe 2019, 56, 49–50. [Google Scholar] [CrossRef]
- Rasetti-Escargueil, C.; Lemichez, E.; Popoff, M.R. Public Health Risk Associated with Botulism as Foodborne Zoonoses. Toxins 2019, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Woudstra, C.; Skarin, H.; Anniballi, F.; Fenicia, L.; Bano, L.; Drigo, I.; Koene, M.; Bäyon-Auboyer, M.H.; Buffereau, J.P.; De Medici, D.; et al. Neurotoxin gene profiling of clostridium botulinum types C and D native to different countries within Europe. Appl. Environ. Microbiol. 2012, 78, 3120–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, L.; Drigo, I.; Tonon, E.; Agnoletti, F.; Giovanardi, D.; Morandini, E. Rice hulls as a possible source of Clostridium botulinum type C spores for poultry. Vet. Rec. 2013, 173, 427. [Google Scholar] [CrossRef] [PubMed]
- Anza, I.; Skarin, H.; Vidal, D.; Lindberg, A.; Baverud, V.; Mateo, R. The same clade of Clostridium botulinum strains is causing avian botulism in southern and northern Europe. Anaerobe 2014, 26, 20–23. [Google Scholar] [CrossRef]
- Woudstra, C.; Tevell Åberg, A.; Skarin, H.; Anniballi, F.; De Medici, D.; Bano, L.; Koene, M.; Löfström, C.; Hansen, T.; Hedeland, M.; et al. Animal botulism outcomes in the AniBioThreat project. Biosecur. Bioterror. 2013, 11 (Suppl. 1), S177–S182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, L.; Drigo, I.; Tonon, E.; Berto, G.; Tavella, A.; Woudstra, C.; Capello, K.; Agnoletti, F. Evidence for a natural humoral response in dairy cattle affected by persistent botulism sustained by non-chimeric type C strains. Anaerobe 2015, 36, 25–29. [Google Scholar] [CrossRef]
- Abbitt, B.; Murphy, M.J.; Ray, A.C.; Reagor, J.C.; Eugster, A.K.; Gayle, L.G.; Whitford, H.W.; Sutherland, R.J.; Fiske, R.A.; Pusok, J. Catastrophic death losses in a dairy herd attributed to type D botulism. J. Am. Vet. Med. Assoc. 1984, 185, 798–801. [Google Scholar]
- Jean, D.; Fecteau, G.; Scott, D.; Higgins, R.; Quessy, S. Clostridium botulinum type C intoxication in feedlot steers being fed ensiled poultry litter. Can. Vet. J. 1995, 36, 626–628. [Google Scholar]
- AFSSA (Agence Française de Sécurité Sanitaire des Aliments) Rapport sur le Botulisme D’origine Aviaire et Bovine. 10/2002, 82. Maisons-Alfort, France. Available online: https://documentation.ehesp.fr/index.php?lvl=notice_display&id=179676 (accessed on 23 September 2021).
