
Investigation of Transovarial Transmission of Bartonella henselae in Rhipicephalus sanguineus sensu lato Ticks Using Artificial Feeding


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
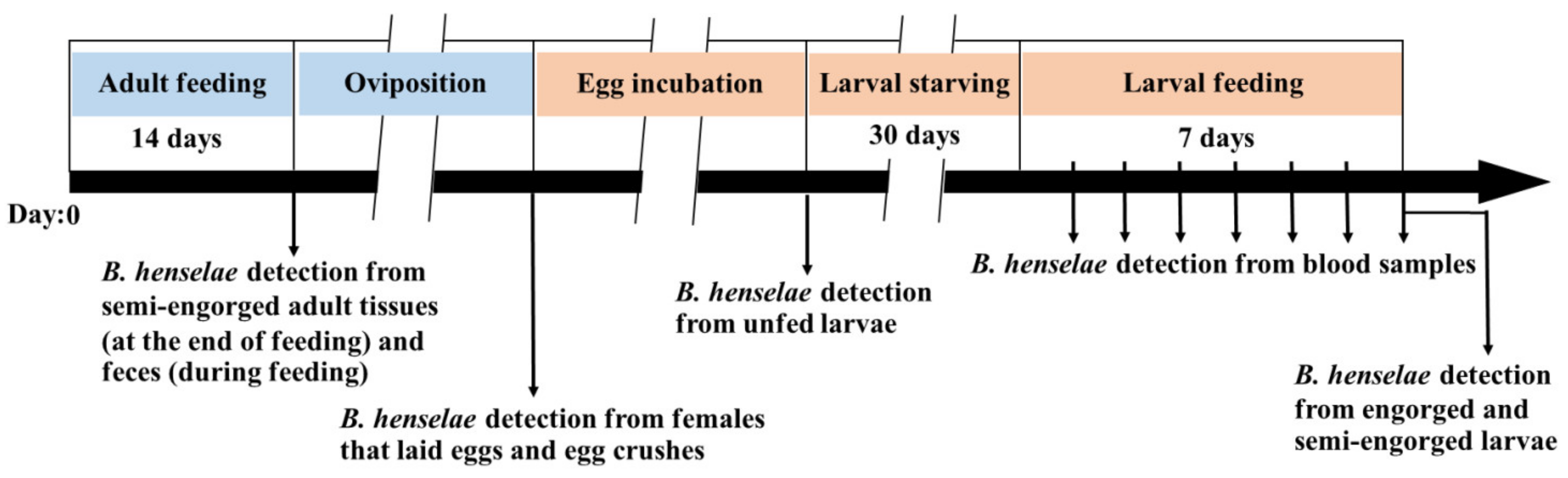
2.1. Study Design
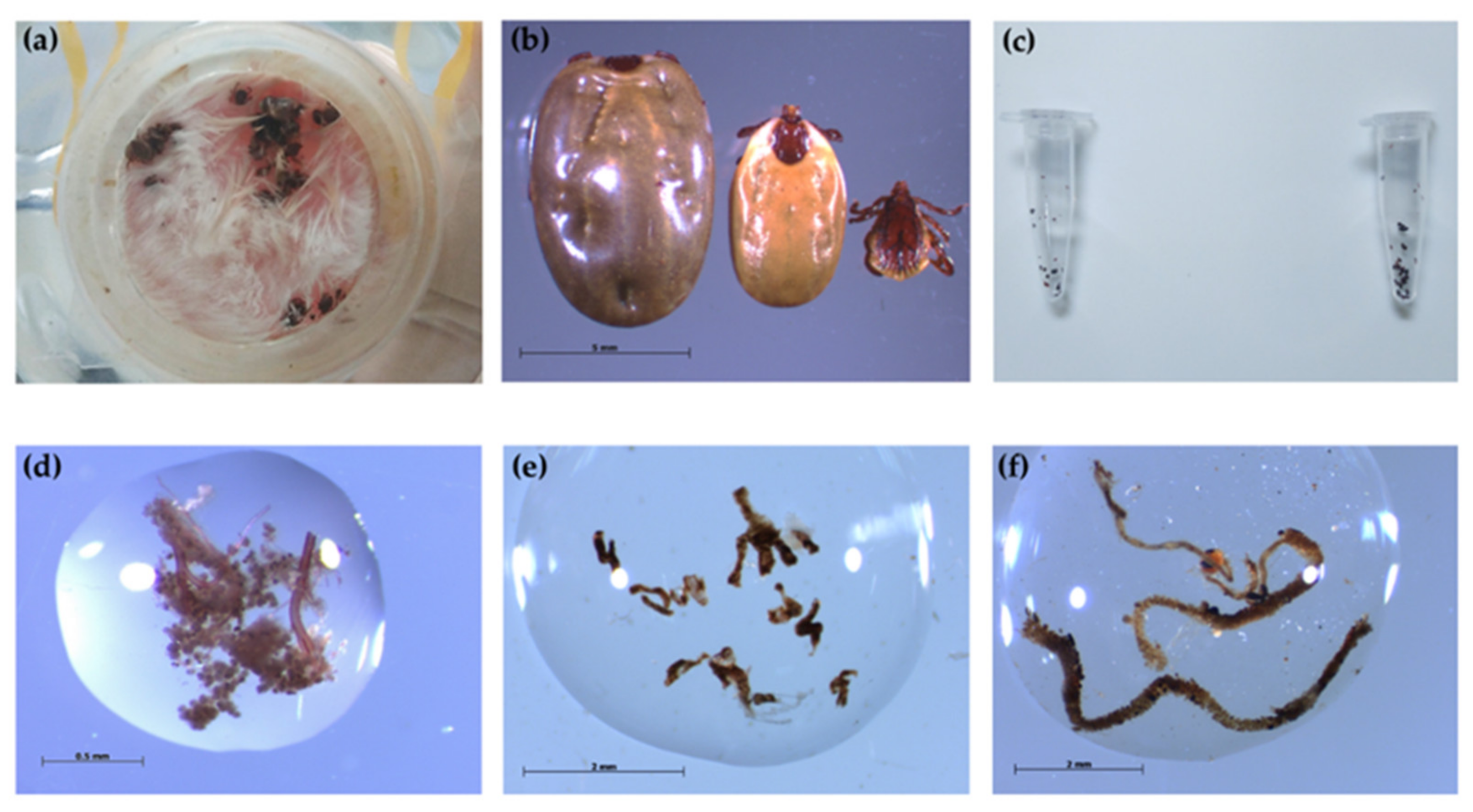
2.2. Rhipicephalus sanguineus s.l. Ticks
2.3. Bartonella henselae Isolates
2.4. Artificial Membrane Feeding System
2.5. B. henselae Infection in R. sanguineus s.l. Adult Ticks
2.6. Feeding Larval Ticks with Non-Infected Blood
2.7. DNA Extraction from Tick Samples, Tick Feces, Blood Samples, and Bacterial Colonies
2.8. Nested PCR Assay
2.9. Culturing of B. henselae from Blood during Larval Feeding and Tick Fecal Samples
2.10. Genetic Characterization of B. henselae
2.11. Identification of Bacterial Colonies Other Than B. henselae by Mass Spectrometry
2.12. Statistical Analysis
3. Results
3.1. Rhipicephalus sanguineus s.l. Ticks Engorged by Artificial Membrane Feeding System
