
Role of Maturation of Lipoproteins in the Pathogenesis of the Infection Caused by Streptococcus suis Serotype 2


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains and Growth Conditions
2.3. DNA Manipulations
2.4. Construction of the Lipoprotein Maturation Isogenic Mutants
2.5. Complementation of the Mutant Strains
2.6. Growth Analysis
2.7. Bacterial Surface Hydrophobicity Assay
2.8. Preparation of Heat-Killed S. suis
2.9. Preparation of Bacterial Supernatants
2.10. Porcine Tracheal Epithelial Cell Culture and Bacterial Adhesion and Invasion Assays
2.11. Whole Blood Bactericidal (Killing) Assay
2.12. Generation of Bone Marrow-Derived Dendritic Cells (bmDC)
2.13. S. suis Infection of bmDCs
2.14. S. suis Virulence Mouse Model of Systemic Infection
2.15. Measurement of Plasma (Systemic) Pro-Inflammatory Mediators
2.16. Statistical Analyses
3. Results
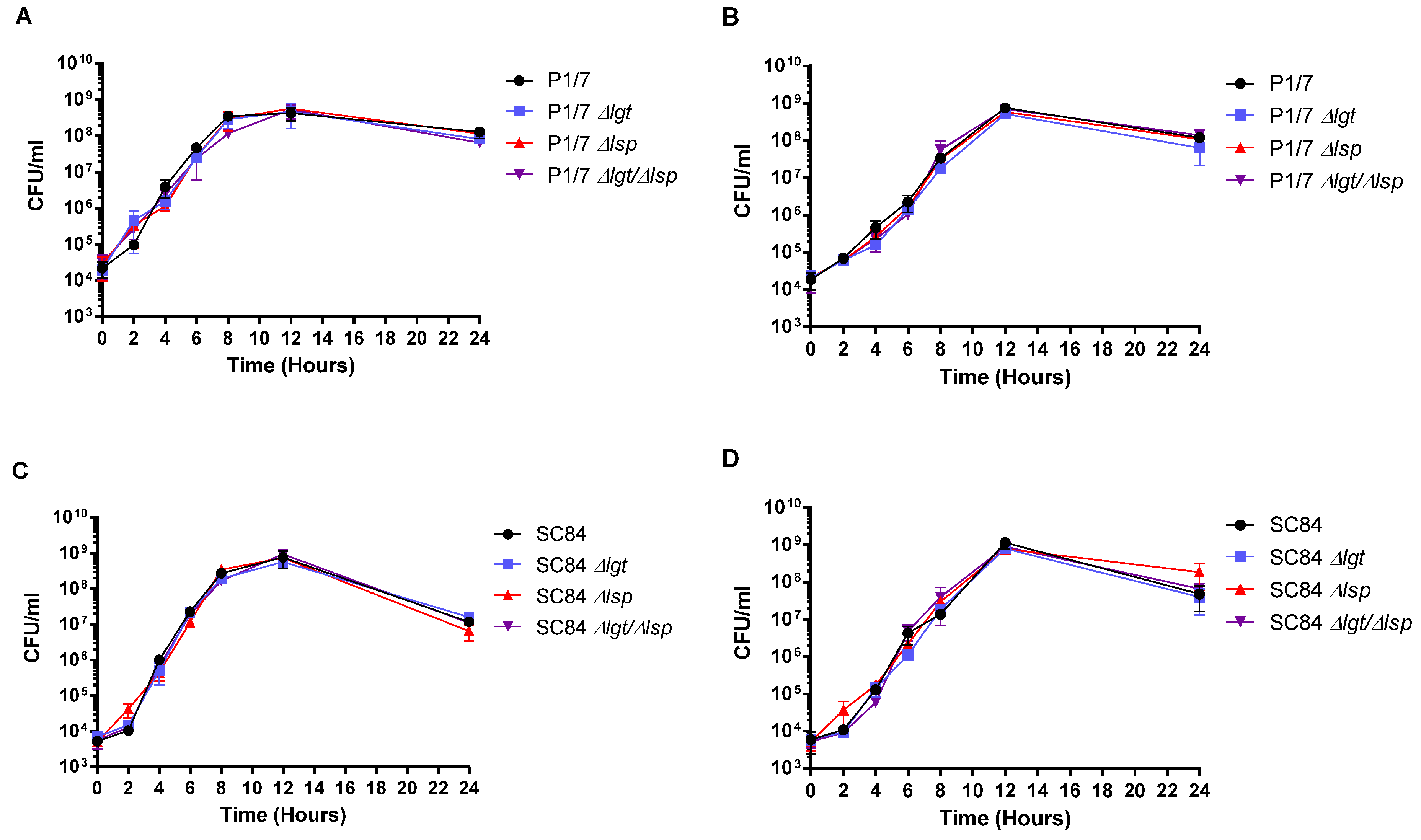
3.1. Characteristics of the Δlgt, Δlsp and Δlgt/Δlsp Mutants: Normal Growth in Rich Medium and in Plasma
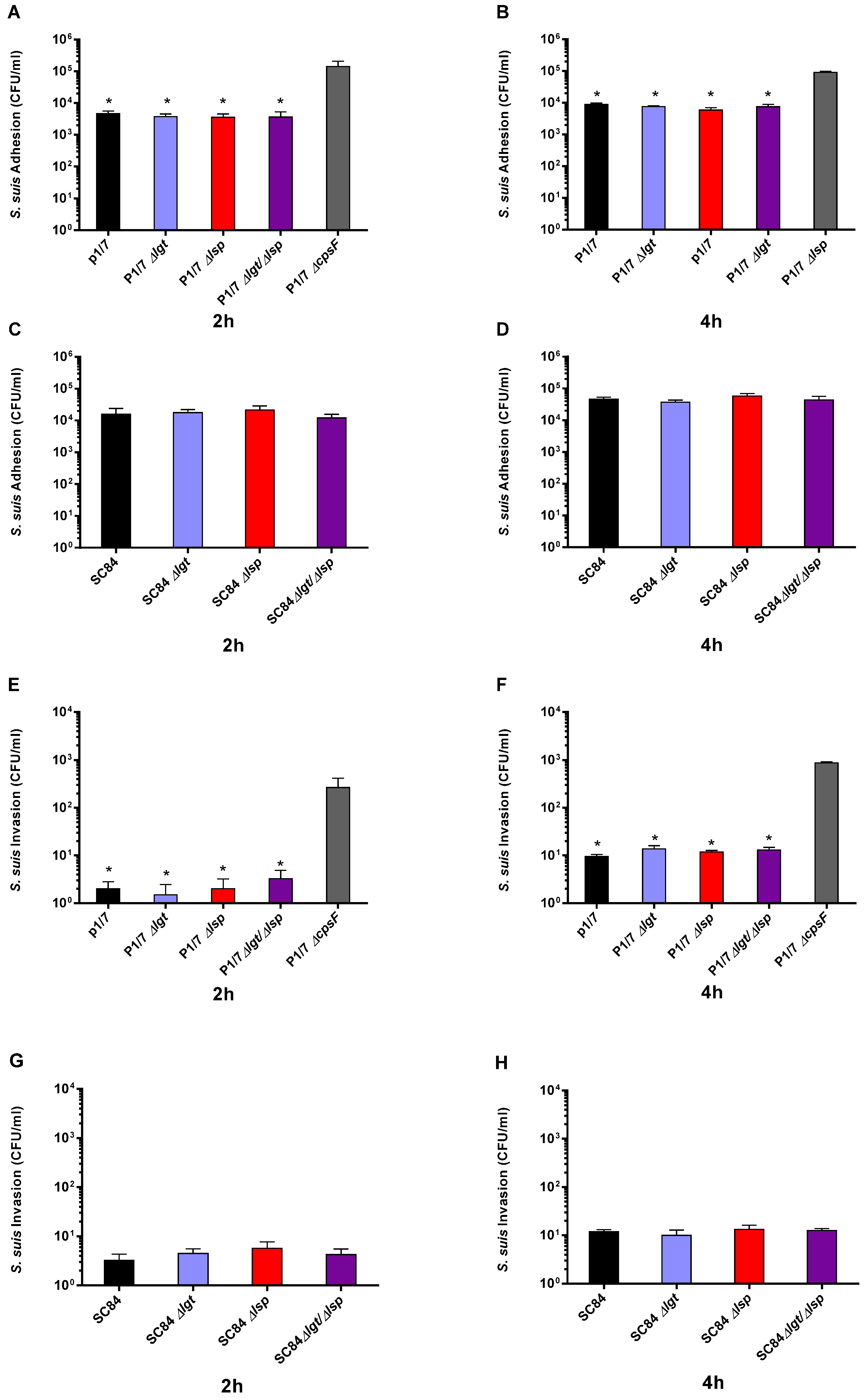
3.2. Absence of the Lipoprotein Maturation Enzymes Does Not Affect S. suis Serotype 2 Adhesion to and Invasion of Respiratory Epithelial Cells
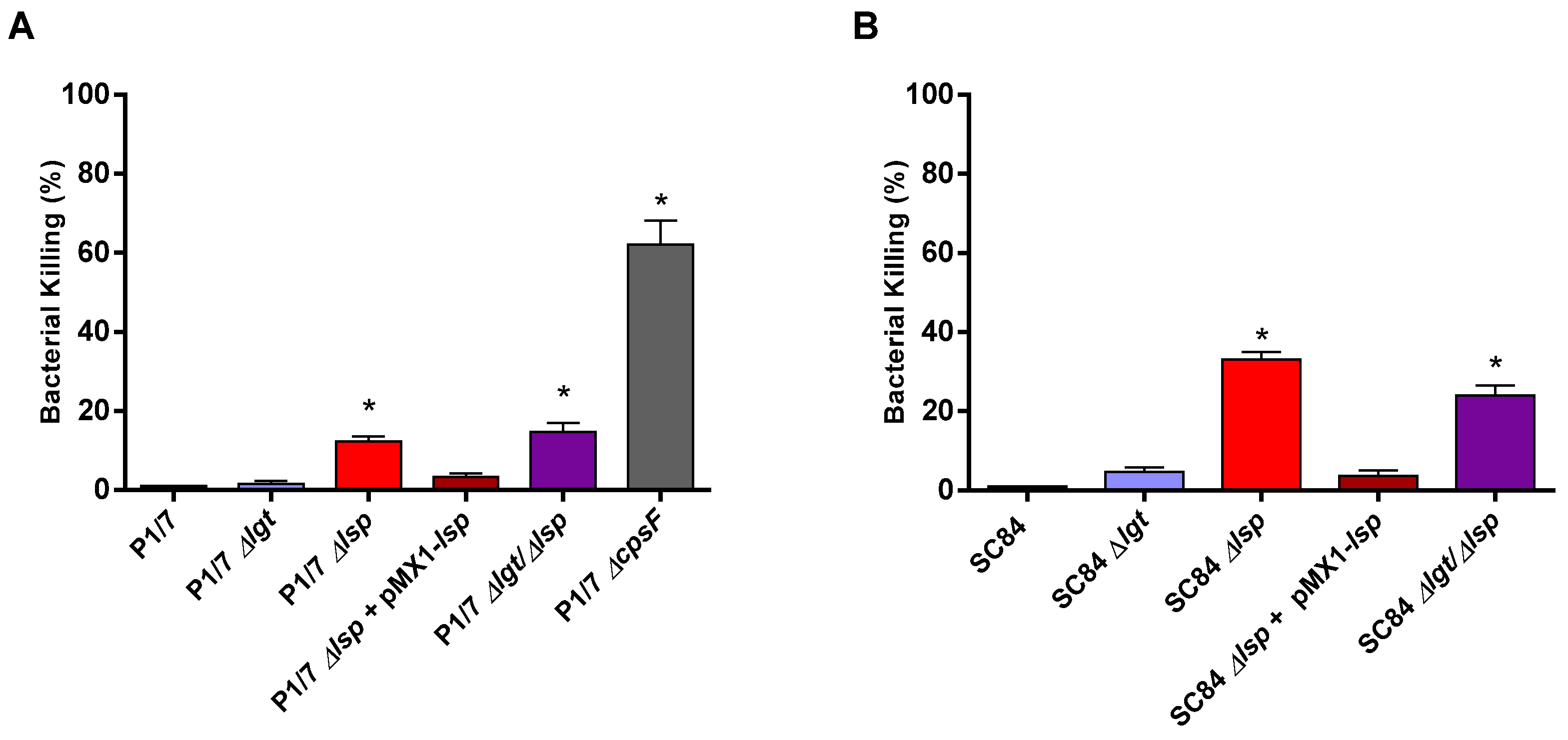
3.3. Presence of the Prolipoprotein Signal Peptidase Type II Is Partially Required for S. suis Serotype 2 In Vitro Resistance to Bacterial Killing in a Whole Blood Test
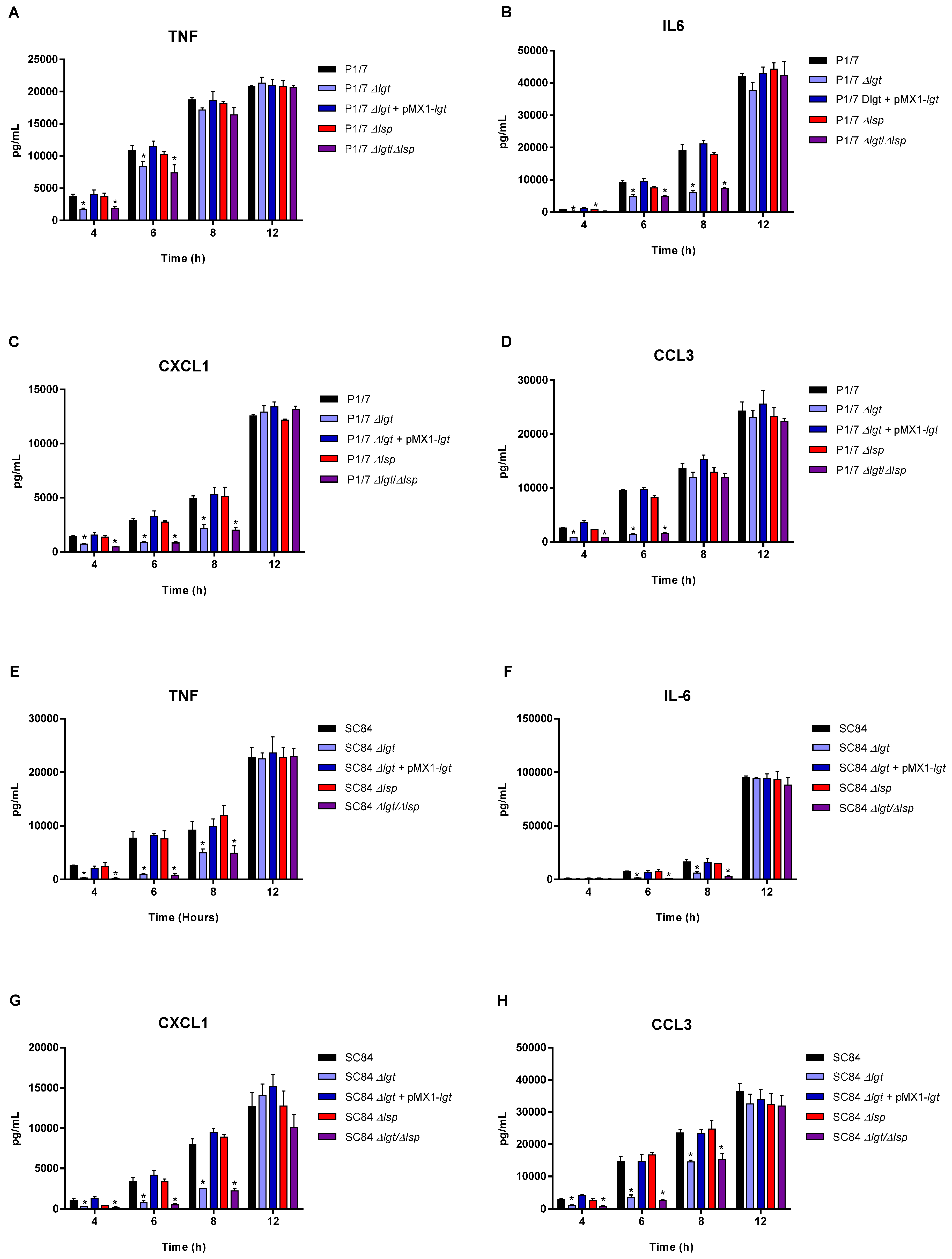
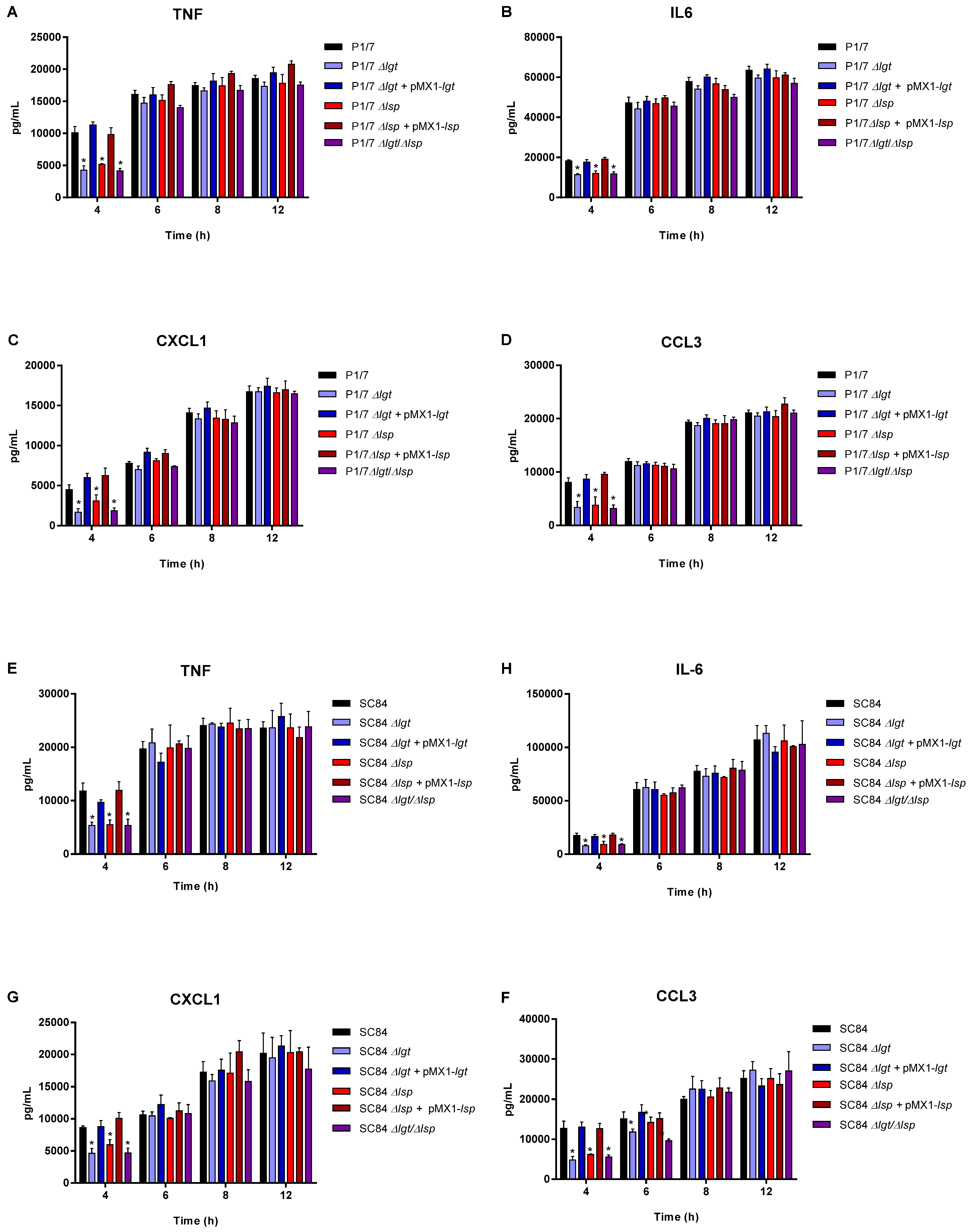
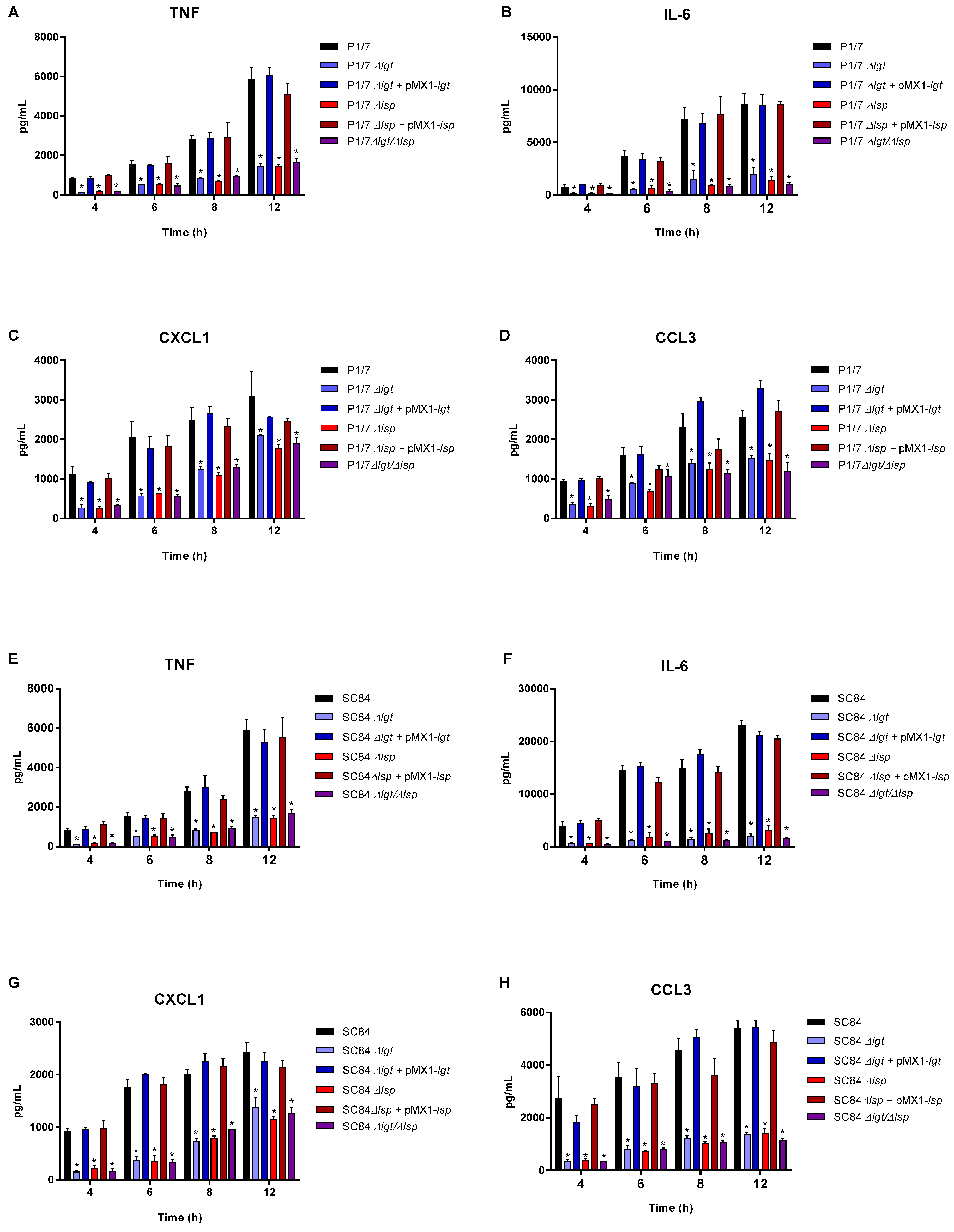
3.4. The Diacyl Motif and the Peptide Signal Cleavage Are Important for the Recognition of Periplasmic and/or Secreted S. suis Serotype 2 Lipoproteins by Innate Immune Cells
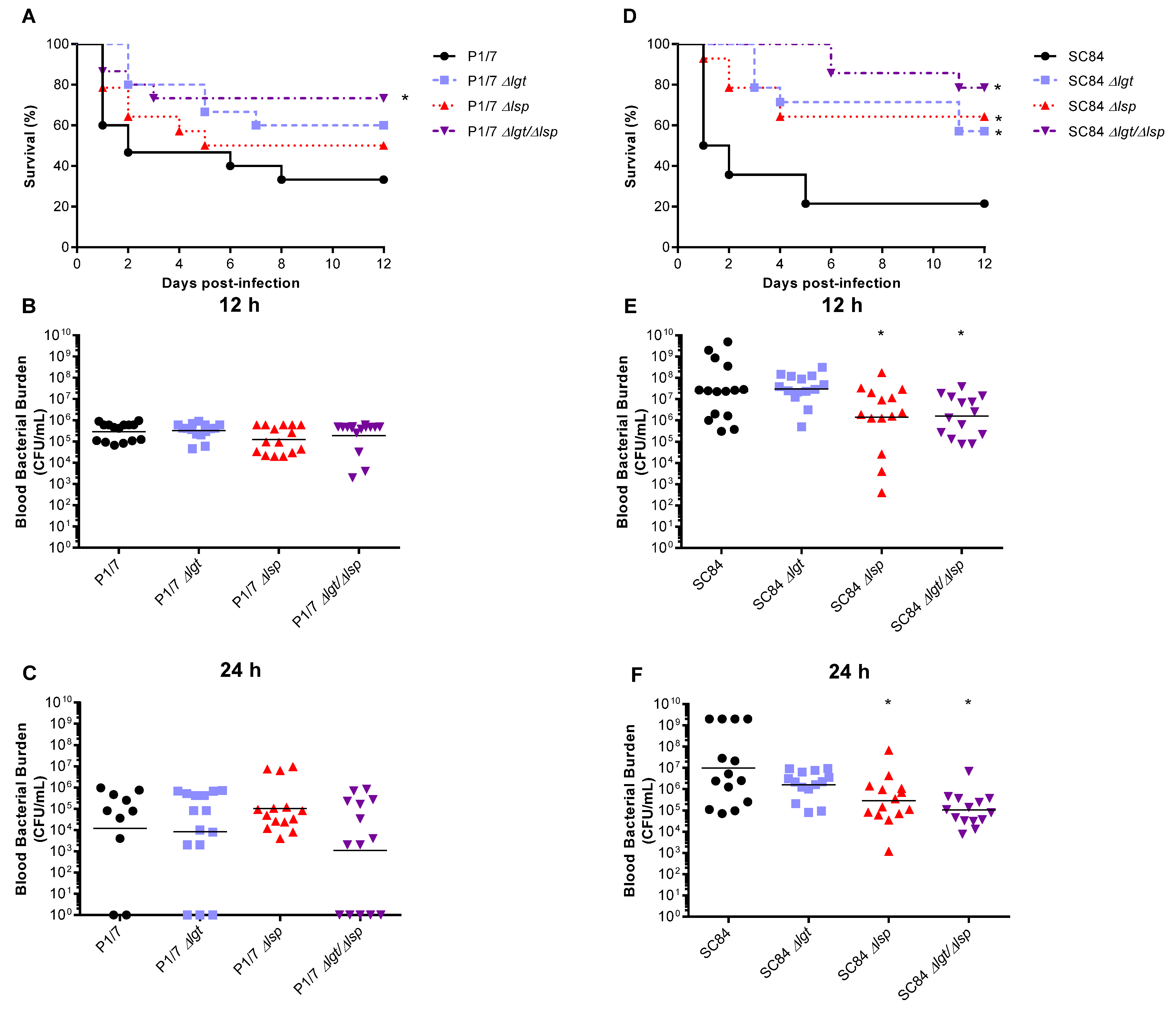
3.5. Absence of the Lgt and/or Lsp Enzymes Affects S. suis Serotype 2 Virulence in a Strain-Dependent Manner
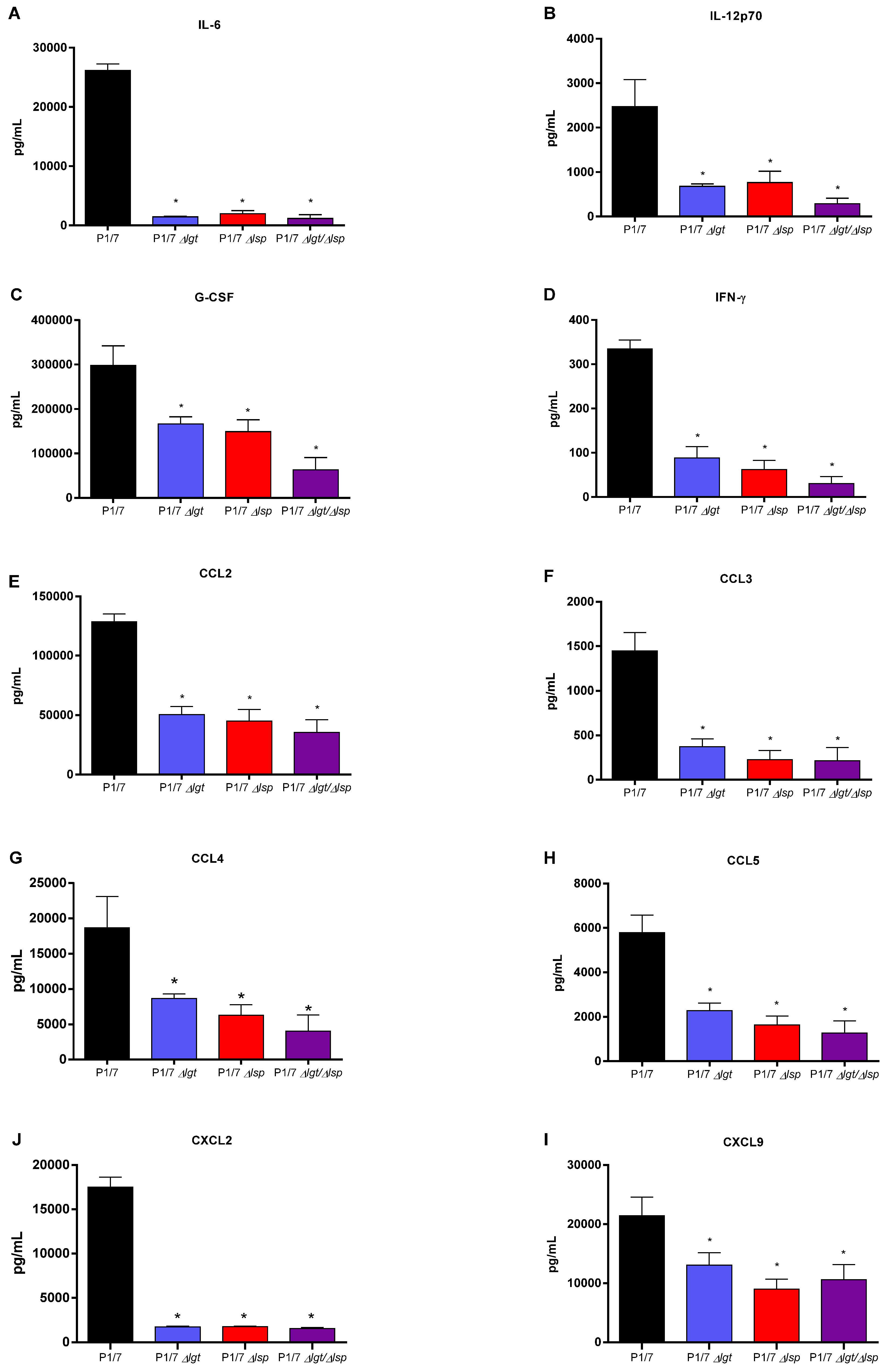
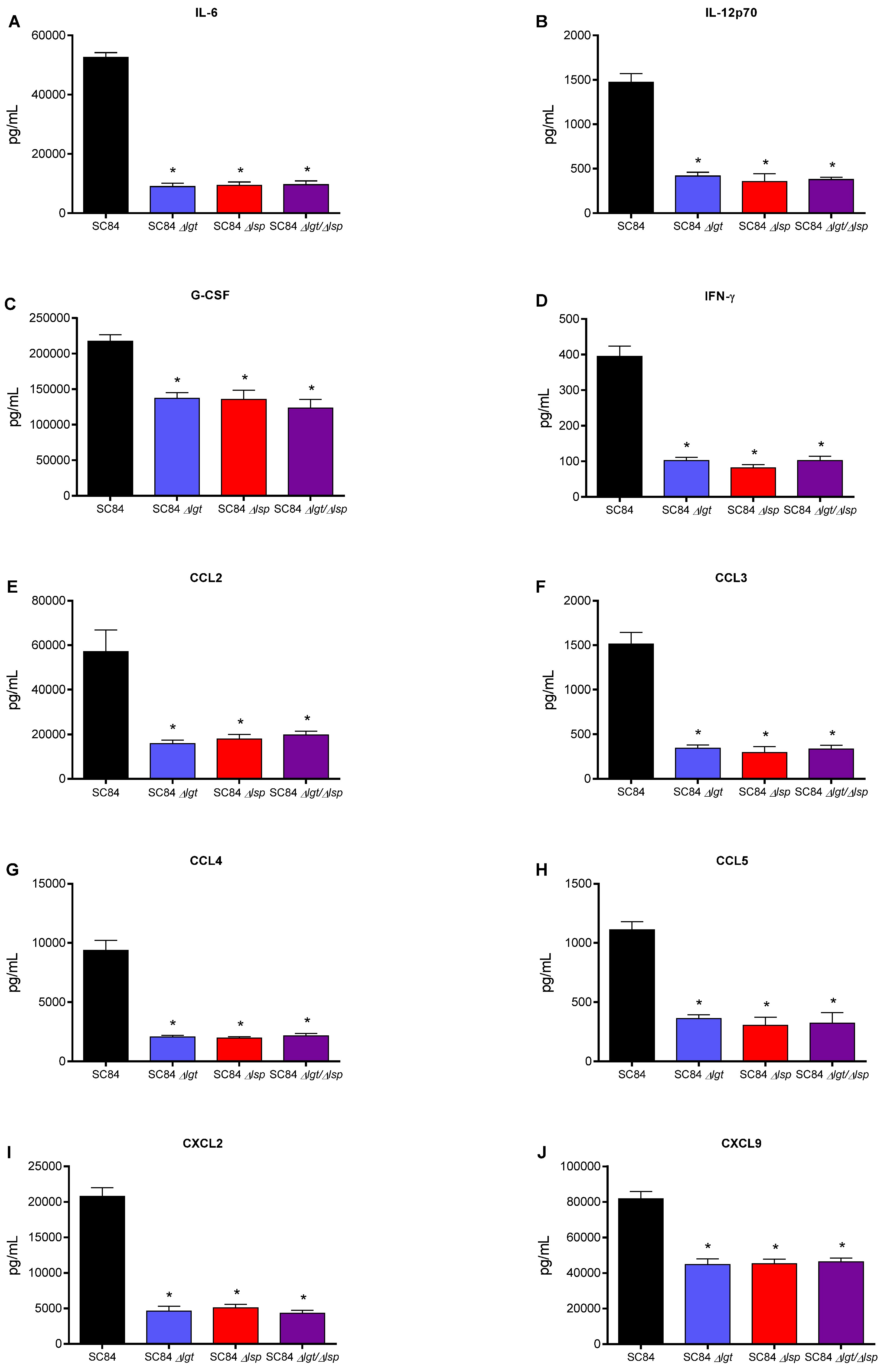
3.6. Absence of the Lsp and/or Lgt Enzymes Reduces the In Vivo Inflammatory Response after Infection of Mice with Both Wild-Type S. suis Serotype 2 Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: A new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010, 5, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microb. Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis Virulence Factors: Are They All Really Critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Meekhanon, N.; Okura, M.; Osaki, M.; Gottschalk, M.; Sekizaki, T.; Takamatsu, D. Streptococcus suis serotype 2 capsule in vivo. Emerg. Infect. Dis. 2016, 22, 1793–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef]
