
Psychrophilic Bacterial Phosphate-Biofertilizers: A Novel Extremophile for Sustainable Crop Production under Cold Environment


Abstract
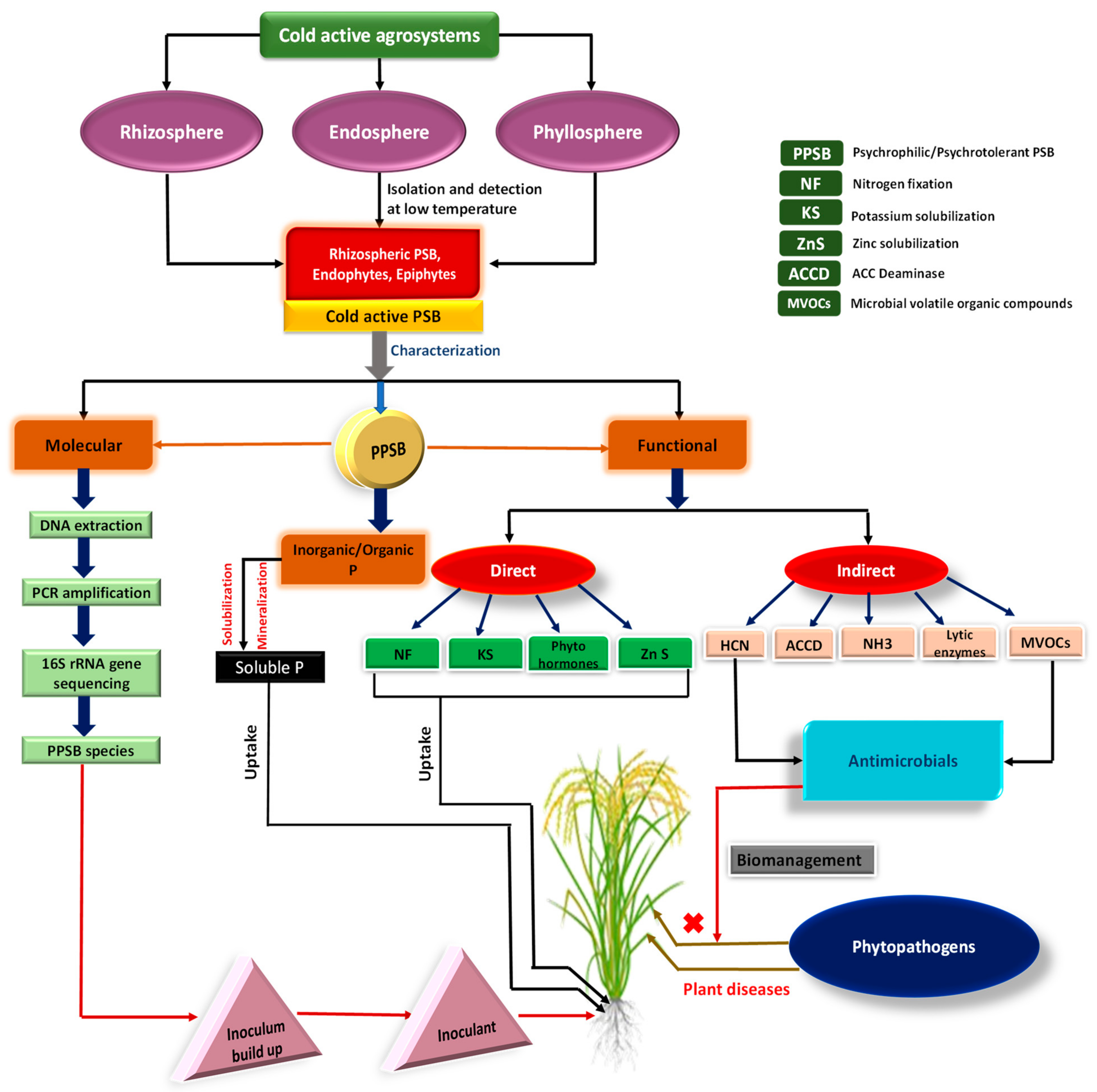
:1. Introduction
2. Psychrophiles and Psychrotrophs: An Overview
2.1. Definition, Ecological Habitats, and Agronomic Importance
2.2. How Do Cold-Active Bacteria Survive under Cold Stress?
3. Low Temperature Effects on Plants: Physiological Changes and Stress Alleviation
4. Importance of P and Rationale for Using Cold-Active Bacterial Phosphate Biofertilizers in Low-Temperature Environments
5. Mechanisms of P-Solubilization in Cold-Adapted Bacteria: An Overview
6. Mechanisms Used by Cold-Adapted PSB to Facilitate Plant Growth
6.1. Direct Mechanisms
6.2. Indirect Mechanisms
7. Performance of Cold-Active P-Biofertilizers under the Low-Temperature Environment
8. Molecular Engineering of Phosphate-Biofertilizers
8.1. Development of mps Positive Bacterial Strains
8.2. Development of Enzyme Engineered Bacterial Strains
8.2.1. Development of Bacterial Strains with Phosphatase Activity
8.2.2. Development of Bacterial Strains with Phytase Activity
9. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, F.; Lu, K.; Gu, Y.; Zhang, L.; Li, W.; Li, Z. Effects of low-temperature stress and brassinolide application on the photosynthesis and leaf structure of tung tree seedlings. Front. Plant Sci. 2020, 10, 1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia-Vázquez, I.; Sánchez-Cruz, R.; Arroyo-Domínguez, M.; Lira-Ruan, V.; Sánchez-Reyes, A.; del Rayo Sánchez-Carbente, M.; Padilla-Chacón, D.; Batista-García, R.A.; Folch-Mallol, J.L. Isolation and characterization of psychrophilic and psychrotolerant plant-growth promoting microorganisms from a high-altitude volcano crater in Mexico. Microbiol. Res. 2020, 232, 126394. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Singh, P.C.; Singh, S. A Review Report: Low-temerature Stress for Crop Production. Int. J. Pure Appl. Biosci. 2018, 6, 575–598. [Google Scholar] [CrossRef]
- Muhammad, I.; Khan, F.; Khan, A.; Wang, J. Soil fertility in response to urea and farmyard manure incorporation under different tillage systems in Peshawar, Pakistan. Int. J. Agric. Biol. 2018, 20, 1539–1547. [Google Scholar]
- Muhammad, I.; Sainju, U.M.; Zhao, F.; Khan, A.; Ghimire, R.; Fu, X.; Wang, J. Regulation of soil CO2 and N2O emissions by cover crops: A meta-analysis. Soil Tillage Res. 2019, 192, 103–112. [Google Scholar] [CrossRef]
- Thant, S.; Aung, N.N.; Aye, O.M. Phosphate solubilization of Bacillus megaterium isolated from non-saline soils under salt stressed conditions. J. Bacteriol. Mycol. Open Access 2018, 6, 335–341. [Google Scholar]
- Lin, Y.T.; Jia, Z.; Wang, D.; Chiu, C.Y. Effects of temperature on the composition and diversity of bacterial communities in bamboo soils at different elevations. Biogeosciences 2017, 14, 4879–4889. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, A.; Zaidi, A.; Ameen, F.; Ahmed, B.; AlKahtani, M.D.; Khan, M.S. Heavy metal induced stress on wheat: Phytotoxicity and microbiological management. RSC Adv. 2020, 10, 38379–38403. [Google Scholar] [CrossRef]
- Zeng, Q.; Wu, X.; Wen, X. Identification and characterization of the rhizosphere phosphate-solubilizing bacterium Pseudomonas frederiksbergensis JW-SD2, and its plant growth-promoting effects on poplar seedlings. Ann. Microbiol. 2016, 66, 1343–1354. [Google Scholar] [CrossRef]
- Vassileva, M.; Serrano, M.; Bravo, V.; Jurado, E.; Nikolaeva, I.; Martos, V.; Vassilev, N. Multifunctional properties of phosphate solubilizing microorganisms grown on agro-industrial wastes in fermentation and soil conditions. Appl. Microbiol. Biotechnol. 2010, 85, 1287–1299. [Google Scholar] [CrossRef] [PubMed]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef]
- Vega-Celedón, P.; Bravo, G.; Velásquez, A.; Cid, F.P.; Valenzuela, M.; Ramírez, I.; Vasconez, I.N.; Álvarez, I.; Jorquera, M.A.; Seeger, M. Microbial diversity of psychrotolerant bacteria isolated from wild flora of Andes mountains and Patagonia of Chile towards the selection of plant growth-promoting bacterial consortia to alleviate cold stress in plants. Microorganisms 2021, 9, 538. [Google Scholar] [CrossRef]
- Cid, F.P.; Inostroza, N.G.; Graether, S.P.; Bravo, L.A.; Jorquera, M.A. Bacterial community structures and ice recrystallization inhibition activity of bacteria isolated from the phyllosphere of the Antarctic vascular plant Deschampsia antarctica. Polar Biol. 2017, 40, 1319–1331. [Google Scholar] [CrossRef]
- Selvakumar, G.; Joshi, P.; Nazim, S.; Mishra, P.; Bisht, J.; Gupta, H. Phosphate solubilization and growth promotion by Pseudomonas fragi CS11RH1 (MTCC 8984), a psychrotolerant bacterium isolated from a high altitude Himalayan rhizosphere. Biologia 2009, 64, 239–245. [Google Scholar] [CrossRef]
- Negi, Y.K.; Garg, S.K.; Kumar, J. Cold-tolerant fluorescent Pseudomonas isolates from Garhwal Himalayas as potential plant growth promoting and biocontrol agents in pea. Curr. Sci. 2005, 25, 2151–2156. [Google Scholar]
- Rondón, J.J.; Ball, M.M.; Castro, L.T.; Yarzábal, L.A. Eurypsychrophilic Pseudomonas spp. Isolated from Venezuelan tropical glaciers as promoters of wheat growth and biocontrol agents of plant pathogens at low-temperatures. Environ. Sustain. 2019, 2, 265–275. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Kaushik, R.; Saxena, A.K. Cold active hydrolytic enzymes production by psychrotrophic bacilli isolated from three sub-glacial lakes of NW Indian Himalayas. J. Basic Microbiol. 2016, 56, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Balcázar, W.; Rondón, J.; Rengifo, M.; Ball, M.M.; Melfo, A.; Gómez, W.; Yarzábal, L.A. Bioprospecting glacial ice for plant growth promoting bacteria. Microbiol. Res. 2015, 177, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.R.; Seghal, A.; Jumaa, S.; Bheemanahalli, R.; Kakar, N.; Redoña, E.D.; Wijewardana, C.; Alsajri, F.A.; Chastain, D.; Gao, W. Morpho-physiological characterization of diverse rice genotypes for seedling stage high-and low-temperature tolerance. Agronomy 2021, 11, 112. [Google Scholar] [CrossRef]
- Adhikari, P.; Jain, R.; Sharma, A.; Pandey, A. Plant growth promotion at low-temperature by phosphate-solubilizing Pseudomonas spp. isolated from high-altitude Himalayan soil. Microb. Ecol. 2021, 29, 1–11. [Google Scholar] [CrossRef]
- Patni, B.; Panwar, A.; Negi, P.; Joshi, G.K. Plant growth promoting traits of psychrotolerant bacteria: A boon for agriculture in hilly terrains. Plant Sci. Today 2018, 5, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Meena, R.K.; Singh, R.K.; Singh, N.P.; Meena, S.K.; Meena, V.S. Isolation of low-temperature surviving plant growth–promoting rhizobacteria (PGPR) from pea (Pisum sativum L.) and documentation of their plant growth promoting traits. Biocat. Agric. Biotechnol. 2015, 4, 806–811. [Google Scholar] [CrossRef]
- Zubair, M.; Hanif, A.; Farzand, A.; Sheikh, T.M.M.; Khan, A.R.; Suleman, M.; Ayaz, M.; Gao, X. Genetic screening and expression analysis of psychrophilic Bacillus spp. reveal their potential to alleviate cold stress and modulate phytohormones in wheat. Microorganisms 2019, 7, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przemieniecki, S.W.; Kurowski, T.P.; Korzekwa, K.; Karwowska, A. The effect of psychrotrophic bacteria isolated from the root zone of winter wheat on selected biotic and abiotic factors. J. Plant Prot. Res. 2014, 54, 407–413. [Google Scholar] [CrossRef]
- Morel, M.A.; Tapia-Vázquez, I.; Castro-Sowinski, S.; Batista-García, R.A. Fighting plant pathogens with cold-active microorganisms: Biopesticide development and agriculture intensification in cold climates. Appl. Microbiol. Biotechnol. 2020, 17, 1–4. [Google Scholar]
- Pal, K.K.; Tilak, K.V.B.R.; Saxena, A.K.; Dey, R.; Singh, C.S. Antifungal characteristics of a fluorescent Pseudomonas strain involved in the biological control of Rhizoctonia solani. Microbiol. Res. 2001, 155, 233–242. [Google Scholar] [CrossRef]
- Selvakumar, G.; Joshi, P.; Suyal, P.; Mishra, P.K.; Joshi, G.K.; Venugopalan, R.; Bisht, J.K.; Bhatt, J.C. Gupta, H.S. Rock phosphate solubilization by psychrotolerant Pseudomonas spp. and their efect on lentil growth and nutrient uptake under polyhouse conditions. Ann. Microbiol. 2013, 63, 1353–1362. [Google Scholar] [CrossRef]
- Bhandari, G.; Nautiyal, N.; Sharma, M. Assessment of Genes and Enzymes of Microorganisms of High Altitudes and Their Application in Agriculture. In Microbiological Advancements for Higher Altitude Agro-Ecosystems & Sustainability; Springer: Singapore, 2020; pp. 307–326. [Google Scholar]
- Margesin, R.; Miteva, V. Diversity and ecology of psychrophilic microorganisms. Res. Microbiol. 2011, 162, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 146–167. [Google Scholar] [CrossRef] [PubMed]
- Rondón, J.; Gómez, W.; Ball, M.M.; Melfo, A.; Rengifo, M.; Balcázar, W.; Dávila-Vera, D.; Balza-Quintero, A.; Mendoza-Briceño, R.V.; Yarzábal, L.A. Diversity of culturable bacteria recovered from Pico Bolívar’s glacial and subglacial environments, at 4950 m, in Venezuelan tropical Andes. Can. J. Microbiol. 2016, 62, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ball, M.M.; Gómez, W.; Magallanes, X.; Moreno, R.; Melfo, A.; Yarzábal, L.A. Bacteria recovered from a high-altitude, tropical glacier in Venezuelan Andes. World J. Microbiol. Biotechnol. 2014, 30, 931–941. [Google Scholar] [CrossRef]