3.2. Bartonella henselae Acquisition by Adult Ticks during Feeding on Infected Blood Meal
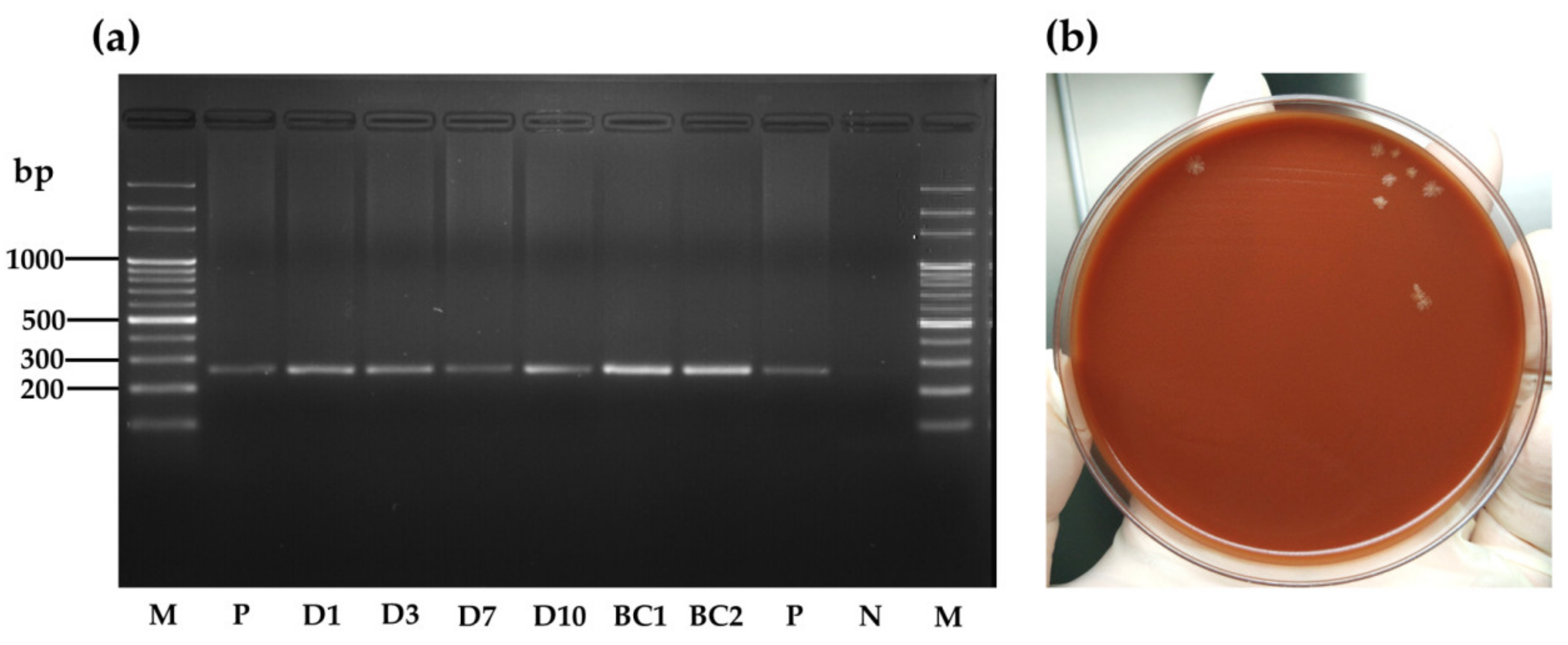
3.2.1. Bartonella henselae Detection in R. sanguineus s.l. Adult Samples
3.2.2. Persistence of B. henselae in R. sanguineus s.l. Feces
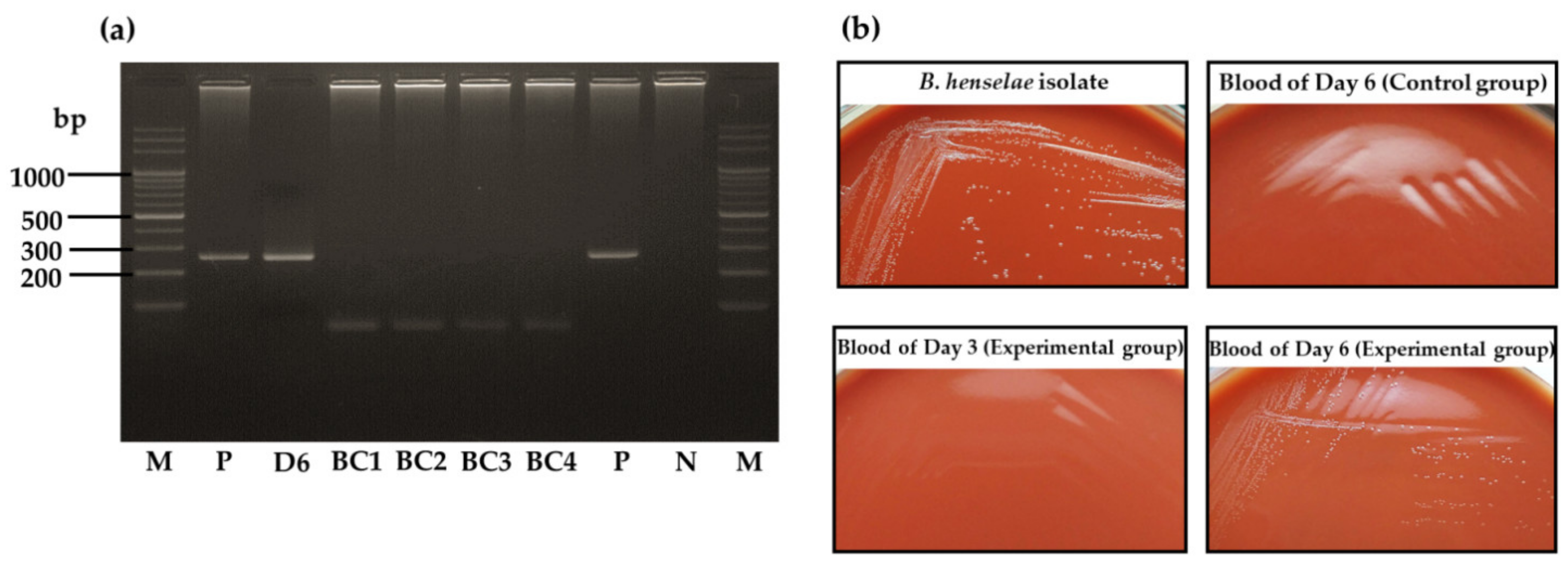
3.3. Absence of B. henselae DNA in Pooled Eggs and Unfed Larvae Derived from B. henselae-Infected Female Ticks
3.4. Bartonella henselae Transmission by Larvae Infected at the Adult Stage
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chomel, B.B.; Kasten, R.W. Bartonellosis, an increasingly recognized zoonosis. J. Appl. Microbiol. 2010, 109, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.C.; Chomel, B.B.; Kasten, R.W.; Floyd-Hawkins, K.A.; Kikuchi, Y.; Koehler, J.E.; Pedersen, N.C. Experimental and natural infection with Bartonella henselae in domestic cats. Comp. Immunol. Microbiol. Infect. Dis. 1997, 20, 41–51. [Google Scholar] [CrossRef]
