
Establishment Genes Present on pLS20 Family of Conjugative Plasmids Are Regulated in Two Different Ways

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Oligonucleotides
2.2. Construction of Plasmids and Strains
2.3. Flow Cytometry
2.4. In Silico Analyses
3. Results
3.1. pLS20 Contains an Anti-Restriction Gene Similar to the One on Plasmid p576, but Both Genes Are Preceded by Very Different Sequences
3.2. Sequences Highly Similar to Those Upstream of ardCpLS20 Are also Present Upstream of Four Operons on pLS20: Identification of the Establishment Regulon of pLS20
3.3. Features of the EGeRS1 Sequences
3.4. Functional Analysis of pLS20 EGeRS1-B
- EGeRS1-B does not contain a constitutive promoter
- EGeRS1-B sequence prevents readthrough of upstream promoters
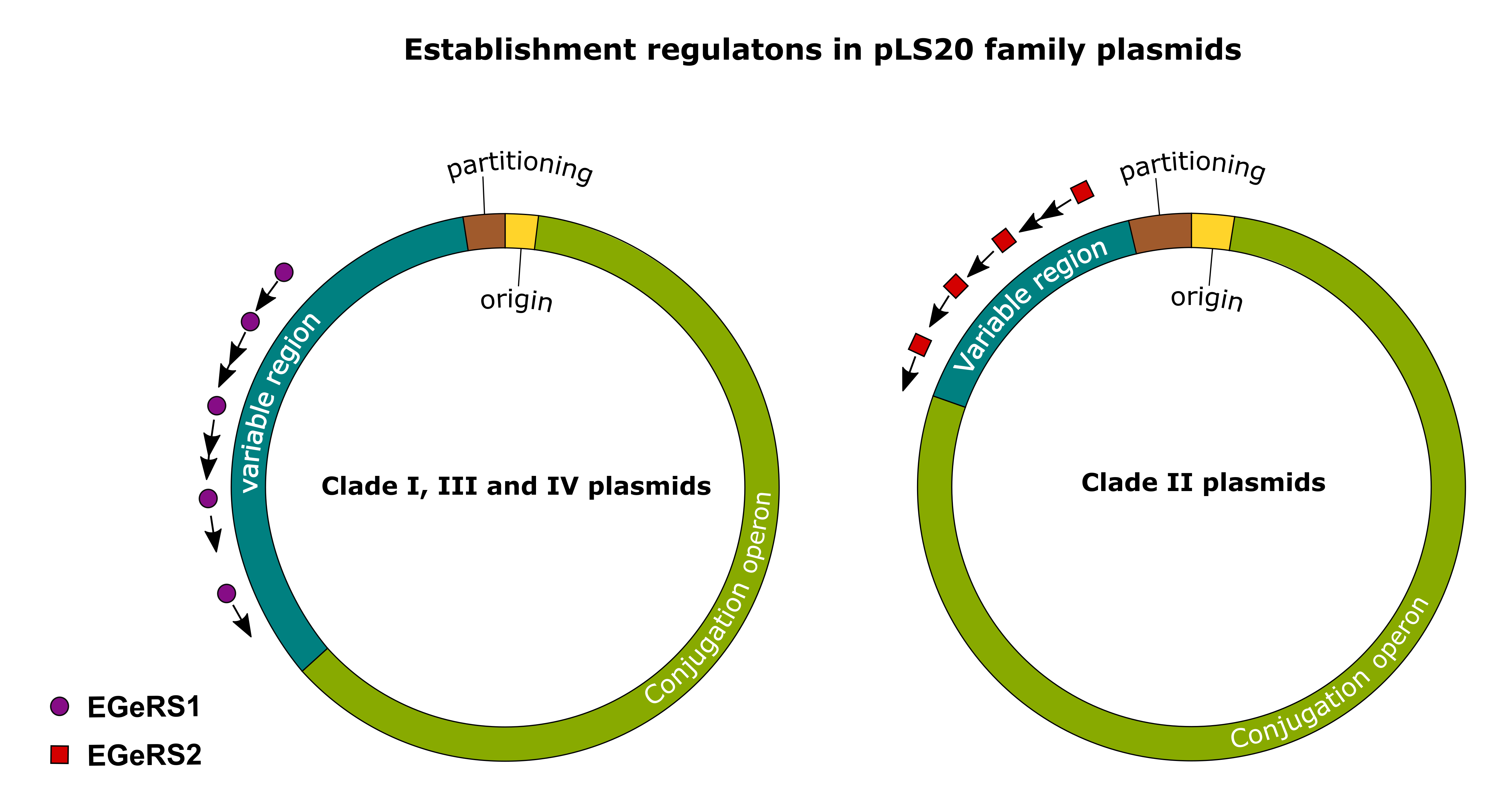
3.5. Establishment Genes Present on pLS20 Family Plasmids Are Regulated by One of the Two Different Mechanisms
3.6. Analysis of the EGeRS1 Sequences
3.7. Analysis of the EGeRS2 Sequences
3.8. (Putative) Establishment Genes Regulated by EGeRS2
3.9. (Putative) Establishment Genes Present on pLS20 Family Plasmids Regulated by EGeRS1
4. Discussion
4.1. Establishment Gene Regulatory Sequences Present on pLS20 Family of Plasmids
4.2. Establishment Regulons Present on pLS20 Family of Plasmids and Implications of Establishment Regulons in General
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Tock, M.R.; Dryden, D.T. The biology of restriction and anti-restriction. Curr. Opin. Microbiol. 2005, 8, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.H.; Touchon, M.; Rocha, E.P. The interplay of restriction-modification systems with mobile genetic elements and their prokaryotic hosts. Nucleic Acids Res. 2014, 42, 10618–10631. [Google Scholar] [CrossRef] [PubMed]
- Mojica, F.J.; Rodriguez-Valera, F. The discovery of CRISPR in archaea and bacteria. FEBS J. 2016, 283, 3162–3169. [Google Scholar] [CrossRef] [PubMed]
- Bernheim, A.; Sorek, R. The pan-immune system of bacteria: Antiviral defence as a community resource. Nat. Rev. Microbiol. 2020, 18, 113–119. [Google Scholar] [CrossRef]
- Chilley, P.M.; Wilkins, B.M. Distribution of the ardA family of antirestriction genes on conjugative plasmids. Microbiology 1995, 141 Pt. 9, 2157–2164. [Google Scholar] [CrossRef] [Green Version]
- Belogurov, A.A.; Delver, E.P.; Rodzevich, O.V. Plasmid pKM101 encodes two nonhomologous antirestriction proteins (ArdA and ArdB) whose expression is controlled by homologous regulatory sequences. J. Bacteriol. 1993, 175, 4843–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belogurov, A.A.; Delver, E.P.; Agafonova, O.V.; Belogurova, N.G.; Lee, L.Y.; Kado, C.I. Antirestriction protein Ard (Type C) encoded by IncW plasmid pSa has a high similarity to the "protein transport" domain of TraC1 primase of promiscuous plasmid RP4. J. Mol. Biol. 2000, 296, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, B.M. Plasmid promiscuity: Meeting the challenge of DNA immigration control. Environ. Microbiol. 2002, 4, 495–500. [Google Scholar] [CrossRef]