- Presta, L.; Inzucchi, I.; Bosi, E.; Fondi, M.; Perrin, E.; Maida, I.; Miceli, E.; Tutino, M.L.; Lo Giudice, A.; de Pascale, D. Draft genome sequences of the antimicrobial producers Pseudomonas sp. TAA207 and Pseudomonas sp. TAD18 isolated from Antarctic sediments. Genome Announc. 2016, 4, e00728. [Google Scholar] [CrossRef] [Green Version]
- See-Too, W.S.; Lim, Y.L.; Ee, R.; Convey, P.; Pearce, D.A.; Yin, W.F.; Chan, K.G. Complete genome of Pseudomonas sp. strain L10.10, a psychrotolerant biofertilizer that could promote plant growth. J. Biotechnol. 2016, 222, 84–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Saxena, A.K. Prospecting cold deserts of north western Himalayas for microbial diversity and plant growth promoting attributes. J. Biosci. Bioeng. 2015, 119, 683–693. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Tyagi, S.P.; Kaushik, R.; Saxena, A.K. Culturable diversity and functional annotation of psychrotrophic bacteria from cold desert of Leh Ladakh (India). World J. Microbiol. Biotechnol. 2015, 31, 95–108. [Google Scholar] [CrossRef]
- Bisht, S.C.; Mishra, P.K.; Joshi, G.K. Genetic and functional diversity among root-associated psychrotrophic Pseudomonad’s isolated from the Himalayan plants. Arch. Microbiol. 2013, 195, 605–615. [Google Scholar] [CrossRef]
- Meyer, A.F.; Lipson, D.A.; Schadt, C.W.; Martin, A.P.; Schmidt, S.Z. Molecular and metabolic characterization of cold-tolerant alpine soil Pseudomonas sensustricto. Appl. Environ. Microbiol. 2004, 70, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancinelli, R.L. Population-dynamics of alpine tundra soil bacteria, Niwot Ridge, Colorado Front Range, USA. Arc. Alp. Res. 1984, 16, 185–192. [Google Scholar] [CrossRef]
- Dolkar, D.; Dolkar, P.; Stobdan, T.; Chaurasia, O.P.; Katiyar, A.K. Tomato growth promotion induced by stress tolerant phosphate solubilising Pseudomonas simiae in Arid trans-Himalaya. Defence Life Sci. J. 2018, 3, 105–110. [Google Scholar] [CrossRef]
- Mishra, P.K.; Bisht, S.C.; Ruwari, P.; Selvakumar, G.; Joshi, G.K.; Bisht, J.K.; Bhatt, J.C.; Gupta, H.S. Alleviation of cold stress in inoculated wheat (Triticum aestivum L.) seedlings with psychrotolerant Pseudomonads from NW Himalayas. Arch. Microbiol. 2011, 193, 497–513. [Google Scholar] [CrossRef]
- Araya, M.A.; Valenzuela, T.; Inostroza, N.G.; Maruyama, F.; Jorquera, M.A.; Acuña, J.J. Isolation and characterization of cold-tolerant hyper-ACC-degrading bacteria from the rhizosphere, endosphere, and phyllosphere of antarctic vascular plants. Microorganisms 2021, 8, 1788. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Kumar, S.; Saxena, A.K.; Suman, A. Molecular diversity and multifarious plant growth promoting attributes of Bacilli associated with wheat rhizosphere from six agro-ecological zones of India. J. Basic Microbiol. 2015, 56, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Vyas, P.; Joshi, R.; Sharma, K.C.; Rahi, P.; Gulati, A.; Gulati, A. Cold-adapted and rhizosphere-competent strain of Rahnella sp. with broad-spectrum plant growth-promotion potential. J. Microbiol. Biotechnol. 2010, 20, 1724–1734. [Google Scholar] [PubMed]
- Kadioglu, G.B.; Koseoglu, M.S.; Ozdal, M.U.; Sezen, A.L.; Ozdal, O.G.; Algur, O.F. Isolation of cold tolerant and ACC deaminase producing plant growth promoting rhizobacteria from high altitudes. Rom. Biotechnol. Lett. 2018, 23, 13479–13486. [Google Scholar]
- Herbert, R. Psychrophilic organisms. In Microbes in Extreme Environments; Herbert, R., Codd, G., Eds.; Academic Press: London, UK, 1986; pp. 1–23. [Google Scholar]
- Subramanian, P.; Kim, K.; Krishnamoorthy, R.; Mageswari, A.; Selvakumar, G.; Sa, T. Cold stress tolerance in psychrotolerant soil bacteria and their conferred chilling resistance in tomato (Solanum lycopersicum Mill.) under low-temperatures. PLoS ONE 2016, 11, e0161592. [Google Scholar] [CrossRef]
- Furhan, J. Adaptation, production, and biotechnological potential of cold-adapted proteases from psychrophiles and psychrotrophs: Recent overview. J. Gen. Eng. Biotechnol. 2020, 18, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Feller, G. Psychrophilic enzymes: From folding to function and biotechnology. Scientifica 2013, 2013, 512840. [Google Scholar] [CrossRef]
- Sun, X.; Griffith, M.; Pasternak, J.J.; Glick, B.R. Low-temerature growth, freezing survival, and production of antifreeze protein by the plant growth promoting rhizobacterium Pseudomonas putida GR12-2. Can. J. Microbiol. 1995, 41, 776–784. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K.; Reddy, G.S.; Shivaji, S. Psychrophilic bacteria: Biodiversity, molecular basis of cold adaptation and biotechnological implications. Curr. Opin. Biotechnol. 2014, 3, 100–116. [Google Scholar] [CrossRef]
- Bej, A.K.; Aislabie, J.; Atlas, R.M. Polar Microbiology: The Ecology, Biodiversity and Bioremediation Potential of Microorganisms in Extremely Cold Environments; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Králová, S. Role of fatty acids in cold adaptation of Antarctic psychrophilic Flavobacterium spp. Syst. Appl. Microbiol. 2017, 40, 329–333. [Google Scholar] [CrossRef]
- Ernst, F.G.; Erber, L.; Sammler, J.; Jühling, F.; Betat, H.; Mörl, M. Cold adaptation of tRNA nucleotidyl transferases: A tradeoff in activity, stability and fidelity. RNA Biol. 2018, 15, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Suyal, D.C.; Soni, R.; Yadav, A.N.; Goel, R. Cold Adapted Microorganisms: Survival Mechanisms and Applications. In Microbiomes of Extreme Environments; CRC Press: Boca Raton, FL, USA, 2021; pp. 177–191. [Google Scholar]
- Khan, N.M.; Arai, T.; Tsuda, S.; Kondo, H. Characterization of microbial antifreeze protein with intermediate activity suggests that a bound-water network is essential for hyperactivity. Sci. Rep. 2021, 11, 5971. [Google Scholar] [CrossRef]
- Muñoz, P.A.; Márquez, S.L.; González-Nilo, F.D.; Márquez-Miranda, V.; Blamey, J.M. Structure and application of antifreeze proteins from Antarctic bacteria. Microb. Cell Factor. 2017, 16, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Ali, P.; Shah, A.A.; Hasan, F.; Hertkorn, N.; Gonsior, M.; Sajjad, W.; Chen, F. A glacier bacterium produces high yield of cryoprotective exopolysaccharide. Front. Microbiol. 2020, 10, 3096. [Google Scholar] [CrossRef] [PubMed]
- Deming, J.W.; Young, J.N. The role of exopolysaccharides in microbial adaptation to cold habitats. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Cham, Switzerland, 2017; pp. 259–284. [Google Scholar]
- Casillo, A.; Parrilli, E.; Sannino, F.; Mitchell, D.E.; Gibson, M.I.; Marino, G.; Lanzetta, R.; Parrilli, M.; Cosconati, S.; Novellino, E.; et al. Structure-activity relationship of the exopolysaccharide from a psychrophilic bacterium: A strategy for cryoprotection. Carbohydr. Polym. 2017, 156, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrion, O.; Delgado, L.; Mercade, E. New emulsifying and cryoprotective exopolysaccharide from Antarctic Pseudomonas sp. ID1. Carbohydr. Polym. 2015, 117, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Bremer, E. Protection of Bacillus subtilis against cold stress via compatible-solute acquisition. J. Bacteriol. 2011, 193, 1552–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christner, B.C. Bioprospecting for microbial products that affect ice crystal formation and growth. Appl. Microbiol. Biotechnol. 2010, 85, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Moreno, R.; Rojo, F. Features of Pseudomonads growing at low-temperatures: Another facet of their versatility. Environ. Microbiol. Rep. 2014, 6, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Shivaji, S.; Prakash, J.S.S. How do bacteria sense and respond to low-temperature? Arch. Microbiol. 2010, 192, 85–95. [Google Scholar] [CrossRef]
- Allen, D.; Huston, A.L.; Wells, L.E.; Deming, J.W. Biotechnological use of psychrophiles. In Psychrophiles: From Biodiversity to Biotechnology; Bitton, G., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2001; pp. 1–17. [Google Scholar]
- D’Amico, S.; Collins, T.; Marx, J.C.; Feller, G.; Gerday, C. Psychrophilic microorganisms: Challenges for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Feller, G. Life at low-temeratures: Is disorder the driving force? Extremophiles 2007, 11, 211–216. [Google Scholar] [CrossRef]
- Eskandari, A.; Leow, T.C.; Rahman, M.B.; Oslan, S.N. Antifreeze proteins and their practical utilization in industry, medicine, and agriculture. Biomolecules 2020, 10, 1649. [Google Scholar] [CrossRef] [PubMed]
- Phadtare, S. Recent developments in bacterial cold-shock response. Curr. Issues Mol. Biol. 2004, 6, 125–136. [Google Scholar]
- Gilbert, J.A.; Davies, P.L.; Laybourn−Parry, J. A hyperactive, Ca2+−dependent antifreeze protein in an Antarctic bacterium. FEMS Microbiol. Lett. 2005, 245, 67–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, C.A.M.; Guezennec, J.; Bowman, J.P. Bacterial exopolysaccharides from extreme marine environments with special consideration of the southern ocean, sea ice, and deep-sea hydrothermal vents: A review. Mar. Biotechnol. 2005, 7, 253–271. [Google Scholar] [CrossRef] [Green Version]
- Monsalves, M.T.; Ollivet-Besson, G.P.; Amenabar, M.J.; Blamey, J.M. Isolation of a psychrotolerant and UV-C-resistant bacterium from elephant island, antarctica with a highly thermoactive and thermostable catalase. Microorganisms 2020, 8, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.Z.; Jin, Y.N.; Ding, X.H.; Wang, W.J.; Zhai, S.S.; Bai, L.P.; Guo, Z.F. Gene regulation and signal transduction in the ICE–CBF–COR signaling pathway during cold stress in plants. Biochemistry 2017, 82, 1103–1117. [Google Scholar] [CrossRef]