- Tanabe, S.; Bonifait, L.; Fittipaldi, N.; Grignon, L.; Gottschalk, M.; Grenier, D. Pleiotropic effects of polysaccharide capsule loss on selected biological properties of Streptococcus suis. Can. J. Vet. Res. 2010, 74, 65–70. [Google Scholar] [PubMed]
- Wichgers Schreur, P.J.; Rebel, J.M.; Smits, M.A.; van Putten, J.P.; Smith, H.E. Differential activation of the Toll-like receptor 2/6 complex by lipoproteins of Streptococcus suis serotypes 2 and 9. Vet. Microbiol. 2010, 143, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Lecours, M.P.; Gottschalk, M.; Houde, M.; Lemire, P.; Fittipaldi, N.; Segura, M. Critical role for Streptococcus suis cell wall modifications and suilysin in resistance to complement-dependent killing by dendritic cells. J. Infect. Dis. 2011, 204, 919–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisch, N.; Auger, J.P.; Thomsen, S.; Roy, D.; Xu, J.; Schwudke, D.; Gottschalk, M. Structural analysis and immunostimulatory potency of lipoteichoic acids isolated from three Streptococcus suis serotype 2 strains. J. Biol. Chem. 2018, 293, 12011–12025. [Google Scholar] [CrossRef] [Green Version]
- Kohler, S.; Voss, F.; Gomez Mejia, A.; Brown, J.S.; Hammerschmidt, S. Pneumococcal lipoproteins involved in bacterial fitness, virulence, and immune evasion. FEBS Lett. 2016, 590, 3820–3839. [Google Scholar] [CrossRef]
- Sander, P.; Rezwan, M.; Walker, B.; Rampini, S.K.; Kroppenstedt, R.M.; Ehlers, S.; Keller, C.; Keeble, J.R.; Hagemeier, M.; Colston, M.J.; et al. Lipoprotein processing is required for virulence of Mycobacterium tuberculosis. Mol. Microbiol. 2004, 52, 1543–1552. [Google Scholar] [CrossRef]
- Henneke, P.; Dramsi, S.; Mancuso, G.; Chraibi, K.; Pellegrini, E.; Theilacker, C.; Hubner, J.; Santos-Sierra, S.; Teti, G.; Golenbock, D.T.; et al. Lipoproteins are critical TLR2 activating toxins in group B streptococcal sepsis. J. Immunol. 2008, 180, 6149–6158. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Tawaratsumida, K.; Kariya, H.; Kiyohara, A.; Suda, Y.; Krikae, F.; Kirikae, T.; Götz, F. Not lipoteichoic acid but lipoproteins appear to be the dominant immunobiologically active compounds in Staphylococcus aureus. J. Immunol. 2006, 177, 3162–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, M.I.; Palmer, T.; Harrington, D.J.; Sutcliffe, I.C. Lipoprotein biogenesis in Gram-positive bacteria: Knowing when to hold ‘em, knowing when to fold ‘em. Trends Microbiol. 2009, 17, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs-Simon, A.; Titball, R.W.; Michell, S.L. Lipoproteins of bacterial pathogens. Infect. Immun. 2011, 79, 548–561. [Google Scholar] [CrossRef] [Green Version]