- Carlier, J.P.; Popoff, M.R. Situation actuelle du botulisme bovin et aviaire en France. Bull. l’Académie Vétérinaire Fr. 2002, 155, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Braun, U.; Feige, K.; Schweizer, G.; Pospischil, A. Clinical findings and treatment of 30 cattle with botulism. Vet. Rec. 2005, 156, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Myllykoski, J.L.; Lindström, M.; Keto-Timonen, R.; Söderholm, H.; Jakala, J.; Kallio, H.; Sukura, A.; Korkeala, H. Type C bovine botulism outbreak due to carcass contaminated non-acidified silage. Epidemiol. Infect. 2009, 137, 284–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souillard, R.; Woudstra, C.; Le Marechal, C.; Dia, M.; Bayon-Auboyer, M.H.; Chemaly, M.; Fach, P.; Le Bouquin, S. Investigation of Clostridium botulinum in commercial poultry farms in France between 2011 and 2013. Avian Pathol. 2014, 43, 458–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyboldt, C.; Discher, S.; Jordan, E.; Neubauer, H.; Jensen, K.C.; Campe, A.; Kreienbrock, L.; Scheu, T.; Wichern, A.; Gundling, F.; et al. Occurrence of Clostridium botulinum neurotoxin in chronic disease of dairy cows. Vet. Microbiol. 2015, 177, 398–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relun, A.; Dorso, L.; Douart, A.; Chartier, C.; Guatteo, R.; Mazuet, C.; Popoff, M.R.; Assié, S. A large outbreak of bovine botulism possibly linked to a massive contamination of grass silage by type D/C Clostridium botulinum spores on a farm with dairy and poultry operations. Epidemiol. Infect. 2017, 145, 3477–3485. [Google Scholar] [CrossRef] [Green Version]
- Yeh, J.Y.; Seo, H.J.; Cho, Y.S.; Park, J.Y.; Cho, Y.S.; Cho, I.S.; Lee, J.H.; Hwang, J.M.; Choi, I.S. Livestock agroterrorism: The deliberate introduction of a highly infectious animal pathogen. Foodborne Pathog. Dis. 2012, 9, 869–877. [Google Scholar] [CrossRef]
- Anniballi, F.; Fiore, A.; Löfström, C.; Skarin, H.; Auricchio, B.; Woudstra, C.; Bano, L.; Segerman, B.; Koene, M.; Båverud, V.; et al. Management of animal botulism outbreaks: From clinical suspicion to practical countermeasures to prevent or minimize outbreaks. Biosecur. Bioterror. 2013, 11 (Suppl. 1), S191–S199. [Google Scholar] [CrossRef] [Green Version]
- Skarin, H.; Tevell Åberg, A.; Woudstra, C.; Hansen, T.; Löfström, C.; Koene, M.; Bano, L.; Hedeland, M.; Anniballi, F.; De Medici, D.; et al. The workshop on animal botulism in Europe. Biosecur. Bioterror. 2013, 11 (Suppl. 1), S183–S190. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.K.; Smith, T.J.; Helma, C.H.; Ticknor, L.O.; Foley, B.T.; Svensson, R.T.; Brown, J.L.; Johnson, E.A.; Smith, L.A.; Okinaka, R.T.; et al. Genetic diversity among Botulinum Neurotoxin-producing clostridial strains. J. Bacteriol. 2007, 189, 818–832. [Google Scholar] [CrossRef] [Green Version]
- Skarin, H.; Lindberg, A.; Blomqvist, G.; Aspan, A.; Baverud, V. Molecular characterization and comparison of Clostridium botulinum type C avian strains. Avian Pathol. 2010, 39, 511–518. [Google Scholar] [CrossRef]