- Castellano, I.; Di Maro, A.; Ruocco, M.R.; Chambery, A.; Parente, A.; Di Martino, M.T.; Parlato, G.; Masullo, M.; De Vendittis, E. Psychrophilic superoxide dismutase from Pseudoalteromonas haloplanktis: Biochemical characterization and identification of a highly reactive cysteine residue. Biochimie 2006, 88, 1377–1389. [Google Scholar] [CrossRef]
- Chi, Y.X.; Yang, L.; Zhao, C.J.; Muhammad, I.; Zhou, X.B.; De Zhu, H. Effects of soaking seeds in exogenous vitamins on active oxygen metabolism and seedling growth under low-temperature stress. Saudi J. Biol. Sci. 2021, 28, 3254–3261. [Google Scholar]
- Sanghera, G.S. Sugarcane disorders associated with temperature extremes and mitigation strategies. East Afr. Sch. J. Agric. Life Sci. 2020, 3, 101–114. [Google Scholar]
- Hussain, S.; Khaliq, A.; Ali, B.; Hussain, H.A.; Qadir, T.; Hussain, S. Temperature Extremes: Impact on rice growth and development. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; Volume 4, pp. 153–171. [Google Scholar]
- Alsamir, M.; Mahmood, T.; Trethowan, R.; Ahmad, N. An overview of heat stress in tomato (Solanum lycopersicum L.). Saudi J. Biol. Sci. 2021, 3, 1654. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Lindsay, D.L.; Davis, A.R.; Wang, Z.; MacLean, D.E.; Warkentin, T.D.; Bueckert, R.A. Impact of heat stress on pod-based yield components in field pea (Pisum sativum L.). J. Agron. Crop Sci. 2020, 206, 76–89. [Google Scholar] [CrossRef]
- Alsajri, F.A.; Singh, B.; Wijewardana, C.; Irby, J.T.; Gao, W.; Reddy, K.R. Evaluating soybean cultivars for low- and high temperature tolerance during the seedling growth stage. Agronomy 2019, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Singhal, P.; Jan, A.T.; Azam, M.; Haq, Q.M.R. Plant abiotic stress: A prospective strategy of exploiting promoters as alternative to overcome the escalating burden. Front. Life Sci. 2016, 9, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Zaheer, A.; Raza, Z.A.; Mahmood, S. Effect of salinity stress on various growth and physiological attributes of two contrasting maize genotypes. Braz. Arch. Biol. Technol. 2020, 18, 63. [Google Scholar]
- Zhang, H.; Duan, L.; Dai, J.S.; Zhang, C.Q.; Li, J.; Gu, M.H.; Liu, Q.Q.; Zhu, Y. Major QTLs reduce the deleterious effects of high temperature on rice amylose content by increasing splicing efficiency of Wx pre-mRNA. Theor. Appl. Genet. 2014, 127, 273–282. [Google Scholar] [CrossRef]
- Sowiński, P.; Fronk, J.; Jończyk, M.; Grzybowski, M.; Kowalec, P.; Sobkowiak, A. Maize response to low-temeratures at the gene expression level: A critical survey of transcriptomic studies. Front. Plant Sci. 2020, 11, 576941. [Google Scholar] [CrossRef]
- Riva-Roveda, L.; Perilleux, C. Effects of cold temperatures on the early stages of maize (Zea mays L.): A review. Biotechnol. Agron. Soc. Environ. 2015, 19, 42–52. [Google Scholar]
- Alt, D.S.; Lindsey, A.J.; Sulc, R.M.; Lindsey, L.E. Effect of temperature on survival and yield components of field-acclimated soft red winter wheat. Crop Sci. 2020, 60, 475–484. [Google Scholar] [CrossRef]
- Zheng, D.; Yang, X.; Mínguez, M.I.; Mu, C.; He, Q.; Wu, X. Effect of freezing temperature and duration on winter survival and grain yield of winter wheat. Agric. For. Meteorol. 2018, 260, 1–8. [Google Scholar] [CrossRef]
- Ayankojo, I.T.; Morgan, K.T. Increasing air temperatures and its effects on growth and productivity of tomato in South Florida. Plants 2020, 9, 1245. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Thiele, B.; Watt, M.; Kraska, T.; Ulbrich, A.; Kuhn, A.J. Effects of root cooling on plant growth and fruit quality of cocktail tomato during two consecutive seasons. J. Food Qual. 2019, 2019, 3598172. [Google Scholar] [CrossRef]
- Scheelbeek, P.F.; Bird, F.A.; Tuomisto, H.L.; Green, R.; Harris, F.B.; Joy, E.J.M.; Chalabi, Z.; Allen, E.; Haines, A.; Dangour, A.D. Effect of environmental changes on vegetable and legume yields and nutritional quality. Proc. Nat. Acad. Sci. USA 2018, 115, 6804–6809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amini, S.; Maali-Amiri, R.; Kazemi-Shahandashti, S.S.; López-Gómez, M.; Sadeghzadeh, B.; Sobhani-Najafabadi, A.; Kariman, K. Effect of cold stress on polyamine metabolism and antioxidant responses in chickpea. J. Plant Physiol. 2021, 258, 153387. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xu, X.; Wei, D.; Lin, X.; Qiu, S.; Ciampitti, I.; He, P. Soybean yield, nutrient uptake and stoichiometry under different climate regions of northeast China. Sci. Rep. 2020, 10, 8431. [Google Scholar] [CrossRef]
- Menzel, C.M. Temperature has a greater effect on fruit growth than defoliation or fruit thinning in strawberries in the subtropics. Agriculture 2019, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, A.; Shaji, S.; Beena, R.; Sarada, S.; Rani, T.S.; Stephen, R.; Manju, R.V.; Viji, M.M. High temperature induced changes in quality and yield parameters of tomato (Solanum lycopersicum L.) and similarity coefficients among genotypes using SSR markers. Heliyon 2021, 7, e05988. [Google Scholar] [CrossRef]
- Nishad, A.; Mishra, A.N.; Chaudhari, R.; Aryan, R.K.; Katiyar, P. Effect of temperature on growth and yield of rice (Oryza sativa L.) cultivars. Int. J. Chem. Stud. 2018, 6, 1381–1383. [Google Scholar]
- Szczerba, A.; Płażek, A.; Pastuszak, J.; Kopeć, P.; Hornyák, M.; Dubert, F. Effect of low-temperature on germination, growth, and seed yield of four soybean (Glycine max L.) cultivars. Agronomy 2021, 11, 800. [Google Scholar] [CrossRef]
- Savitch, L.V.; Ivanov, A.G.; Gudynaite-Savitch, L.; Huner, N.P.; Simmonds, J. Effects of low-temerature stress on excitation energy partitioning and photoprotection in Zea mays. Funct. Plant Biol. 2009, 36, 37–49. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; et al. Effect of low-temperature on chlorophyll biosynthesis and chloroplast biogenesis of rice seedlings during greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhou, Y.; Xiao, J.; Bao, F. Effects of chilling on the structure, function and development of chloroplasts. Front. Plant Sci. 2018, 9, 1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, X.; Qin, W.; Zhang, Z.; Yao, Y.; Xiao, Y.; Zhu, J. Effects of low-temperature on nitrogen metabolism activities and photosynthetic pigments in different cabbage seedlings. J. South. Agric. 2019, 50, 2728–2733. [Google Scholar]
- Li, S.; Yang, Y.; Zhang, Q.; Liu, N.; Xu, Q.; Hu, L. Differential physiological and metabolic response to low-temperature in two zoysiagrass genotypes native to high and low latitude. PLoS ONE 2018, 13, e0198885. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.M.; Kuang, J.F.; Ji, S.J.; Chen, Q.F.; Deng, W.; Min, T.; Shan, W.; Chen, J.Y.; Lu, W.J. The membrane lipid metabolism in horticultural products suffering chilling injury. Food Qual. Saf. 2020, 4, 9–14. [Google Scholar] [CrossRef]
- Theocharis, A.; Clément, C.; Barka, E.A. Physiological and molecular changes in plants grown at low-temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef]
- Maoyan, W. Effects of low-temperature on the nucleic acid content and the nucleases activities in the leaves of maize seedlings. Acta Agric. Boreali Sin. 1989, 4, 28–33. [Google Scholar]
- Göbel, L.; Coners, H.; Hertel, D.; Willinghöfer, S.; Leuschner, C. The role of low soil temperature for photosynthesis and stomatal conductance of three graminoids from different elevations. Front. Plant Sci. 2019, 10, 330. [Google Scholar] [CrossRef] [Green Version]
- Boubakri, H.; Chong, J.; Poutaraud, A.; Schmitt, C.; Bertsch, C.; Mliki, A.; Masson, J.E.; Soustre-Gacougnolle, I. Riboflavin (Vitamin B 2) induces defence responses and resistance to Plasmopara viticola in grapevine. Eur. J. Plant Pathol. 2013, 136, 837–855. [Google Scholar] [CrossRef]
- Korkmaz, A.; Korkmaz, Y.; Demirkıran, A.R. Enhancing chilling stress tolerance of pepper seedlings by exogenous application of 5-aminolevulinic acid. Environ. Exp. Bot. 2010, 67, 495–501. [Google Scholar] [CrossRef]
- Schortemeyer, M.; Evans, J.R.; Bruhn, D.; Bergstrom, D.M.; Ball, M.C. Temperature responses of photosynthesis and respiration in a sub-Antarctic megaherb from Heard Island. Funct. Plant Biol. 2015, 42, 552–564. [Google Scholar] [CrossRef]
- Hsu, C.H.; Hsu, Y.T. Biochemical responses of rice roots to cold stress. Bot. Stud. 2019, 60, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gechev, T.; Petrov, V. Reactive oxygen species and abiotic stress in plants. Int. J. Mol. Sci. 2020, 21, 7433. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Ann. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarropoulou, V.; Dimassi-Theriou, K.; Therios, I.; Koukourikou-Petridou, M. Melatonin enhances root regeneration, photosynthetic pigments, biomass, total carbohydrates and proline content in the cherry rootstock PHL-C (Prunus avium Prunus cerasus). Plant Physiol. Biochem. 2012, 61, 162–168. [Google Scholar] [CrossRef]
- Turk, H.; Erdal, S.; Genisel, M.; Atici, O.; Demir, Y.; Yanmis, D. The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul. 2014, 74, 139–152. [Google Scholar] [CrossRef]
- Najeeb, S.; Ali, J.; Mahender, A.; Pang, Y.L.; Zilhas, J.; Murugaiyan, V.; Vemireddy; Lakshminarayana, R.; Li, Z. Identification of main-effect quantitative trait loci (QTLs) for low-temperature stress tolerance germination- and early seedling vigor-related traits in rice (Oryza sativa L.). Mol. Breed. 2020, 40, 10. [Google Scholar] [CrossRef] [Green Version]
- Cong Dien, D.; Yamakawa, T. Phenotypic variation and selection for cold-tolerant rice (Oryza sativa L.) at germination and seedling stages. Agriculture 2019, 9, 162. [Google Scholar] [CrossRef] [Green Version]