- Pribyl, T.; Moche, M.; Dreisbach, A.; Bijlsma, J.J.; Saleh, M.; Abdullah, M.R.; Hecker, M.; van Dijl, J.M.; Becher, D.; Hammerschmidt, S. Influence of impaired lipoprotein biogenesis on surface and exoproteome of Streptococcus pneumoniae. J. Proteome Res. 2014, 13, 650–667. [Google Scholar] [CrossRef]
- Bray, B.A.; Sutcliffe, I.C.; Harrington, D.J. Impact of lgt mutation on lipoprotein biosynthesis and in vitro phenotypes of Streptococcus agalactiae. Microbiology 2009, 155, 1451–1458. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, G.; Chimalapati, S.; Pollard, T.; Lapp, T.; Cohen, J.; Camberlein, E.; Stafford, S.; Periselneris, J.; Aldridge, C.; Vollmer, W.; et al. TLR-mediated inflammatory responses to Streptococcus pneumoniae are highly dependent on surface expression of bacterial lipoproteins. J. Immunol. 2014, 193, 3736–3745. [Google Scholar] [CrossRef] [Green Version]
- Petit, C.M.; Brown, J.R.; Ingraham, K.; Bryant, A.P.; Holmes, D.J. Lipid modification of prelipoproteins is dispensable for growth in vitro but essential for virulence in Streptococcus pneumoniae. FEMS Microbiol. Lett. 2001, 200, 229–233. [Google Scholar] [CrossRef]
- Chimalapati, S.; Cohen, J.M.; Camberlein, E.; MacDonald, N.; Durmort, C.; Vernet, T.; Hermans, P.W.; Mitchell, T.; Brown, J.S. Effects of deletion of the Streptococcus pneumoniae lipoprotein diacylglyceryl transferase gene lgt on ABC transporter function and on growth in vivo. PLoS ONE 2012, 7, e41393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, A.; Robinson, C.; Sutcliffe, I.C.; Slater, J.; Maskell, D.J.; Davis-Poynter, N.; Smith, K.; Waller, A.; Harrington, D.J. Mutation of the maturase lipoprotein attenuates the virulence of Streptococcus equi to a greater extent than does loss of general lipoprotein lipidation. Infect. Immun. 2006, 74, 6907–6919. [Google Scholar] [CrossRef] [Green Version]
- De Greeff, A.; Hamilton, A.; Sutcliffe, I.C.; Buys, H.; van Alphen, L.; Smith, H.E. Lipoprotein signal peptidase of Streptococcus suis serotype 2. Microbiology 2003, 149, 1399–1407. [Google Scholar] [CrossRef]
- Wichgers Schreur, P.J.; Rebel, J.M.; Smits, M.A.; van Putten, J.P.; Smith, H.E. Lgt processing is an essential step in Streptococcus suis lipoprotein mediated innate immune activation. PLoS ONE 2011, 6, e22299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavagna, A.; Auger, J.P.; Dumesnil, A.; Roy, D.; Girardin, S.E.; Gisch, N.; Segura, M.; Gottschalk, M. Interleukin-1 signaling induced by Streptococcus suis serotype 2 is strain-dependent and contributes to bacterial clearance and inflammation during systemic disease in a mouse model of infection. Vet. Res. 2019, 50, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecours, M.P.; Segura, M.; Fittipaldi, N.; Rivest, S.; Gottschalk, M. Immune receptors involved in Streptococcus suis recognition by dendritic cells. PLoS ONE 2012, 7, e44746. [Google Scholar] [CrossRef]