- Skarin, H.; Segerman, B. Horizontal gene transfer of toxin genes in Clostridium botulinum: Involvement of mobile elements and plasmids. Mob. Genet. Elem. 2011, 1, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Woudstra, C.; Le Maréchal, C.; Souillard, R.; Bayon-Auboyer, M.H.; Mermoud, I.; Desoutter, D.; Fach, P. New Insights into the Genetic Diversity of Clostridium botulinum Group III through Extensive Genome Exploration. Front. Microbiol. 2016, 7, 757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eklund, M.W.; Poysky, F.T.; Meyers, J.A.; Pelroy, G.A. Interspecies conversion of Clostridium botulinum type C to Clostridium novyi type A by bacteriophage. Science 1974, 186, 456. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.T.; Paul, C.J.; Mason, D.R.; Twine, S.M.; Alston, M.J.; Logan, S.M.; Austin, J.W.; Peck, M.W. Independent evolution of neurotoxin and flagellar genetic loci in proteolytic Clostridium botulinum. BMC Genom. 2009, 10, 115. [Google Scholar] [CrossRef] [Green Version]
- Woudstra, C.; Lambert, D.; Anniballi, F.; De Medici, D.; Austin, J.; Fach, P. Genetic diversity of the flagellin genes of Clostridium botulinum groups I and II. Appl. Environ. Microbiol. 2013, 79, 3926–3932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardella, M.A.; Duff, J.T.; Gottfried, C.; Begel, J.S. Studies on immunity to toxins of Clostridium botulinum. IV. Production and purification of type C toxin for conversion to toxoid. J. Bacteriol. 1958, 75, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Del Fabbro, C.; Scalabrin, S.; Morgante, M.; Giorgi, F.M. An Extensive Evaluation of Read Trimming Effects on Illumina NGS Data Analysis. PLoS ONE 2013, 8, e85024. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Agren, J.; Sundstrom, A.; Hafstrom, T.; Segerman, B. Gegenees: Fragmented alignment of multiple genomes for determining phylogenomic distances and genetic signatures unique for specified targe groups. PLoS ONE 2012, 7, e39107. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Vallvé, S.; Palau, J.; Romeu, A. Horizontal gene transfer in glycosyl hydrolases inferred from codon usage in Escherichia coli and Bacillus subtilis. Mol. Biol. Evol. 1999, 16, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, S.N.; Slezak, T.; Hall, B.G. kSNP3.0: SNP detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinformatics 2015, 31, 2877–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. PlasmidSPAdes: Assembling plasmids from whole genome sequencing data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Didelot, X.; Falush, D. Inference of bacterial microevolution using multilocus sequence data. Genetics 2007, 175, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Didelot, X.; Lawson, D.; Darling, A.; Falush, D. Inference of homologous recombination in bacteria using whole-genome sequences. Genetics 2010, 186, 1435–1449. [Google Scholar] [CrossRef] [Green Version]
- Giordani, F.; Fillo, S.; Anselmo, A.; Palozzi, A.M.; Fortunato, A.; Gentile, B.; Azarnia Tehran, D.; Ciammaruconi, A.; Spagnolo, F.; Pittiglio, V.; et al. Genomic characterization of Italian Clostridium botulinum group I strains. Infect. Genet Evol. 2015, 36, 62–71. [Google Scholar] [CrossRef]
- Woudstra, C.; Brito, R.B.; Fonseca Júnior, A.A.; Silva, R.O.S.; Lobato, F.C.F.; Fach, P. Draft Genome Sequences of Five Brazilian Clostridium botulinum Group III Type D/C Strains. Genome Announc. 2017, 5, e00349-17. [Google Scholar] [CrossRef] [Green Version]