- Bezirganoglu, I.; Uysal, P.; Yiğit, O.R. Cold stress resistance and the antioxidant enzyme system in Pisum sativum. J. Anim. Plant Sci. 2018, 28, 561–567. [Google Scholar]
- Kumar, S.; Nayyar, H.; Bhanwara, R.K.; Upadhyaya, H.D. Chilling stress effects on reproductive biology of chickpea. J. SAT Agric. Res. 2010, 8, 1–4. [Google Scholar]
- Kumar, S.; Malik, J.; Thakur, P.; Kaistha, S.; Sharma, K.D.; Upadhyaya, H.D.; Berger, J.D.; Nayyar, H. Growth and metabolic responses of contrasting chickpea (Cicer arietinum L.) genotypes to chilling stress at reproductive phase. Acta Physiol. Plant. 2011, 33, 779–787. [Google Scholar] [CrossRef]
- Nayyar, H.; Kaur, G.; Kumar, S.; Upadhyaya, H.D. Low-temperature effects during seed filling on chickpea genotypes (Cicer arietinum L.): Probing mechanisms affecting seed reserves and yield. J. Agron. Crop Sci. 2007, 193, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, X.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Alleviation of field low-temperature stress in winter wheat by exogenous application of salicylic acid. J. Plant Growth Regul. 2021, 40, 811–823. [Google Scholar] [CrossRef]
- Gámez-Vázquez, A.J.; Avila-Perches, M.A.; Virgen-Vargas, J.; Ruiz-Torres, N.A.; Gámez-Vázquez, F.P.; Ascencio-Álvarez, A. Physiological quality of the seed and maize seedlings development at low temperatures. Rev. Mex. Cienc. Agrícolas 2015, 6, 1769–1779. [Google Scholar]
- Kayess, M.O.; Hassan, M.M.; Nurhasan, M.; Ahmed, K. Effect of low-temperature on chlorophyll and carotenoid content on the seedlings of some selected boro rice varieties. Am. J. Plant Sci. 2020, 11, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Ntatsi, G.; Savvas, D.; Kläring, H.P.; Schwarz, D. Growth, yield, and metabolic responses of temperature-stressed tomato to grafting onto rootstocks differing in cold tolerance. J. Am. Soc. Hortic. Sci. 2014, 139, 230–243. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Luo, W.; Ji, R.; Xu, Y.; Xu, G.; Qiu, S.; Tang, H.A. Comparative proteomic study of cold responses in potato leaves. Heliyon 2021, 7, e06002. [Google Scholar] [CrossRef]
- Rodríguez, V.M.; Soengas, P.; Alonso-Villaverde, V.; Sotelo, T.; Cartea, M.E.; Velasco, P. Effect of temperature stress on the early vegetative development of Brassica oleracea L. BMC Plant Biol. 2015, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.H.; Koo, S.S.; Oh, H.T.; Lee, E.S.; Park, J.H.; Phan, K.A.T.; Wi, S.D.; Bae, S.B.; Paeng, S.K.; Chae, H.B.; et al. The physiological functions of universal stress proteins and their molecular mechanism to protect plants from environmental stresses. Front. Plant Sci. 2019, 10, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhao, X.; Liu, S.; Sun, F.; Zhang, C.; Xi, Y. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, J.; Wang, W.; Sun, Y. Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress. Photosynthetica 2016, 54, 19–27. [Google Scholar] [CrossRef]
- Ahmad, S.; Su, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Javed, T.; Han, Q. Foliar application of melatonin delay leaf senescence in maize by improving the antioxidant defense system and enhancing photosynthetic capacity under semi-arid regions. Protoplasma 2020, 257, 1079–1092. [Google Scholar] [CrossRef]
- Ahmad, S.; Cui, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Su, W.; Javed, T.; El-Serehy, H.A.; Jia, Z.; et al. Exogenous application of melatonin induces tolerance to salt stress by improving the photosynthetic efficiency and antioxidant defense system of maize seedling. J. Plant Growth Regul. 2020, 19, 1–4. [Google Scholar] [CrossRef]
- Kamran, M.; Wennan, S.; Ahmad, I.; Xiangping, M.; Wenwen, C.; Xudong, Z.; Siwei, M.; Khan, A.; Qingfang, H.; Tiening, L. Application of paclobutrazol affect maize grain yield by regulating root morphological and physiological characteristics under a semiarid region. Sci. Rep. 2018, 8, 4818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metwali, E.M.; Abdelmoneim, T.S.; Bakheit, M.A.; Kadasa, N.M. Alleviation of salinity stress in faba bean (Vicia faba L.) plants by inoculation with plant growth promoting rhizobacteria (PGPR). Plant Omics 2015, 8, 449. [Google Scholar]
- Pfannschmidt, T.; Munné-Bosch, S. Plastid signaling during the plant life cycle. In Plastid Development in Leaves During Growth and Senescence; Springer: Berlin, Germany, 2013; pp. 503–528. [Google Scholar]
- Saleem, M.; Fariduddin, Q.; Janda, T. Multifaceted role of salicylic acid in combating cold stress in plants: A review. J. Plant Growth Regul. 2021, 40, 464–485. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Ahmad, I.; Hellebust, J.A. The Relationship between inorganic nitrogen metabolism and proline accumulation in osmoregulatory responses of two Euryhaline Microalgae. J. Plant Physiol. 1988, 88, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Gavelienė, V.; Pakalniškytė, L.; Novickienė, L. Regulation of proline and ethylene levels in rape seedlings for freezing tolerance. Open Life Sci. 2014, 9, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, J.K.; Munshi, A.D.; Kumar, R.; Pandey, R.N.; Arora, A.; Bhat, J.S.; Sureja, A.K. Effect of salt stress on cucumber: Na+–K+ ratio, osmolyte concentration, phenols and chlorophyll content. Acta Physiol. Plant. 2010, 32, 103–114. [Google Scholar] [CrossRef]
- Maharajan, T.; Ceasar, S.A.; Ajeesh krishna, T.P.; Ramakrishnan, M.; Duraipandiyan, V.; Naif Abdulla, A.D.; Ignacimuthu, S. Utilization of molecular markers for improving the phosphorus efficiency in crop plants. Plant Breed. 2018, 137, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Beheshti, M.; Etesami, H.; Alikhani, H.A. Interaction study of biochar with phosphate-solubilizing bacterium on phosphorus availability in calcareous soil. Arch. Agron. Soil Sci. 2017, 60, 1572–1581. [Google Scholar] [CrossRef]
- Lobo, C.B.; Juárez Tomása, M.S.; Viruel, E.; Ferrero, M.A.; Lucca, M.E. Development of low cost formulations of plant growth-promoting bacteria to be used as inoculants in beneficial agricultural technologies. Microbiol. Res. 2019, 219, 12–25. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana; Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 171–190. [Google Scholar]
- Mukhtar, S.; Shahid, I.; Mehnaz, S.; Malik, K.A. Assessment of two carrier materials for phosphate solubilizing biofertilizers and their effect on growth of wheat (Triticum aestivum L.). Microbiol. Res. 2017, 205, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Rychter, A.M.; Rao, I.M.; Cardoso, J.A. Role of phosphorus in photosynthetic carbon assimilation and partitioning. In Handbook of Photosynthesis; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Takahashi, S.; Anwar, M.R. Wheat grain yield, phosphorus uptake and soil phosphorus fraction after 23 years of annual fertilizer application to an Andosol. Field Crop. Res. 2007, 101, 160–171. [Google Scholar] [CrossRef]
- Solankia, M.; Kundub, B.S.; Nehrac, K. Molecular diversity of phosphate solubilizing bacteria isolated from the rhizosphere of chickpea, mustard and wheat. Ann. Agrar. Sci. 2018, 16, 458–463. [Google Scholar] [CrossRef]
- Wei, Y.; Zhao, Y.; Shi, M.; Cao, Z.; Lu, Q.; Yang, T.; Fan, Y.; Wei, Z. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Biores. Technol. 2018, 247, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Sabir, M.; Farooq, M.; Maqsood, M.A.; Ahmad, H.R.; Warraich, E.A. Phosphorus deficiency in plants: Responses, adaptive mechanisms, and signaling. In Plant Signaling: Understanding the Molecular Crosstalk; Springer: New Delhi, India, 2014; pp. 133–148. [Google Scholar]
- Blanco-Vargas, A.; Rodríguez-Gacha, L.M.; Sánchez-Castro, N.; Herrera-Carlosama, L.; Poutou-Piñales, R.A.; Díaz-Ariza, L.A.; Gutiérrez-Romero, V.; Rivera-Hoyos, C.M.; Ardila-Leal, L.D.; Pedroza-Rodriguez, A.M. Bioinoculant production composed by Pseudomonas sp., Serratia sp., and Kosakonia sp., preliminary effect on Allium cepa L., growth at plot scale. Univ. Sci. 2021, 26, 79–118. [Google Scholar] [CrossRef]
- Islam, M.K.; Sano, A.; Majumder, M.S.I.; Hossain, M.A.; Sakagami, J.I. Isolation and molecular characterization of phosphate solubilizing filamentous fungi from subtropical soils in Okinawa. Appl. Ecol. Environ. Res. 2019, 17, 9145–9157. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [Green Version]
- Khabarov, N.; Obersteiner, M. Global phosphorus fertilizer market and national policies: A case study revisiting the 2008 price peak. Front. Nutr. 2017, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Batool, S.; Iqbal, A. Phosphate solubilizing rhizobacteria as alternative of chemical fertilizer for growth and yield of Triticum aestivum (Var. Galaxy 2013). Saudi J. Biol. Sci. 2019, 26, 1400–1410. [Google Scholar] [CrossRef]
- Praveen, K.V.; Singh, A. Realizing the potential of a low-cost technology to enhance crop yields: Evidence from a meta-analysis of biofertilizers in India. Agric. Econ. Res. Rev. 2019, 32, 77–91. [Google Scholar] [CrossRef]
- Haouas, A.; El Modafar, C.; Douira, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; Moukhli, A.; Amir, S. Alcaligenes aquatilis GTE53: Phosphate solubilizing and bioremediation bacterium isolated from new biotope “phosphate sludge enriched-compost”. Saudi J. Biol. Sci. 2021, 28, 371–379. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as biofertilizers, a potential approach for sustainable crop production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Asoegwu, C.R.; Awuchi, C.G.; Nelson, K.; Orji, C.G.; Nwosu, O.U.; Egbufor, U.C.; Awuchi, C.G. A review on the role of biofertilizers in reducing soil pollution and increasing soil nutrients. Himal. J. Agric. 2020, 1, 34–38. [Google Scholar]