- Auger, J.-P.; Santinón, A.; Roy, D.; Mossman, K.; Xu, J.; Segura, M.; Gottschalk, M. Type I interferon induced by Streptococcus suis serotype 2 is strain-dependent and may be beneficial for host survival. Front. Immunol. 2017, 8, 1039. [Google Scholar] [CrossRef] [Green Version]
- Auger, J.-P.; Fittipaldi, N.; Benoit-Biancamano, M.-O.; Segura, M.; Gottschalk, M. Virulence studies of different sequence types and geographical origins of Streptococcus suis serotype 2 in a mouse model of infection. Pathogens 2016, 5, 48. [Google Scholar] [CrossRef]
- Dominguez-Punaro, M.; Segura, M.; Plante, M.M.; Lacouture, S.; Rivest, S.; Gottschalk, M. Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J. Immunol. 2007, 179, 1842–1854. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Punaro, M.; Segura, M.; Radzioch, D.; Rivest, S.; Gottschalk, M. Comparison of the susceptibilities of C57BL/6 and A/J mouse strains to Streptococcus suis serotype 2 infection. Infect. Immun. 2008, 76, 3901–3910. [Google Scholar] [CrossRef] [Green Version]
- Slater, J.D.; Allen, A.G.; May, J.P.; Bolitho, S.; Lindsay, H.; Maskell, D.J. Mutagenesis of Streptococcus equi and Streptococcus suis by transposon Tn917. Vet. Microbiol. 2003, 93, 197–206. [Google Scholar] [CrossRef]
- Ye, C.; Zheng, H.; Zhang, J.; Jing, H.; Wang, L.; Xiong, Y.; Wang, W.; Zhou, Z.; Sun, Q.; Luo, X.; et al. Clinical, experimental, and genomic differences between intermediately pathogenic, highly pathogenic, and epidemic Streptococcus suis. J. Infect. Dis. 2009, 199, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Casadaban, M.J.; Cohen, S.N. Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J. Mol. Biol. 1980, 138, 179–207. [Google Scholar] [CrossRef]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 2001, 46, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Okura, M.; Osaki, M.; Fittipaldi, N.; Gottschalk, M.; Sekizaki, T.; Takamatsu, D. The minor pilin subunit Sgp2 is necessary for assembly of the pilus encoded by the srtG cluster of Streptococcus suis. J. Bacteriol. 2011, 193, 822–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrens, A.N.; Jones, M.D.; Lechler, R.I. Splicing by overlap extension by PCR using asymmetric amplification: An improved technique for the generation of hybrid proteins of immunological interest. Gene 1997, 186, 29–35. [Google Scholar] [CrossRef]