- Vos, M.; Didelot, X. A comparison of homologous recombination rates in bacteria and archaea. ISME J. 2009, 3, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Williamson, C.H.; Sahl, J.W.; Smith, T.J.; Xie, G.; Foley, B.T.; Smith, L.A.; Fernández, R.A.; Lindström, M.; Korkeala, H.; Keim, P.; et al. Comparative genomic analyses reveal broad diversity in botulinum-toxin-producing Clostridia. BMC Genom. 2016, 17, 180. [Google Scholar] [CrossRef] [Green Version]
- Weedmark, K.A.; Mabon, P.; Hayden, K.L.; Lambert, D.; Van Domselaar, G.; Austin, J.W.; Corbett, C.R. Clostridium botulinum Group II Isolate Phylogenomic Profiling Using Whole-Genome Sequence Data. Appl. Environ. Microbiol. 2015, 81, 5938–5948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, Y.; Hayashi, T.; Kurokawa, K.; Nakayama, K.; Oshima, K.; Fujinaga, Y.; Ohnishi, M.; Ohtsubo, E.; Hattori, M.; Oguma, K. The genome sequence of Clostridium botulinum type C neurotoxin-converting phage and the molecular mechanisms of unstable lysogeny. Proc. Natl. Acad. Sci. USA 2005, 29, 17472–17477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, M.W.; Smith, T.J.; Anniballi, F.; Austin, J.W.; Bano, L.; Bradshaw, M.; Cuervo, P.; Cheng, L.W.; Derman, Y.; Dorner, B.G.; et al. Historical perspectives and guidelines for botulinum neurotoxin subtype nomenclature. Toxins 2017, 9, 38. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Serotype | fliC Type | Collection Year | Collection Location | Isolation Source | Host | Outbreak ID |
|---|---|---|---|---|---|---|---|
| IZSVe-TV 10380/13/14 | D/C | VI-VII | 2014 | Cervignano del Friuli (UD)–Friuli VG | faeces | Bos taurus | 1 |
| IZSVe-TV 10380/17/14 | D/C | VI-VII | 2014 | Cervignano del Friuli (UD)–Friuli VG | rectum | Bos taurus | 1 |
| IZSVe-TV 10380/18/14 | D/C | VI-VII | 2014 | Cervignano del Friuli (UD)–Friuli VG | colon | Bos taurus | 1 |
| IZSVe-TV 10380/20/14 | D/C | VI-VII | 2014 | Cervignano del Friuli (UD)–Friuli VG | feed | 1 | |
| IZSVe-TV 1585/18/11 | D/C | IV | 2011 | Piove di Sacco (PD)–Veneto | ruminal content | Bos taurus | 2 |
| IZSVe-TV 1585/19/11 | D/C | IV | 2011 | Piove di Sacco (PD)–Veneto | ruminal content | Bos taurus | 2 |
| IZSVe-TV 1671/2/11 | D/C | IV | 2011 | Piove di Sacco (PD)–Veneto | ruminal content | Bos taurus | 2 |
| IZSVe-TV 2454/6/14 | D/C | IV | 2014 | Mansuè (TV)–Veneto | faeces | Bos taurus | 3 |
| IZSVe-TV 2454/7/14 | D/C | IV | 2014 | Mansuè (TV)–Veneto | faeces | Bos taurus | 3 |
| IZSVe-TV 2667/2/14 | D/C | IV | 2014 | Mansuè (TV)–Veneto | drinking water | 3 | |
| IZSVe-TV 2659/1/10 | C/D | I | 2010 | Calvenzano (BG)–Lombardia | intestine content | Gallus gallus | 4 |
| IZSVe-TV 2659/2/10 | C/D | I | 2010 | Calvenzano (BG)–Lombardia | intestine content | Gallus gallus | 4 |
| IZSVe-TV 4879B/20/10 | C/D | I | 2010 | Calvenzano (BG)–Lombardia | rice hulls (litter for chicken) | 4 | |
| IZSVe-TV 3522/4/13 | D/C | IV | 2013 | Trebaseleghe (PD)–Veneto | faeces | Bos taurus | 5 |
| IZSVe-TV 3522/5/13 | D/C | IV | 2013 | Trebaseleghe (PD)–Veneto | faeces | Bos taurus | 5 |
| IZSVe-TV 3524/3/13 | D/C | IV | 2013 | Trebaseleghe (PD)–Veneto | intestine content | Bos taurus | 5 |
| IZSVe-TV 3650/1/13 | C/D | I | 2013 | Caprino Veronese (VR)–Veneto | liver | Gallus gallus | 6 |