- Sammauria, R.; Kumawat, S.; Kumawat, P.; Singh, J.; Jatwa, T.K. Microbial inoculants: Potential tool for sustainability of agricultural production systems. Arch. Microbiol. 2020, 202, 677–693. [Google Scholar] [CrossRef]
- Trivedi, P.; Pandey, A.; Palni, L.M.S. Bacterial inoculants for field applications under Mountain Ecosystem: Present initiatives and future prospects. In Bacteria in Agrobiology: Plant Probiotics; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 15–44. [Google Scholar]
- Pandey, A.; Jain, R.; Sharma, A.; Dhakar, K.; Kaira, G.S.; Rahi, P. 16S rRNA gene sequencing and MALDI-TOF mass spectrometry based comparative assessment and bioprospection of psychrotolerant bacteria isolates from high altitudes under mountain ecosystem. SN Appl. Sci. 2019, 1, 278. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.; Pandey, A. A phenazine-1 carboxylic acid producing polyextremophilic Pseudomonas chlororaphis (MCC2693) strain, isolated from mountain ecosystem, possesses biocontrol and plant growth promotion abilities. Microbiol. Res. 2016, 190, 63–71. [Google Scholar] [CrossRef]
- Yarzábal, L.A. Cold-tolerant phosphate-solubilizing microorganisms and agriculture development in mountainous regions of the world. In Phosphate Solubilizing Microorganisms; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Cham, Switzerland, 2014; pp. 113–135. [Google Scholar]
- Gupta, R.; Kumari, A.; Noureldeen, A.; Darwish, H. Rhizosphere mediated growth enhancement using phosphate solubilizing rhizobacteria and their tri-calcium phosphate solubilization activity under pot culture assays in rice (Oryza sativa). Saudi J. Biol. Sci. 2021, 28, 3692–3700. [Google Scholar] [CrossRef] [PubMed]
- Cumpa-Velásquez, L.M.; Moriconi, J.I.; Dip, D.P.; Castagno, L.N.; Puig, M.L.; Maiale, S.J.; Santa-María, G.E.; Sannazzaro, A.I.; Estrella, M.J. Prospecting phosphate solubilizing bacteria in alkaline-sodic environments reveals intra-specific variability in Pantoea eucalypti affecting nutrient acquisition and rhizobial nodulation in Lotus tenuis. Appl. Soil Ecol. 2021, 168, 104125. [Google Scholar] [CrossRef]
- Hassimi, M.S.; Hamdali, H.; Ouhdouch, Y.; Pinelli, E.; Merlina, G.; Claude, R.J.; Hafidi, M. Moroccan rock phosphate solubilization during a thermo-anaerobic grassland waste biodegradation process. Afr. J. Biotechnol. 2013, 12, 6859–6865. [Google Scholar]
- Yadav, K.; Kumar, C.; Archana, G.; Kumar, N. Pseudomonas fluorescens ATCC 13525 containing an artificial oxalate operon and vitreoscilla hemoglobin secretes oxalic acid and solubilizes rock phosphate in acidic alfisols. PLoS ONE 2014, 9, e92400. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture—A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, S. Identification and characterization of the phosphate-solubilizing bacterium Pantoea sp. S32 in reclamation soil in Shanxi, China. Front. Microbiol. 2019, 10, 2171. [Google Scholar] [CrossRef] [Green Version]
- Pande, A.; Pandey, P.; Mehra, S.; Singh, M.; Kaushik, S. Phenotypic and genotypic characterization of phosphate solubilizing bacteria and their efficiency on the growth of maize. J. Gen. Eng. Biotechnol. 2017, 15, 379–391. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Ahmad, E. Mechanism of phosphate solubilization and physiological functions of phosphate-solubilizing microorganisms. In Phosphate Solubilizing Microorganisms; Springer: Cham, Switzerland, 2014; pp. 31–62. [Google Scholar]
- De Amaral Leite, A.; de Souza Cardoso, A.A.; de Almeida Leite, R.; de Oliveira-Longatti, S.M.; Lustosa Filho, J.F.; de Souza Moreira, F.M.; Melo, L.C.A. Selected bacterial strains enhance phosphorus availability from biochar-based rock phosphate fertilizer. Ann. Microbiol. 2020, 70, 1–3. [Google Scholar] [CrossRef]
- Suleman, M.; Yasmin, S.; Rasul, M.; Yahya, M.; Atta, B.M.; Mirza, M.S. Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLoS ONE 2018, 13, e0204408. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yang, F.; Zhang, L.; Wang, J. Organic acid secretion and phosphate solubilizing efficiency of Pseudomonas sp. PSB12: Effects of phosphorus forms and carbon sources. Geomicrobiol. J. 2016, 33, 870–877. [Google Scholar] [CrossRef]
- Lessie, T.G.; Phibbs, P.V. Alternative pathways of carbohydrate utilization in pseudomonads. Annu. Rev. Microbiol. 1984, 38, 359–388. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.H.; Rogers, R.D.; Mead, G. Mining by microbe. Bio/Technology 1993, 11, 1250–1254. [Google Scholar] [CrossRef]
- Cunningham, J.E.; Kuiack, C. Production of citric and oxalic acids and solubilization of calcium phosphate by Penicillium bilaii. Appl. Environ. Microbiol. 1992, 58, 1451–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, A.F. Influence of Streptomyces sp. Kp109810 on Solubilization of inorganic phosphate and growth of maize (Zea mays L.). J. Appl. Plant Prot. 2020, 9, 17–24. [Google Scholar] [CrossRef]
- Surapat, W.; Pukahuta, C.; Rattanachaikunsopon, P.; Aimi, T.; Boonlue, S. Characteristics of phosphate solubilization by phosphate-solubilizing bacteria isolated from agricultural chili soil and their efficiency on the growth of chili (Capsicum frutescens L. cv. Hua Rua). Chiang Mai J. Sci. 2013, 40, 11–25. [Google Scholar]
- Asea, P.E.A.; Kucey, R.M.N.; Stewart, J.W.B. Inorganic phosphate solubilization by two Penicillium species in solution culture and soil. Soil Biol. Biochem. 1988, 20, 459–464. [Google Scholar] [CrossRef]
- Illmer, P.; Schinner, F. Solubilization of inorganic phosphates by microorganisms isolated from forest soils. Soil Biol. Biochem. 1992, 24, 389–395. [Google Scholar] [CrossRef]
- Kim, K.Y.; McDonald, G.A.; Jordan, D. Solubilization of hydroxyapatite by Enterobacter agglomerans and cloned Escherichia coli in culture medium. Biol. Fertil. Soils 1997, 24, 347–352. [Google Scholar] [CrossRef]
- Borgi, M.A.; Saidi, I.; Moula, A.; Rhimi, S.; Rhimi, M. The attractive Serratia plymuthica BMA1 strain with high rock phosphate-solubilizing activity and its effect on the growth and phosphorus uptake by Vicia faba L. plants. Geomicrobiol. J. 2020, 37, 437–445. [Google Scholar] [CrossRef]
- Chawngthu, L.; Hnamte, R.; Lalfakzuala, R. Isolation and characterization of rhizospheric phosphate solubilizing bacteria from wetland paddy field of Mizoram, India. Geomicrobiol. J. 2020, 37, 366–375. [Google Scholar] [CrossRef]
- Rfaki, A.; Zennouhi, O.; Aliyat, F.Z.; Nassiri, L.; Ibijbijen, J. Isolation, selection and characterization of root-associated rock phosphate solubilizing bacteria in Moroccan wheat (Triticum aestivum L.). Geomicrobiol. J. 2020, 37, 230–241. [Google Scholar] [CrossRef]
- Azaroual, S.E.; Hazzoumi, Z.; Mernissi, N.E.; Aasfar, A.; Meftah Kadmiri, I.; Bouizgarne, B. Role of inorganic phosphate solubilizing bacilli isolated from Moroccan phosphate rock mine and rhizosphere soils in wheat (Triticum aestivum L.) phosphorus uptake. Curr. Microbiol. 2020, 77, 2391–2404. [Google Scholar] [CrossRef]
- Zaheer, A.; Malik, A.; Sher, A.; Qaisrani, M.M.; Mehmood, A.; Khan, S.U.; Ashraf, M.; Mirza, Z.; Karim, S.; Rasool, M. Isolation, characterization, and effect of phosphate-zinc-solubilizing bacterial strains on chickpea (Cicer arietinum L.) growth. Saudi J. Biol. Sci. 2019, 26, 1061–1067. [Google Scholar] [CrossRef]
- Gulati, A.; Sharma, N.; Vyas, P.; Sood, S.; Rahi, P.; Pathania, V.; Prasad, R. Organic acid production and plant growth promotion as a function of phosphate solubilization by Acinetobacter rhizosphaerae strain BIHB 723 isolated from the cold deserts of the trans-Himalayas. Arch. Microbiol. 2010, 192, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, P.; Sa, T. Pseudomonas corrugata (NRRL B-30409) mutants increased phosphate solubilization, organic acid production and plant growth at lower temperatures. Curr. Microbiol. 2008, 56, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Vinod, K.; Prashant, S.; Punesh, S.; Verma, A.K.; Sanjeev, A. Phytase producing bacteria from Himalayan soil and their tricalcium phosphate solubilizing abilities. Ann. Biol. 2011, 27, 121–128. [Google Scholar]
- Zavaleta-Pastor, M.; Sohlenkamp, C.; Gao, J.L.; Guan, Z.; Zaheer, R.; Finan, T.M.; Raetz, C.R.H.; López-Lara, I.M.; Geiger, O. Sinorhizobium meliloti phospholipase C required for lipid remodeling during phosphorus limitation. Proc. Nat. Acad. Sci. USA 2010, 107, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Shulse, C.N.; Chovatia, M.; Agosto, C.; Wang, G.; Hamilton, M.; Deutsch, S.; Yoshikuni, Y.; Blow, M.J. Engineered root bacteria release plant-available phosphate from phytate. Appl. Environ. Microbiol. 2019, 85, e01210–e01219. [Google Scholar] [CrossRef] [Green Version]
- Behera, B.C.; Yadav, H.; Singh, S.K.; Mishra, R.R.; Sethi, B.K.; Dutta, S.K.; Thatoi, H.N. Phosphate solubilization and acid phosphatase activity of Serratia sp. isolated from mangrove soil of Mahanadi river delta, Odisha, India. J. Gen. Eng. Biotechnol. 2017, 15, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, A.; Khan, M.S.; Ahemad, M.; Oves, M.; Wani, P.A. Recent advances in plant growth promotion by phosphate-solubilizing microbes. In Microbial Strategies for Crop Improvement; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 23–50. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi—Current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Shahid, M.; Ahmed, B.; Khan, M.S. Evaluation of microbiological management strategy of herbicide toxicity to greengram plants. Biocatal. Agric. Biotechnol. 2018, 14, 96–108. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, N.; Yadav, A.N. Bioprospecting of phosphorus solubilizing bacteria from Renuka Lake ecosystems, lesser Himalayas. J. Appl. Biol. Biotechnol. 2019, 7, 1–6. [Google Scholar]