- Val-Calvo, J.; Luque-Ortega, J.R.; Crespo, I.; Miguel-Arribas, A.; Abia, D.; Sanchez-Hevia, D.L.; Serrano, E.; Gago-Cordoba, C.; Ares, S.; Alfonso, C.; et al. Novel regulatory mechanism of establishment genes of conjugative plasmids. Nucleic Acids Res. 2018, 46, 11910–11926. [Google Scholar] [CrossRef] [PubMed]
- Val-Calvo, J.; Miguel-Arribas, A.; Abia, D.; Wu, L.J.; Meijer, W.J.J. pLS20 is the archetype of a new family of conjugative plasmids harboured by Bacillus species. NAR Genom. Bioinform. 2021, 3, lqab096. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Bron, S.; Meijer, W.J.J.; Holsappel, S.; Haima, P. Plasmid instability and molecular cloning in Bacillus subtilis. Res. Microbiol. 1991, 142, 875–883. [Google Scholar] [CrossRef]
- Miguel-Arribas, A.; Val-Calvo, J.; Gago-Cordoba, C.; Izquierdo, J.M.; Abia, D.; Wu, L.J.; Errington, J.; Meijer, W.J.J. A novel bipartite antitermination system widespread in conjugative elements of Gram-positive bacteria. Nucleic Acids Res. 2021. [Google Scholar] [CrossRef]
- Gago-Cordoba, C.; Val-Calvo, J.; Miguel-Arribas, A.; Serrano, E.; Singh, P.K.; Abia, D.; Wu, L.J.; Meijer, W.J.J. Surface Exclusion Revisited: Function Related to Differential Expression of the Surface Exclusion System of Bacillus subtilis Plasmid pLS20. Front. Microbiol. 2019, 10, 1502. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Honer Zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- Hofacker, I.L. RNA consensus structure prediction with RNAalifold. Methods Mol. Biol. 2007, 395, 527–544. [Google Scholar] [CrossRef]
- Erb, I.; Gonzalez-Vallinas, J.R.; Bussotti, G.; Blanco, E.; Eyras, E.; Notredame, C. Use of ChIP-Seq data for the design of a multiple promoter-alignment method. Nucleic Acids Res. 2012, 40, e52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, D.; Proctor, J.R.; Zhu, J.Y.; Meyer, I.M. R-CHIE: A web server and R package for visualizing RNA secondary structures. Nucleic Acids Res. 2012, 40, e95. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.K.; Ballestero-Beltran, S.; Ramachandran, G.; Meijer, W.J. Complete nucleotide sequence and determination of the replication region of the sporulation inhibiting plasmid p576 from Bacillus pumilus NRS576. Res. Microbiol. 2010, 161, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Meijer, W.J.J.; de Boer, A.; van Tongeren, S.; Venema, G.; Bron, S. Characterization of the replication region of the Bacillus subtilis plasmid pLS20: A novel type of replicon. Nucleic Acids Res. 1995, 23, 3214–3223. [Google Scholar] [CrossRef]
- Titok, M.A.; Chapuis, J.; Selezneva, Y.V.; Lagodich, A.V.; Prokulevich, V.A.; Ehrlich, S.D.; Jannière, L. Bacillus subtilis soil isolates: Plasmid replicon analysis and construction of a new theta-replicating vector. Plasmid 2003, 49, 53–62. [Google Scholar] [CrossRef]