- Auger, J.-P.; Chuzeville, S.; Roy, D.; Mathieu-Denoncourt, A.; Xu, J.; Grenier, D.; Gottschalk, M. The bias of experimental design, including strain background, in the determination of critical Streptococcus suis serotype 2 virulence factors. PLoS ONE 2017, 12, e0181920. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Construction and characterization of Streptococcus suis-Escherichia coli shuttle cloning vectors. Plasmid 2001, 45, 101–113. [Google Scholar] [CrossRef]
- Auger, J.P.; Dolbec, D.; Roy, D.; Segura, M.; Gottschalk, M. Role of the Streptococcus suis serotype 2 capsular polysaccharide in the interactions with dendritic cells is strain-dependent but remains critical for virulence. PLoS ONE 2018, 13, e0200453. [Google Scholar] [CrossRef]
- Segura, M.; Stankova, J.; Gottschalk, M. Heat-killed Streptococcus suis capsular type 2 strains stimulate tumor necrosis factor alpha and interleukin-6 production by murine macrophages. Infect. Immun. 1999, 67, 4646–4654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gagnon, C.A.; Savard, C.; Music, N.; Srednik, M.; Segura, M.; Lachance, C.; Bellehumeur, C.; Gottschalk, M. Capsular sialic acid of Streptococcus suis serotype 2 binds to swine influenza virus and enhances bacterial interactions with virus-infected tracheal epithelial cells. Infect. Immun. 2013, 81, 4498–4508. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.; Su, Z.; Piccirillo, C.; Stevenson, M.M. Impairment of dendritic cell function by excretory-secretory products: A potential mechanism for nematode-induced immunosuppression. Eur. J. Immunol. 2007, 37, 1887–1904. [Google Scholar] [CrossRef] [PubMed]
- Lavagna, A.; Auger, J.P.; Girardin, S.E.; Gisch, N.; Segura, M.; Gottschalk, M. Recognition of Lipoproteins by Toll-like Receptor 2 and DNA by the AIM2 Inflammasome Is Responsible for Production of Interleukin-1beta by Virulent Suilysin-negative Streptococcus suis Serotype 2. Pathogens 2020, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachance, C.; Gottschalk, M.; Gerber, P.P.; Lemire, P.; Xu, J.; Segura, M. Exacerbated type II interferon response drives hypervirulence and toxic shock by an emergent epidemic strain of Streptococcus suis. Infect. Immun. 2013, 81, 1928–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, M.; Higgins, R.; Boudreau, M. Use of polyvalent coagglutination reagents for serotyping of Streptococcus suis. J. Clin. Microbiol. 1993, 31, 2192–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutcliffe, I.C.; Russell, R.R. Lipoproteins of gram-positive bacteria. J. Bacteriol 1995, 177, 1123–1128. [Google Scholar] [CrossRef] [Green Version]
- Auger, J.P.; Payen, S.; Roy, D.; Dumesnil, A.; Segura, M.; Gottschalk, M. Interactions of Streptococcus suis serotype 9 with host cells and role of the capsular polysaccharide: Comparison with serotypes 2 and 14. PLoS ONE 2019, 14, e0223864. [Google Scholar] [CrossRef]