| IZSVe-TV 3650/3/13 | C/D | I | 2013 | Caprino Veronese (VR)–Veneto | intestine content | Gallus gallus | 6 |
| IZSVe-TV 4863/2/13 | C/D | I | 2013 | Castelvetro (PC)–Emilia Romagna | gastric content | Phasianus colchicus | 7 |
| IZSVe-TV 4863/4/13 | C/D | I | 2013 | Castelvetro (PC)–Emilia Romagna | gastric content | Phasianus colchicus | 7 |
| IZSVe-TV 5363/4/13 | C/D | I | 2013 | Castelvetro (PC)–Emilia Romagna | intestine content | Phasianus colchicus | 7 |
| IZSVe-TV 5262/1/14 | D/C | IV | 2014 | San Zenone degli Ezzelini (TV)–Veneto | faeces | Bos taurus | 8 |
| IZSVe-TV 5262/6/14 | D/C | IV | 2014 | San Zenone degli Ezzelini (TV)–Veneto | faeces | Bos taurus | 8 |
| IZSVe-TV 5792/11 | C/D | I | 2011 | Volta Mantovana (MN)–Lombardia | rice hulls (litter for chicken) | 9 | |
| IZSVe-TV 8103/09 | C/D | I | 2009 | Volta Mantovana (MN)–Lombardia | intestine content | Gallus gallus | 9 |
| IZSVe-TV 5313/3/14 | C/D | I | 2014 | Agugliaro (VI)–Veneto | intestine content | Gallus gallus | 10 |
| IZSVe-TV 5313/4/14 | C/D | I | 2014 | Agugliaro (VI)–Veneto | intestine content | Gallus gallus | 10 |
| IZSVe-TV 5792/1/15 | C/D | I | 2015 | Castelnovo Bariano (RO)–Veneto | liver | Meleagris gallopavo | 11 |
| IZSVe-TV 5792/2/15 | C/D | I | 2015 | Castelnovo Bariano (RO)–Veneto | intestine content | Meleagris gallopavo | 11 |
| IZSVe-TV 6186/1/16 | C/D | I | 2016 | Muggia (TS)–Friuli VG | intestine content | Anas platyrhynchos | 12 |
| IZSVe-TV 6188/5/16 | C/D | I | 2016 | Muggia (TS)–Friuli VG | intestine content | Anas platyrhynchos | 12 |
| IZSVe-TV 6433/2/13 | C | II | 2013 | Sarentino (BZ)–Trentino AA | faeces | Bos taurus | 13 |
| IZSVe-TV 6433/3/13 | C | II | 2013 | Sarentino (BZ)–Trentino AA | faeces | Bos taurus | 13 |
| IZSVe-TV 6433/4/13 | C | II | 2013 | Sarentino (BZ)–Trentino AA | faeces | Bos taurus | 13 |
| IZSVe-TV 6433/6/13 | C | II | 2013 | Sarentino (BZ)–Trentino AA | feed | 13 | |
| IZSVe-TV 6537/1/13 | C | II | 2013 | Sarentino (BZ)–Trentino AA | ruminal content | Bos taurus | 13 |
| IZSVe-TV 6606L/3/14 | C/D | I | 2014 | Naturno (BZ)–Trentino AA | liver | gallus gallus | 14 |
| IZSVe-TV 6606I/3/14 | C/D | I | 2014 | Naturno (BZ)–Trentino AA | intestine content | gallus gallus | 14 |
| IZSVe-TV 6503/1/13 | C/D | I | 2013 | Montalto di Castro (VT)–Lazio | liver | Phasianus colchicus | 15 |
| IZSVe-TV 7494/7/14 | C/D | I | 2014 | Montalto di Castro (VT)–Lazio | Soil (pheasant farm) | 15 | |
| IZSVe-TV 10018/12 | C | II | 2012 | Scena (BZ)–Trentino AA | hay | 16 | |
| IZSVe-TV 9877/1/12 | C | II | 2012 | Scena (BZ)–Trentino AA | faeces | Bos taurus | 16 |
| IZSVe-TV 9877/3/12 | C | II | 2012 | Scena (BZ)–Trentino AA | faeces | Bos taurus | 16 |
| IZSVe-TV 7573/3/12 | C | II | 2012 | San Lorenzo di Sebato (BZ)–Trentino AA | faeces | Bos taurus | 17 |
| IZSVe-TV 7573/4/12 | C | II | 2012 | San Lorenzo di Sebato (BZ)–Trentino AA | faeces | Bos taurus | 17 |
| IZSVe-TV 1899/2/14 | D/C | IV | 2014 | Levico Terme (TN)–Trentino AA | bovine ruminal content | Bos taurus | 18 |
| IZSVe-TV 3859/5/11 | D/C | IV | 2011 | Galgagnano (MI)–Lombardia | faeces | Bos taurus | 19 |
| IZSVe-TV 4045/5/13 | D/C | IV | 2013 | Bleggio Superiore (TN)–Trentino AA | liver | Bos taurus | 20 |
| IZSVe-TV 5694/2/13 | C/D | I | 2013 | Mogliano Veneto (TV)–Veneto | intestine content | Anas platyrhynchos | 21 |