- Titok, M.; Suski, C.; Dalmais, B.; Ehrlich, S.D.; Janniere, L. The replicative polymerases PolC and DnaE are required for theta replication of the Bacillus subtilis plasmid pBS72. Microbiology 2006, 152, 1471–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, S.Y.; Borges, A.L.; Chen, K.H.; Swaney, D.L.; Krogan, N.J.; Bondy-Denomy, J.; Davidson, A.R. Anti-CRISPR-Associated Proteins Are Crucial Repressors of Anti-CRISPR Transcription. Cell 2019, 178, 1452–1464 e1413. [Google Scholar] [CrossRef]
- Hamon, M.A.; Stanley, N.R.; Britton, R.A.; Grossman, A.D.; Lazazzera, B.A. Identification of AbrB-regulated genes involved in biofilm formation by Bacillus subtilis. Mol. Microbiol. 2004, 52, 847–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osuna, B.A.; Karambelkar, S.; Mahendra, C.; Sarbach, A.; Johnson, M.C.; Kilcher, S.; Bondy-Denomy, J. Critical Anti-CRISPR Locus Repression by a Bi-functional Cas9 Inhibitor. Cell Host Microbe 2020, 28, 23–30 e25. [Google Scholar] [CrossRef]
- Bates, S.; Roscoe, R.A.; Althorpe, N.J.; Brammar, W.J.; Wilkins, B.M. Expression of leading region genes on IncI1 plasmid ColIb-P9: Genetic evidence for single-stranded DNA transcription. Microbiology 1999, 145 Pt. 10, 2655–2662. [Google Scholar] [CrossRef] [Green Version]
- Masai, H.; Arai, K. Frpo: A novel single-stranded DNA promoter for transcription and for primer RNA synthesis of DNA replication. Cell 1997, 89, 897–907. [Google Scholar] [CrossRef] [Green Version]
- Nasim, M.T.; Eperon, I.C.; Wilkins, B.M.; Brammar, W.J. The activity of a single-stranded promoter of plasmid ColIb-P9 depends on its secondary structure. Mol. Microbiol. 2004, 53, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Althorpe, N.J.; Chilley, P.M.; Thomas, A.T.; Brammar, W.J.; Wilkins, B.M. Transient transcriptional activation of the Incl1 plasmid anti-restriction gene (ardA) and SOS inhibition gene (psiB) early in conjugating recipient bacteria. Mol. Microbiol. 1999, 31, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.; Miguel-Arribas, A.; Abia, D.; Singh, P.K.; Crespo, I.; Gago-Cordoba, C.; Hao, J.A.; Luque-Ortega, J.R.; Alfonso, C.; Wu, L.J.; et al. Discovery of a new family of relaxases in Firmicutes bacteria. PLoS Genet. 2017, 13, e1006586. [Google Scholar] [CrossRef]
- Goryanin, I.I.; Kudryavtseva, A.A.; Balabanov, V.P.; Biryukova, V.S.; Manukhov, I.V.; Zavilgelsky, G.B. Antirestriction activities of KlcA (RP4) and ArdB (R64) proteins. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Gonzalez-Montes, L.; Del Campo, I.; Garcillan-Barcia, M.P.; de la Cruz, F.; Moncalian, G. ArdC, a ssDNA-binding protein with a metalloprotease domain, overpasses the recipient hsdRMS restriction system broadening conjugation host range. PLoS Genet. 2020, 16, e1008750. [Google Scholar] [CrossRef] [PubMed]