- Segura, M.; Calzas, C.; Grenier, D.; Gottschalk, M. Initial steps of the pathogenesis of the infection caused by Streptococcus suis: Fighting against nonspecific defenses. FEBS Lett. 2016, 590, 3772–3799. [Google Scholar] [CrossRef] [Green Version]
- Baums, C.G.; Valentin-Weigand, P. Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim. Health Res. Rev. 2009, 10, 65–83. [Google Scholar] [CrossRef]
- Stoll, H.; Dengjel, J.; Nerz, C.; Götz, F. Staphylococcus aureus deficient in lipidation of prelipoproteins is attenuated in growth and immune activation. Infect. Immun. 2005, 73, 2411–2423. [Google Scholar] [CrossRef] [Green Version]
- Tenenbaum, T.; Spellerberg, B.; Adam, R.; Vogel, M.; Kim, K.S.; Schroten, H. Streptococcus agalactiae invasion of human brain microvascular endothelial cells is promoted by the laminin-binding protein Lmb. Microbes Infect. 2007, 9, 714–720. [Google Scholar] [CrossRef]
- Terao, Y.; Kawabata, S.; Kunitomo, E.; Nakagawa, I.; Hamada, S. Novel laminin-binding protein of Streptococcus pyogenes, Lbp, is involved in adhesion to epithelial cells. Infect. Immun. 2002, 70, 993–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.M.; Shao, Z.Q.; Wang, J.; Wang, L.; Li, X.; Wang, C.; Tang, J.; Pan, X. Prevalent distribution and conservation of Streptococcus suis Lmb protein and its protective capacity against the Chinese highly virulent strain infection. Microbiol. Res. 2014, 169, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Esgleas, M.; Lacouture, S.; Gottschalk, M. Streptococcus suis serotype 2 binding to extracellular matrix proteins. FEMS Microbiol. Lett. 2005, 244, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Pei, X.; Liu, M.; Zhou, H.; Fan, H. Screening for phagocytosis resistance-related genes via a transposon mutant library of Streptococcus suis serotype 2. Virulence 2020, 11, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Bartual, S.G.; Abdullah, M.R.; Jensch, I.; Asmat, T.M.; Petruschka, L.; Pribyl, T.; Gellert, M.; Lillig, C.H.; Antelmann, H.; et al. Molecular architecture of Streptococcus pneumoniae surface thioredoxin-fold lipoproteins crucial for extracellular oxidative stress resistance and maintenance of virulence. EMBO Mol. Med. 2013, 5, 1852–1870. [Google Scholar] [CrossRef]
- Chuzeville, S.; Auger, J.P.; Dumesnil, A.; Roy, D.; Lacouture, S.; Fittipaldi, N.; Grenier, D.; Gottschalk, M. Serotype-specific role of antigen I/II in the initial steps of the pathogenesis of the infection caused by Streptococcus suis. Vet. Res. 2017, 48, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenenbaum, T.; Asmat, T.M.; Seitz, M.; Schroten, H.; Schwerk, C. Biological activities of suilysin: Role in Streptococcus suis pathogenesis. Future Microbiol. 2016, 11, 941–954. [Google Scholar] [CrossRef]