- Verma, P.; Yadav, A.N.; Kazy, S.K.; Saxena, A.K.; Suman, A. Evaluating the diversity and phylogeny of plant growth promoting bacteria associated with wheat (Triticum aestivum) growing in central zone of India. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 432–447. [Google Scholar]
- Katiyar, V.; Goel, R. Siderophore mediated plant growth promotion at low-temperature by mutant of fluorescent Pseudomonads. Plant Growth Regul. 2004, 42, 239–244. [Google Scholar] [CrossRef]
- Selvakumar, G.; Joshi, P.; Suyal, P.; Mishra, P.K.; Joshi, G.K.; Bisht, J.K.; Bhatt, J.C.; Gupta, H.S. Pseudomonas lurida M2RH3 (MTCC 9245), a psychrotolerant bacterium from the Uttarakhand Himalayas, solubilizes phosphate and promotes wheat seedling growth. World J. Microbiol. Biotechnol. 2011, 27, 1129–1135. [Google Scholar] [CrossRef]
- Rajaee, S.; Alikhani, H.A.; Raiesi, F. Effect of plant growth promoting potentials of Azotobacter chroococcum native strains on growth, yield and uptake of nutrients in wheat. JWSS Isfahan Univ. Technol. 2017, 11, 285–297. [Google Scholar]
- Gautam, N.; Sharma, P.; Rana, J.C.; Singh, M. Characterization of Bacillus weihenstephanensis AGII: A psychrotolerant bacteria isolated from rhizosphere of medicinal plant Gentiana kurroo Royle. Acta Sci. Agric. 2021, 5, 52–57. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Saxena, P.; Sharma, A.; Srivastava, R.; Jamali, H.; Bharati, A.P.; Yadav, J.; Srivastava, A.K.; Kumar, M.; Chakdar, H. Draft genome sequence of a cold-adapted phosphorous-solubilizing Pseudomonas koreensis P2 isolated from Sela Lake, India. 3 Biotech 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yarzábal, L.A.; Monserrate, L.; Buela, L.; Chica, E. Antarctic Pseudomonas spp. promote wheat germination and growth at low-temperatures. Polar Biol. 2018, 41, 2343–2354. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Ojeda, P.; Ogata-Gutiérrez, K.; Zúñiga-Dávila, D. Evaluation of plant growth promoting activity and heavy metal tolerance of psychrotrophic bacteria associated with maca (Lepidium meyenii Walp.) rhizosphere. AIMS Microbiol. 2017, 3, 279. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Saxena, A.K. Bioprospecting of plant growth promoting psychrotrophic bacilli from the cold desert of north western Indian Himalayas. Ind. J. Exp. Biol. 2016, 54, 142–150. [Google Scholar]
- Gusain, Y.S.; Kamal, R.; Mehta, C.M.; Singh, U.S.; Sharma, A.K. Phosphate solubilizing and indole-3-acetic acid producing bacteria from the soil of Garhwal Himalaya aimed to improve the growth of rice. J. Environ. Biol. 2015, 36, 301. [Google Scholar]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Panjiar, N.; Kumar, S.; Saxena, A.K.; Suman, A. Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann. Microbiol. 2015, 65, 1885–1899. [Google Scholar] [CrossRef]
- Beatriz Sánchez, D.; Gómez, R.M.; García, A.M.; Bonilla, R.R. Phosphate solubilizing bacteria isolated from Pennisetum clandestinum associate to livestock systems in the andean area. Rev. UDCA Actual. Divulg. Cient. 2014, 17, 423–431. [Google Scholar] [CrossRef]
- Berríos, G.; Cabrera, G.; Gidekel, M.; Gutiérrez-Moraga, A. Characterization of a novel Antarctic plant growth-promoting bacterial strain and its interaction with Antarctic hair grass (Deschampsia antarctica Desv). Polar Biol. 2013, 36, 349–362. [Google Scholar] [CrossRef]
- Selvakumar, G.; Kundu, S.; Joshi, P.; Nazim, S.; Gupta, A.D.; Gupta, H.S. Growth promotion of wheat seedlings by Exiguobacterium acetylicum 1P (MTCC 8707) a cold tolerant bacterial strain from the Uttarakhand Himalayas. Ind. J. Microbiol. 2010, 50, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Prithiviraj, B.; Charles, T.C.; Driscoll, B.T.; Smith, D.L. Low-temperature tolerant Bradyrhizobium japonicum strains allowing improved nodulation and nitrogen fixation of soybean in a short season (cool spring) area. Eur. J. Agron. 2003, 19, 205–213. [Google Scholar] [CrossRef]
- Selvakumar, G.; Kundu, S.; Joshi, P.; Nazim, S.; Gupta, A.D.; Mishra, P.K.; Gupta, H.S. Characterization of a cold tolerant plant growth-promoting bacterium Pantoea dispersa 1A isolated from a sub-alpine soil in the North Western Indian Himalayas. World J. Microbiol. Biotechnol. 2008, 24, 955–960. [Google Scholar] [CrossRef]
- Kumar, S.; Suyal, D.C.; Dhauni, N.; Bhoriyal, M.; Goel, R. Relative plant growth promoting potential of Himalayan Psychrotolerant Pseudomonas jesenii strain MP1 against native Cicer arietinum (L.)., Vigna mungo (L.) Hepper; Vigna radiata (L.) Wilczek., Cajanus cajan (L.) Millsp. and Eleusine coracana (L.) Gaertn. Afr. J. Microbiol. Res. 2014, 8, 3931–3943. [Google Scholar]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Sidhu, G.P.S.; Kumar, R.; Kohli, S.K.; Yadav, P.; Kapoor, D.; Bali, A.S.; Shahzad, B.; Khanna, K. Abiotic stress management in plants: Role of ethylene. Mol. Plant Abiotic Stress Biol. Biotechnol. 2019, 185–208. [Google Scholar] [CrossRef]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environ. Exp. Bot. 2020, 178, 104124. [Google Scholar] [CrossRef]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The pivotal role of ethylene in plant growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Del Carmen Orozco-Mosqueda, M.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Cheng, Z.; Park, E.; Glick, B.R. 1-Aminocyclopropane-1- carboxylate (ACC) deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Can. J. Microbiol. 2007, 53, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Yarzábal, L.A.; Chica, E. Potential for developing low-input sustainable agriculture in the tropical Andes by making use of native microbial resources. In Plantmicrobe Interactions in Agro-Ecological Perspectives; Singh, D., Singh, H., Prabha, R., Eds.; Springer: Singapore, 2017; Volume 2, pp. 29–54. [Google Scholar]
- Pandey, A.; Yarzábal, L.A. Bioprospecting cold-adapted plant growth promoting microorganisms from mountain environments. Appl. Microbiol. Biotechnol. 2019, 103, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Melo, I.S.; Souza, W.R.; Silva, L.J.; Santos, S.N.; Assalin, M.A.; Zucchi, T.D.; Queiroz, S.C.N. Antifungal activity of Pseudomonas frederiksbergensis CMAA 1323 isolated from the Antarctic hair grass Deschampsia antarctica. Br. Microbiol. Res. J. 2016, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Yan, F.; Wilson, C.; Shen, Q.; Zheng, X. The ability of a cold adapted Rhodotorula mucilaginosa strain from Tibet to control blue mold in pear fruit. Antonie van Leeuwenhoek 2015, 108, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.C.; Sethi, B.K.; Mishra, R.R.; Dutta, S.K.; Thatoi, H.N. Microbial cellulases–Diversity qnd biotechnology with reference to mangrove environment: A review. J. Gen. Eng. Biotechnol. 2017, 15, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Ding, H.; Yu, Y.; Chen, B. A cold-adapted chitinase-producing bacterium from Antarctica and its potential in biocontrol of plant pathogenic fungi. Mar. Drugs 2019, 17, 695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremer, R.J.; Souissi, T. Cyanide production by rhizobacteria and potential for suppression of wheat seedling growth. Curr. Microbiol. 2001, 43, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H. Cyanide-resistant respiration: A non-phosphorylating electron transport pathway acts as an energy overflow. Physiol. Plant. 1982, 55, 478–485. [Google Scholar] [CrossRef]
- Sarniguet, A.; Lucas, P.; Lucas, M.; Samson, R. Soil conduciveness to take all of wheat: Influence of the nitrogen fertilizers on the structure of populations of fluorescent pseudomonads. Plant Soil 1992, 145, 29–36. [Google Scholar] [CrossRef]
- Meena, V.S.; Maurya, B.R.; Verma, J.P. Does a rhizospheric microorganism enhance K+ availability in agricultural soils? Microbiol. Res. 2014, 169, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, A.C.; da Costa Rachid, C.T.C.; de Jesus, H.E.; Rosado, A.S.; Peixoto, R.S. Predicting the biotechnological potential of bacteria isolated from Antarctic soils, including the rhizosphere of vascular plants. Polar Biol. 2017, 40, 1393–1407. [Google Scholar] [CrossRef]
- Canion, A.; Prakash, O.; Green, S.J.; Jahnke, L.; Kuypers, M.M.; Kostka, J.E. Isolation and physiological characterization of psychrophilic denitrifying bacteria from permanently cold Arctic ford sediments (Svalbard, Norway). Environ. Microbiol. 2013, 15, 1606–1618. [Google Scholar] [CrossRef] [PubMed]
- De Curtis, F.; Lima, G.; Vitullo, D.; De Cicco, V. Biocontrol of Rhizoctonia solani and Sclerotium rolfsii on tomato by delivering antagonistic bacteria through a drip irrigation system. Crop Prot. 2010, 29, 663–670. [Google Scholar] [CrossRef]
- Barrientos-Diaz, L.; Gidekel, M.; Gutierrez, A. Characterization of rhizospheric bacteria isolated from Deschampsia antarctica Desv. World J. Microbiol. Biotechnol. 2008, 24, 2289–2296. [Google Scholar] [CrossRef]
- Katiyar, V.; Goel, R. Solubilization of inorganic phosphate and plant growth promotion by cold tolerant mutants of Pseudomonas fluorescens. Microbiol. Res. 2003, 158, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.K.; Mishra, S.; Bisht, S.C.; Selvakumar, G.; Kundu, S.; Bisht, J.K.; Gupta, H.S. Isolation, molecular characterization and growth-promotion activities of a cold tolerant bacterium Pseudomonas sp. NARs9 (MTCC9002) from the Indian Himalayas. Biol. Res. 2009, 42, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, B.; Trivedi, P.; Pandey, A. Pseudomonas corrugata: A suitable bioinoculant for maize grown under rainfed conditions of Himalayan region. Soil Biol. Biochem. 2007, 39, 3093–3100. [Google Scholar] [CrossRef]