- Balabanov, V.P.; Kotova, V.Y.; Kholodii, G.Y.; Mindlin, S.Z.; Zavilgelsky, G.B. A novel gene, ardD, determines antirestriction activity of the non-conjugative transposon Tn5053 and is located antisense within the tniA gene. FEMS Microbiol. Lett. 2012, 337, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, S.A.; Roberts, G.A.; Johnson, K.A.; Cooper, L.P.; Liu, H.; White, J.H.; Carter, L.G.; Sanghvi, B.; Oke, M.; Walkinshaw, M.D.; et al. Extensive DNA mimicry by the ArdA anti-restriction protein and its role in the spread of antibiotic resistance. Nucleic Acids Res. 2009, 37, 4887–4897. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Snir, S.; Koonin, E.V. Defense islands in bacterial and archaeal genomes and prediction of novel defense systems. J. Bacteriol. 2011, 193, 6039–6056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinilla-Redondo, R.; Shehreen, S.; Marino, N.D.; Fagerlund, R.D.; Brown, C.M.; Sorensen, S.J.; Fineran, P.C.; Bondy-Denomy, J. Discovery of multiple anti-CRISPRs highlights anti-defense gene clustering in mobile genetic elements. Nat. Commun. 2020, 11, 5652. [Google Scholar] [CrossRef] [PubMed]
- Mahendra, C.; Christie, K.A.; Osuna, B.A.; Pinilla-Redondo, R.; Kleinstiver, B.P.; Bondy-Denomy, J. Broad-spectrum anti-CRISPR proteins facilitate horizontal gene transfer. Nat. Microbiol. 2020, 5, 620–629. [Google Scholar] [CrossRef]
- Roy, D.; Huguet, K.T.; Grenier, F.; Burrus, V. IncC conjugative plasmids and SXT/R391 elements repair double-strand breaks caused by CRISPR-Cas during conjugation. Nucleic Acids Res. 2020, 48, 8815–8827. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Comparative genomics of defense systems in archaea and bacteria. Nucleic Acids Res. 2013, 41, 4360–4377. [Google Scholar] [CrossRef] [PubMed]
- van Houte, S.; Buckling, A.; Westra, E.R. Evolutionary Ecology of Prokaryotic Immune Mechanisms. Microbiol. Mol. Biol. Rev. 2016, 80, 745–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Makarova, K.S.; Wolf, Y.I. Evolutionary Genomics of Defense Systems in Archaea and Bacteria. Annu. Rev. Microbiol. 2017, 71, 233–261. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.H.; Touchon, M.; Cury, J.; Rocha, E.P.C. The chromosomal organization of horizontal gene transfer in bacteria. Nat. Commun. 2017, 8, 841. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Val-Calvo, J.; Miguel-Arribas, A.; Freire, F.; Abia, D.; Wu, L.J.; Meijer, W.J.J. Establishment Genes Present on pLS20 Family of Conjugative Plasmids Are Regulated in Two Different Ways. Microorganisms 2021, 9, 2465. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122465
Val-Calvo J, Miguel-Arribas A, Freire F, Abia D, Wu LJ, Meijer WJJ. Establishment Genes Present on pLS20 Family of Conjugative Plasmids Are Regulated in Two Different Ways. Microorganisms. 2021; 9(12):2465. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122465
Chicago/Turabian StyleVal-Calvo, Jorge, Andrés Miguel-Arribas, Fernando Freire, David Abia, Ling Juan Wu, and Wilfried J.J. Meijer. 2021. "Establishment Genes Present on pLS20 Family of Conjugative Plasmids Are Regulated in Two Different Ways" Microorganisms 9, no. 12: 2465. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122465