- Réglier-Poupet, H.; Frehel, C.; Dubail, I.; Beretti, J.L.; Berche, P.; Charbit, A.; Raynaud, C. Maturation of lipoproteins by type II signal peptidase is required for phagosomal escape of Listeria monocytogenes. J. Biol. Chem. 2003, 278, 49469–49477. [Google Scholar] [CrossRef] [Green Version]
- Hazenbos, W.L.; Skippington, E.; Tan, M.W. Staphylococcus aureus type I signal peptidase: Essential or not essential, that’s the question. Microb. Cell 2017, 4, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benmimoun, B.; Papastefanaki, F.; Perichon, B.; Segklia, K.; Roby, N.; Miriagou, V.; Schmitt, C.; Dramsi, S.; Matsas, R.; Speder, P. An original infection model identifies host lipoprotein import as a route for blood-brain barrier crossing. Nat. Commun. 2020, 11, 6106. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, M.R.; Corsaut, L.; Dolbec, D.; Gottschalk, M.; Segura, M. Experimental evaluation of protection and immunogenicity of Streptococcus suis bacterin-based vaccines formulated with different commercial adjuvants in weaned piglets. Vet. Res. 2021, 52, 133. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or plasmid | Characteristics | Reference |
|---|---|---|
| Streptococcus suis | ||
| P1/7 | Virulent serotype 2 ST1 strain isolated from a case of pig meningitis in the United Kingdom | [32] |
| P1/7Δlgt | Isogenic mutant derived from P1/7; in frame deletion of the lgt gene | This study |
| P1/7Δlsp | Isogenic mutant derived from P1/7; in frame deletion of the lsp gene | This study |
| P1/7Δlgt/Δlsp | Isogenic mutant derived from P1/7; in frame deletion of the lgt and lsp genes | This study |
| P1/7 comp Δlgt | Mutant Δlgt complemented with the pMX1-lgt complementation vector | This study |
| P1/7 comp Δlsp | Mutant Δlsp complemented with the pMX1-lsp complementation vector | This study |
| SC84 | High virulent serotype 2 ST7 strain isolated from a human streptococcal toxic shock-like syndrome case in China | [33] |
| SC84Δlgt | Isogenic mutant derived from SC84; in frame deletion of the lgt gene | This study |
| SC84Δlsp | Isogenic mutant derived from SC84; in frame deletion of the lsp gene | This study |
| SC84Δlgt/Δlsp | Isogenic mutant derived from SC84; in frame deletion of the lgt and lsp genes | This study |
| SC84 comp Δlgt | Mutant Δlgt complemented with the pMX1-lgt complementation vector | This study |
| SC84 comp Δlsp | Mutant Δlsp complemented with the pMX1-lsp complementation vector | This study |
| Escherichia coli | ||