- Pandey, A.; Trivedi, P.; Kumar, B.; Palni, L.M.S. Characteristics of a phosphate solubilizing and antagonistic strain of Pseudomonas putida (B0) isolated from a sub-alpine location in the Indian central Himalaya. Curr. Microbiol. 2006, 53, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Pandey, A. Low-temperature phosphate solubilization and plant growth promotion by psychrotrophic bacteria, isolated from Indian Himalayan Region. Res. J. Microbiol. 2007, 2, 454–461. [Google Scholar]
- Saranya, K.; Sundaramanickam, A.; Shekhar, S.; Meena, M.; Sathishkumar, R.S.; Balasubramanian, T. Biosorption of multi-heavy metals by coral associated phosphate solubilising bacteria Cronobacter muytjensii KSCAS2. J. Environ. Manag. 2018, 222, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, A.; Ahmed, B.; Zaidi, A.; Khan, M.S. Bioreduction of toxicity influenced by bioactive molecules secreted under metal stress by Azotobacter chroococcum. Ecotoxicology 2019, 28, 302–322. [Google Scholar] [CrossRef]
- Oves, M.; Khan, M.S.; Qari, H.A. Chromium-reducing and phosphate-solubilizing Achromobacter xylosoxidans bacteria from the heavy metal-contaminated soil of the Brass city, Moradabad, India. Int. J. Environ. Sci. Technol. 2019, 16, 6967–6984. [Google Scholar] [CrossRef]
- Shao, W.; Li, M.; Teng, Z.; Qiu, B.; Huo, Y.; Zhang, K. Effects of Pb (II) and Cr (VI) stress on phosphate-solubilizing bacteria (Bacillus sp. strain mrp-3): Oxidative stress and bioaccumulation potential. Int. J. Environ. Res. Public Health 2019, 16, 2172. [Google Scholar] [CrossRef] [Green Version]
- Paul, D.; Sinha, S.N. Isolation and characterization of a phosphate solubilizing heavy metal tolerant bacterium from River Ganga, West Bengal, India. Songklanakarin J. Sci. Technol. 2015, 37, 651–657. [Google Scholar]
- Rajwar, J.; Chandra, R.; Suyal, D.C.; Tomer, S.; Kumar, S.; Goel, R. Comparative phosphate solubilizing efficiency of psychrotolerant Pseudomonas jesenii MP1 and Acinetobacter sp. ST02 against chickpea for sustainable hill agriculture. Biologia 2018, 73, 793–802. [Google Scholar] [CrossRef]
- Tomer, S.; Suyal, D.C.; Shukla, A.; Rajwar, J.; Yadav, A.; Shouche, Y.; Goel, R. Isolation and characterization of phosphate solubilizing bacteria from Western Indian Himalayan soils. 3 Biotech 2017, 7, 95–100. [Google Scholar] [CrossRef]
- Mishra, P.K.; Mishra, S.; Selvakumar, G.; Bisht, S.C.; Kundu, S.; Bisht, J.K.; Gupta, H.S. Characterization of a psychrotrophic plant growth promoting Pseudomonas PGERs17 (MTCC 9000) isolated from North Western Indian Himalayas. Ann. Microbiol. 2008, 58, 561–568. [Google Scholar] [CrossRef]
- Fraga, R.; Rodríguez, H.; González, T. Transfer of the gene encoding the NapA acid phosphatase of Morganell morganii to a Burkholderia cepacia strain. Acta Biotechnol. 2001, 21, 359–369. [Google Scholar] [CrossRef]
- Rodriguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Fraga-Vidal, R.; Rodriguez, H.M.; De Villegas, T.G. Vector for chromosomal integration of the phoC gene in plant growth-promoting bacteria. In First International Meeting on Microbial Phosphate Solubilization; Velazquez, E.C., Rodríguez-Barrueco, R., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 239–244. [Google Scholar]
- Krishnaraj, P.U.; Goldstein, A.H. Cloning of a Serratia marcescens DNA fragment that induces quinoprotein glucose dehydrogenase-mediated gluconic acid production in Escherichia coli in the presence of stationary phase Serratia marcescens. FEMS Microbiol. Lett. 2001, 205, 215–220. [Google Scholar] [CrossRef]
- Goldstein, A.H. Bacterial solubilization of mineral phosphates: Historical perspectives and future prospects. Am. J. Altern. Agric. 1986, 1, 57–65. [Google Scholar] [CrossRef]
- Goldstein, A.H.; Liu, S.T. Molecular cloning and regulation of a mineral phosphate solubilizing gene from Erwinia herbicola. Bio/Technology 1987, 5, 72–74. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef] [Green Version]
- Sashidhar, B.; Podile, A.R. Mineral phosphate solubilization by rhizosphere bacteria and scope for manipulation of the direct oxidation pathway involving glucose dehydrogenase. J. Appl. Microbiol. 2010, 109, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Jordan, D.; Krishnan, H.B. Expression of genes from Rahnella aquatilis that are necessary for mineral phosphate solubilization in Escherichia coli. FEMS Microbiol. Lett. 1988, 159, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, H.; Rossolini, G.M.; Gonzalez, T.; Li, J.P.; Glick, B.R. Isolation of a gene from Burkholderia cepacia IS-16 encoding a protein that facilitates phosphatase activity. Curr. Microbiol. 2000, 40, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Menezes-Blackburn, D.; Giles, C.; Darch, T.; George, T.S.; Blackwell, M.; Stutter, M.; Shand, C.; Lumsdon, D.; Cooper, P.; Wendler, R. Opportunities for mobilizing recalcitrant phosphorus from agricultural soils: A review. Plant Soil 2018, 427, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Rasul, M.; Yasmin, S.; Suleman, M.; Zaheer, A.; Reitz, T.; Tarkka, M.T.; Islam, E.; Mirza, M.S. Glucose dehydrogenase gene containing phosphobacteria for biofortification of Phosphorus with growth promotion of rice. Microbiol. Res. 2019, 223, 1–12. [Google Scholar] [CrossRef]
- Menezes-Blackburn, D.; Inostroza, N.G.; Gianfreda, L.; Greiner, R.; Mora, M.L.; Jorquera, M.A. Phytase-producing Bacillus sp. inoculation increases phosphorus availability in cattle manure. J. Soil Sci. Plant Nutr. 2016, 16, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Otieno, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, H.; Fraga, R.; Gonzalez, T.; Bashan, Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil 2006, 287, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Vidal, R.H.; Mesa, H.R.; de Villegas, T.G. Chromosomal integration and heterologous expression of the Morganella morganii PhoC gene in Pseudomonas putida N-14. In Sustainable Agriculture: Technology, Planning; Salazar, A., Rios, I., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2010. [Google Scholar]
- Thaller, M.C.; Berlutti, F.; Schippa, S.; Lombardi, G.; Rossolini, G.M. Characterization and sequence of PhoC, the principal phosphate-irrepressible acid phosphatase of Morganella morganii. Microbiology 1994, 140, 1341–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, B.L.; Paphazy, M.J.; Haygarth, P.M.; McKelvie, I.D. Inositol phosphates in the environment. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2002, 357, 449–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Kumar, V.; Agrawal, S. Evaluation of phytase producing bacteria for their plant growth promoting activities. Int. J. Microbiol. 2014, 2014, 426483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosal, S.K.; Mehta, A. Molecular approach to study soil bacterial diversity. In Plant-Growth-Promoting Rhizobacteria (PGPR) and Medicinal Plants, 1st ed.; Egamberdieva, D., Shrivastava, S., Varma, A., Eds.; Springer: New York, NY, USA, 2015; pp. 359–380. [Google Scholar]


{kind=link}
| Crops | Scientific Name | Growth Conditions | Applied Low Temperatures | Crop Responses | References |
|---|---|---|---|---|---|
| Soybean | Glycine max | Growth chamber | 25 °C to 10 °C | At 10 °C germination was completely inhibited; very slow at 15 °C but germinated well at 25 °C; highest cell membrane permeability at 10 °C and 15 °C; at 10 °C, the dehydrogenase activity was highest but -α-amylase was poor at 10 °C; photochemical efficiency was higher in Malaga and Petrina plants germinating at 10 °C and 15 °C than at 25 °C; at a lower temperature, dry weight and number of pods reduced but the number of seeds was higher at 10 °C compared to 15 °C and 25°C; seed weight did not differ among temperatures | [98] |
| Peas | Pisum sativum | Greenhouse | 4 °C to −20 °C | Proline content and activities of antioxidant enzymes such as APX, SOD, and CAT gradually increased at cold acclimation | [118] |
| Chickpea | Cicer arietinum | Field | ≤10 °C | Vegetative growth was reduced and all the phenological stages were delayed; caused vegetative aberrations like chlorosis, necrosis of leaf tips and curling of the whole leaf; damage to reproductive stage involved abscission of juvenile buds and flowers and abortion of pods, pollen development was suppressed and seed formation was inhibited | [119] |
| Pots | <20 °C/<10 °C (day/night) | Decreased chlorophyll content, relative leaf water content, dry weight, and yield features such as pods, seed number, and seed yield; increased electrolyte leakage, reduced total sugars, and starch, poor β-amylase, invertase and sucrose synthase; greater oxidative stress, poor levels of enzymatic antioxidants and reduction in proline and ascorbic acid | [120] | ||
| Greenhouse | 11.7/2.3 °C (day/night) | The chilling conditions increased electrolyte leakage, inhibited chlorophyll formation, decreased sucrose content, the water content in leaves, declined total plant weight, reduced the rate and duration of the seed filling, seed size, seed weight, pods per plant and harvest index, reduced the accumulation of starch, proteins, fats, crude fiber, protein fractions like albumins, globulins, prolamins, and glutelins; also, chilling declined the level of sucrose and enzymes such as starch synthase, sucrose synthase, and invertase significantly in the seeds; minerals such as Ca, P and Fe and amino acids were lowered significantly in the stressed seeds | [121] | ||