| IZSVe-TV 5850/15 | C/D | I | 2015 | Correzzola (PD)–Veneto | intestine content | Anas platyrhynchos | 22 |
| IZSVe-TV 6025/15 | D/C | IV | 2015 | Quinto di Treviso (TV)–Veneto | intestine content | Ciconia ciconia | 23 |
| IZSVe-TV 6289/14 | C/D | I | 2014 | Lesina (FG)–Puglia | intestine content | Anas platyrhynchos | 24 |
| IZSVe-TV 6315/2/15 | C/D | I | 2015 | Perugia (PG)–Umbria | intestine content | Anas platyrhynchos | 25 |
| IZSVe-TV 6422/2/14 | C/D | I | 2014 | Cremona (CR)–Lombardia | intestine content | Phasianus colchicus | 26 |
| IZSVe-TV 6660/7/15 | C/D | I | 2015 | San Martino Buonalbergo (VR)–Veneto | intestine content | gallus gallus | 27 |
| IZSVe-TV 7574/5/12 | D/C | IV | 2012 | Stenico (TN)–Trentino AA | faeces | Bos taurus | 28 |
| IZSVe-TV 7608/3/14 | C/D | I | 2014 | Forlì (FC)–Emilia Romagna | intestine content | Gallus gallus | 29 |
| IZSVe-TV 7863/07 | D/C | IV | 2007 | Colle Umberto (TV)–Veneto | faeces | Gallus gallus | 30 |
| IZSVe-TV 900/3/11 | C | II | 2011 | Adria (RO)–Veneto | liver | Bos taurus | 31 |
| IZSVe-TV 9659/15 | C/D | I | 2015 | Polesella (RO)–Veneto | intestine content | Gallus gallus | 32 |
| Primer Name | Sequence 5′ → 3′ | Amplicon Size (bp) |
|---|---|---|
| fliC_VI_F1_f | TGCTGAAAAAGCAAGAATACAGAAAAG | 805 |
| fliC_VI_F1_r | TGTTGCTGCTTTATCTGCCTTTAAT | |
| fliC_VI_F2_f | AGGAGCTAATGAAGGTCAATCAATG | 907 |
| fliC_VI_F2_r | CCTTTTCTCACTTCTTTGAATGGGT | |
| fliC_VI_F3_f | CGATTTAGTTAATGGAACTGAATCCAC | 806 |
| fliC_VI_F3_r | TAAGGCACCTGAAGTCATATCATCC | |
| fliC_VI_F4_f | CAGAAGTTAATAGAATAGCTAATACAACTG | 981 |
| fliC_VI_F4_r | GCCAGAGATTTATCTCCGACTCTTA | |
| fliC_VI_F5_f | AGAGGACAAATCAGAGGTCTTAACC | 686 |
| fliC_VI_F5_r | CCAATTACTCCAGCTGCAGTAGT | |
| fliC_VI_F6_f | AGAGGACAAATCAGAGGTCTTAACC | 663 |
| fliC_VI_F6_r | GTCAGCATTCTTTATTTCAGTGGCT | |
| fliC_VI_F7_f | AATCAACAACCACAAGGAGTTCTTC | 539 |
| fliC_VI_F7_r | GGTTAAGACCTCTGATTTGTCCTCT |
| fliC Variants | ||||||
|---|---|---|---|---|---|---|
| fliC-I | / | 92.66 | 84.86 | 83.94 | 61.24 | 50.69 |
| fliC-III | 92.66 | / | 86.93 | 84.86 | 61.93 | 51.38 |
| fliC-II | 84.86 | 86.93 | / | 86.01 | 61.47 | 50.92 |
| fliC-V | 83.94 | 84.86 | 86.01 | / | 59.86 | 49.31 |
| fliC 10380 VII | 61.24 | 61.93 | 61.47 | 59.86 | / | 82.8 |
| fliC 10380 VI | 50.69 | 51.38 | 50.92 | 49.31 | 82.8 | / |
| fliC-IV | 46.79 | 47.48 | 47.48 | 47.25 | 60.78 | 57.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fillo, S.; Giordani, F.; Tonon, E.; Drigo, I.; Anselmo, A.; Fortunato, A.; Lista, F.; Bano, L. Extensive Genome Exploration of Clostridium botulinum Group III Field Strains. Microorganisms 2021, 9, 2347. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112347
Fillo S, Giordani F, Tonon E, Drigo I, Anselmo A, Fortunato A, Lista F, Bano L. Extensive Genome Exploration of Clostridium botulinum Group III Field Strains. Microorganisms. 2021; 9(11):2347. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112347
Chicago/Turabian StyleFillo, Silvia, Francesco Giordani, Elena Tonon, Ilenia Drigo, Anna Anselmo, Antonella Fortunato, Florigio Lista, and Luca Bano. 2021. "Extensive Genome Exploration of Clostridium botulinum Group III Field Strains" Microorganisms 9, no. 11: 2347. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112347