| TOP10 | F− mrcA Δ(mrr-hsdRMS-mcrBC) φ80 lacZΔM15 ΔlacX74 recA1 araD139 Δ(araleu) 7697 galU galK rpsL (StrR) endA1 nupG | Invitrogen |
| MC1061 | Host for pMX1 derivatives | [34] |
| Plasmids | ||
| pCR2.1 | Apr, Kmr, pUC ori, lacZΔM15 | Invitrogen |
| pSET4s | Spcr, pUC ori, thermosensitive pG+host3 ori, lacZΔM15 | [35] |
| pMX1 | Replication functions of pSSU1, MCS pUC19 lacZ SpR, malX promoter of S. suis, derivative of pSET2 | [35,36] |
| p4Δlg p4Δlsp | pSET-4s carrying the construct for lgt allelic replacement pSET-4s carrying the construct for lsp allelic replacement | This study This study |
| pMX1-lgt | pMX1 carrying the intact lgt gene | This study |
| pMX1-lsp | pMX1 carrying the intact lsp gene | This study |
| Name | Sequence (5′–3′) | Construct |
|---|---|---|
| lgt-ID1 | GGAACGCTATGGAACAGGTC | p4Δlgt |
| lgt-ID2 | CACTCCATGAAAAGGCGACG | p4Δlgt |
| lgt-ID3 | CGTAGACGGCCAAAATTCC | p4Δlgt |
| lgt-ID4 | CGCTTATCTGCTGGATTCTCC | p4Δlgt |
| lgt-ID5 | GCCAATCGTCTGCATCAAGG | p4Δlgt |
| lgt-ID6 | GGGTTGATAGAATGGGATTGCATACCAACG | p4Δlgt |
| lgt-ID7 | CGTTGGTATGCAATCCCATTCTATCAACCC | p4Δlgt |
| lgt-ID8 | GACCGACTTGCTGGTCAAAC | p4Δlgt |
| lsp-ID1 | TGAGAAAACTGTTGTGGGTA | p4Δlsp |
| lsp-ID2 | AGAGCACCAGCAATCATCAA | p4Δlsp |
| lsp-ID3 | TTGATGATTGCTGGTGCTCT | p4Δlsp |
| lsp-ID4 | TAGACAGCGAACAGAGATAC | p4Δlsp |
| lsp-ID5 | GCGCCTGCAGGATGATTTGGCGAACAGAAA | p4Δlsp |
| lsp-ID6 | ACCTACACCAACTGTTAATACTACCATCAA | p4Δlsp |
| lsp-ID7 | TTGATGGTAGTATTAACAGTTGGTGTAGGT | p4Δlsp |
| lsp-ID8 | CGCGCTGCAGTTTTAGTGTTTTCCTTGGGC | p4Δlsp |
| pMX1-lgt-F | CCGCCATGGACAGATGGGGTTTGATGCAAC | pMX1-lgt |
| pMX1-lgt-R | CGCGAATTCGGACAAGGCAATAATCAAGAC | pMX1-lgt |
| pMX1-lsp-F | GTGCCATGGACTTTATTGAAACCATGCAGG | pMX1-lsp |
| pMX1-lsp-R | ATCGAATTCAATACCACCAACCTCAACTCT | pMX1-lsp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Payen, S.; Roy, D.; Boa, A.; Okura, M.; Auger, J.-P.; Segura, M.; Gottschalk, M. Role of Maturation of Lipoproteins in the Pathogenesis of the Infection Caused by Streptococcus suis Serotype 2. Microorganisms 2021, 9, 2386. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112386
Payen S, Roy D, Boa A, Okura M, Auger J-P, Segura M, Gottschalk M. Role of Maturation of Lipoproteins in the Pathogenesis of the Infection Caused by Streptococcus suis Serotype 2. Microorganisms. 2021; 9(11):2386. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112386
Chicago/Turabian StylePayen, Servane, David Roy, Anaïs Boa, Masatoshi Okura, Jean-Philippe Auger, Mariela Segura, and Marcelo Gottschalk. 2021. "Role of Maturation of Lipoproteins in the Pathogenesis of the Infection Caused by Streptococcus suis Serotype 2" Microorganisms 9, no. 11: 2386. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112386