| Wheat | Triticum aestivum | Field air temperature control system (FATC) | 5.3 °C to −7.0 °C | Low-temperature stress prolonged the growth period significantly decreased net photosynthetic rate, plant height, and biomass production, and reduced grain yield | [122] |
| Maize | Zea mays | Greenhouse | 8 °C to 4 °C | Reduced germination, increased number of dead seeds, reduced plumule dry weight and radicle, declined the rate of metabolic activity | [123] |
| Rice | Oryza sativa | Greenhouse | 22 °C to 14 °C | Significantly reduced shoot and root growth, physiological attributes, leaf chlorophyll fluorescence associated parameters, and dry matter production | [19] |
| Field conditions | <15 °C | Increased the chlorophyll-a, chlorophyll-b, chlorophyll-a/b ratio, and total chlorophyll, increased proline concentration but decreased carotenoid content | [124] | ||
| Tomato | Lycopersicon esculentum | Greenhouse | 14.6 °C | Suppressed fruit yield, restricted fruit mass, increased soluble carbohydrates, total amino acids, and guaiacol peroxidase activity in roots, leaves, and fruit, and superoxide dismutase in fruit but significantly lower malondialdehyde content | [125] |
| Potato | Solanum tuberosum | Growth chamber | 4 °C/2 °C (day/night) | Soluble protein, MDA, and proline enhanced with low-temperature exposure duration but the chlorophyll content decreased; protein spots (N = 52) identified in proteomic studies were involved in defense response, energy metabolism, photosynthesis, protein degradation, ribosome formation, signal transduction, cell movement, N metabolism, and other physiological processes | [126] |
| Cabbage | Brassica oleracea | Pot trays under controlled conditions | 12 ± 1 °C | Low temperatures affected photosynthesis and fresh weight; stomatal conductance and leaf water content were significantly reduced; plants had smaller but thicker leaves; chilling conditions did not show any reduction in the dry matter | [127] |
| Cold-Active PSB | Ecological Habitat | Organic Acids | References |
|---|---|---|---|
| Pseudomonas sp., Pseudomonas palleroniana, Pseudomonas proteolytica, Pseudomonas azotoformans | Soils from high altitudes in Indian Himalayas | Oxalic, lactic, malic, citric, and succinic acids | [20] |
| Serratia plymuthica | Soils | Gluconic acid | [187] |
| Bacillus, Burkholderia, Paenibacillus sp. | Paddy field | Gluconic oxalic, citric, tartaric, succinic, formic and acetic acid | [188] |
| Pantoea, Pseudomonas, Serratia, and Enterobacter | Wheat rhizosphere | Oxalic, citric, gluconic succinic, and fumaric acids | [189] |
| Bacilli strains | Wheat rhizospheres and rock phosphate mine | Gluconic, lactic, citric, malic, succinic and propionic acids | [190] |
| Pseudomonas sp. strain AZ5, Bacillus sp. strain AZ17 | Chickpea rhizosphere | Acetic, oxalic and gluconic acids, acetic, citric, and lactic acids | [191] |
| Pseudomonas | Glacial ice samples | Gluconic acid | [18] |
| Rahnella sp. BIHB 783 | Hippophae rhamnoides rhizosphere | Gluconic, citric, and isocitric acids | [44] |
| Acinetobacter rhizosphaerae strain BIHB 723 | Cold deserts of the trans-Himalayas | Gluconic, oxalic, 2-keto gluconic, lactic, malic, and formic acids | [192] |
| Fluorescent Pseudomonas strains | Cold deserts of the Himalayas | Gluconic acid, oxalic acid, 2-ketogluconic acid, lactic acid, succinic acid, formic acid, citric acid and malic acid | [193] |
| Pseudomonas corrugata (NRRL B-30409) | Culture Collection | Gluconic and 2-ketogluconic acids | [194] |
| PPSB | Origin | Media Used | Plant Growth Enhancers | Reference |
|---|---|---|---|---|
| Bacillus weihenstephanensis MF593886 | Gentiana kurroo Royle rhizosphere | PVK | Siderophore, HCN, ammonia, and proteases | [207] |
| Pseudomonas, Serratia, and Flavobacterium | Rhizosphere and phyllosphere of Andes Mountains and Patagonia of Chile | PVK | IAA, ACC deaminase, anti-phytopathogenic activities | [12] |
| Pseudomonas | Snow sample | NBRIP | Siderophores, cellulases, xylanases, and chitinases | [2] |
| Pseudomonas koreensis P2 | Sela Lake | NBRIP | IAA, siderophore, HCN, and iron uptake | [208] |
| Acinetobacter, Bacillus, Enterobacter, Klebsiella, Proteus, Pseudomonas, and Staphylococcus | Renuka Lake | PVK | Ammonia, HCN, Zn solubilization, and hydrolytic enzymes | [202] |
| Pseudomonas | Antarctic soils | NBRIP | IAA, siderophores, HCN, microbial volatile organic compounds (MVOCs) | [209] |
| Pseudomonas simiae PS2 | Seabuckthorn (Hippophae rhamnoides L.) rhizospheric soil of high altitude in trans-Himalaya | PVK | IAA, siderophore and HCN | [40] |
| Pseudomonas, Bacillus, Paenibacillus, Sporosarcina, Cupriavidus and Paenarthrobacter | Lepidium meyenii Walp. Rhizosphere | NBRIP | IAA | [210] |
| Bacillus licheniformis, Bacillus muralis, Desemzia incerta, Paenibacillus tylopili and Sporosarcina globispora | Soil and water samples | PVK | IAA, GA, siderophores, NH3, HCN, ACC deaminase | [211] |
| Pseudomonas koreensis and Arthrobacter nitroguajacolicus strainYB4 | Rainfed agriculture field | PVK | IAA | [212] |
| Pseudomonas | Rhizospheric soil | NBRIP | IAA, siderophore, ACC deaminase, ammonia, NF, and antifungal compounds | [213] |
| Bacillus | HCN, ammonia, and NF | |||
| Stenotrophomonas | IAA, GA, HCN, Siderophore, ACC deaminase, ammonia, NF, and antifungal compounds | |||
| Arthrobacter | IAA, siderophore, ACC deaminase, ammonia, and NF | |||
| Acinetobacter | IAA, GA, HCN, siderophore, ACC deaminase, ammonia, NF, and antifungal compounds | |||
| Exiguobacterium | Siderophores, ammonia, and antifungal compounds | |||
| Providencia | IAA, GA, HCN, siderophore, ACC deaminase, ammonia, and NF | |||
| Flavobacterium | IAA, HCN, siderophore, ACC deaminase, and ammonia | |||
| Kocuria | IAA, GA, HCN, and siderophore | |||
| Pseudomonas | Glacial ice | NBRIP | IAA, HCN, siderophore, proteases amylases and galactosidases | [18] |
| Rhizobia | Pea nodules | PVK | IAA | [22] |
| Azotobacter, Pseudomonas, Micrococcus, and Bacillus | Pennisetum clandestinum rhizospheres | PVK | IAA and siderophores | [214] |
| Pseudomonas spp. | High altitude of the northwest Indian Himalayas | NBRIP | IAA, siderophore | [27] |
| Pseudomonas sp. | Deschampsia antarctica rhizosphere | PVK | IAA and EPS | [215] |
| Exiguobacterium acetylicum strain 1P | High altitude soil | NBRIP | IAA, siderophore and HCN | [216] |
| Pseudomonas fragi CS11RH1 | High altitude garlic rhizosphere | NBRIP | IAA and HCN | [14] |
| Fluorescent Pseudomonas | Garhwal Himalayas region | PVK | Siderophores, antifungal activity | [15] |
| Inoculated Crops | Cold Active PSB | Conditions | Agronomical Traits | References |
|---|---|---|---|---|
| Arabidopsis thaliana | Pseudomonas sp., Pseudomonas proteolytica, Pseudomonas azotoformans | Growth Chamber | Promoted overall growth such as rosette diameter, leaf area, and biomass | [20] |
| Tomato | Mixture of Pseudomonas sp. TmR5a and Curtobacterium sp. BmP22c (BC3) | Pot assay | Promoted the germination by 90% and significantly increased the root lengths | [12] |
| Tomato | Pseudomonas | Greenhouse | Increased germination and plantlets | [2] |
| Wheat | Pseudomonas | Paper Roll Towel Bioassay | Significantly increased root and shoot-lengths | [209] |
| Tomato | Pseudomonas simiae | Pot experiments in green shade net and open field conditions | Enhanced plant growth, increased fruit yield by 9.8% (net house) 19.8% (open field) | [40] |
| Chickpea, green gram, pea, and maize | Lysinibaccilus macroides ST-30, P. palleroniana N-26 and P. jessenii MP-1 | In vitro seed germination assay | Significantly increased the germination efficiency | [253] |
| Red clover | Pseudomonas, Bacillus, Paenibacillus, Sporosarcina, Cupriavidus and Paenarthrobacter | Water agar plates | Increased seed germination | [210] |
| Rice | Pseudomonas koreensis and Arthrobacter nitroguajacolicus strainYB4 | Greenhouse | Efficiently increased the biomass and P uptake | [212] |
| Lentil | Pseudomonas spp. | Temperature controlled polyhouse | Significantly increased the plant growth, grain yield, and P uptake | [27] |
| Barley, chickpea, pea, and maize | Rahnella sp. | Greenhouse | Significantly increased growth of all crops, microplot testing of the PPSB inoculum also significantly increased growth and yield of pea | [44] |
| Wheat | Pseudomonas fragi CS11RH1 (MTCC 8984) | Greenhouse | Increased the percent germination, rate of germination, biomass, and nutrient uptake | [14] |
| Pseudomonas vancouverensis | Greenhouse | Increased germination and root and shoot length | [254] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizvi, A.; Ahmed, B.; Khan, M.S.; Umar, S.; Lee, J. Psychrophilic Bacterial Phosphate-Biofertilizers: A Novel Extremophile for Sustainable Crop Production under Cold Environment. Microorganisms 2021, 9, 2451. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122451
Rizvi A, Ahmed B, Khan MS, Umar S, Lee J. Psychrophilic Bacterial Phosphate-Biofertilizers: A Novel Extremophile for Sustainable Crop Production under Cold Environment. Microorganisms. 2021; 9(12):2451. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122451
Chicago/Turabian StyleRizvi, Asfa, Bilal Ahmed, Mohammad Saghir Khan, Shahid Umar, and Jintae Lee. 2021. "Psychrophilic Bacterial Phosphate-Biofertilizers: A Novel Extremophile for Sustainable Crop Production under Cold Environment" Microorganisms 9, no. 12: 2451. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122451