- Chomel, B.B.; Kasten, R.W.; Floyd-Hawkins, K.; Chi, B.; Yamamoto, K.; Roberts-Wilson, J.; Gurfield, A.N.; Abbott, R.C.; Pedersen, N.C.; Koehler, J.E. Experimental transmission of Bartonella henselae by the cat flea. J. Clin. Microbiol. 1996, 34, 1952–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foil, L.; Andress, E.; Freeland, R.L.; Roy, A.F.; Rutledge, R.; Triche, P.C.; O’Reilly, K.L. Experimental infection of domestic cats with Bartonella henselae by inoculation of Ctenocephalides felis (Siphonaptera: Pulicidae) feces. J. Med. Entomol. 1998, 35, 625–628. [Google Scholar] [CrossRef]
- Higgins, J.A.; Radulovic, S.; Jaworski, D.C.; Azad, A.F. Acquisition of the cat scratch disease agent Bartonella henselae by cat fleas (Siphonaptera: Pulicidae). J. Med. Entomol. 1996, 33, 490–495. [Google Scholar] [CrossRef]
- Iannino, F.; Salucci, S.; Di Provvido, A.; Paolini, A.; Ruggieri, E. Bartonella infections in humans dogs and cats. Vet. Ital. 2018, 54, 63–72. [Google Scholar] [CrossRef]
- Carithers, H.A.; Carithers, C.M.; Edwards, R.O., Jr. Cat scratch disease: The larger view. Pediatrics 1969, 43, 629–631. [Google Scholar]
- Arisoy, E.S.; Correa, A.G.; Wagner, M.L.; Kaplan, S.L. Hepatosplenic cat-scratch disease in children: Selected clinical features and treatment. Clin. Infect. Dis. 1999, 28, 778–784. [Google Scholar] [CrossRef] [Green Version]
- Chian, C.A.; Arrese, J.E.; Pierard, G.E. Skin manifestations of Bartonella infections. Int. J. Dermatol. 2002, 41, 461–466. [Google Scholar] [CrossRef]
- Chang, C.C.; Chomel, B.B.; Kasten, R.W.; Romano, V.; Tietze, N. Molecular evidence of Bartonella spp. in questing adult Ixodes pacificus ticks in California. J. Clin. Microbiol. 2001, 39, 1221–1226. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.Y.; Kasten, R.W.; Paff, S.M.; Van Horn, B.A.; Vayssier-Taussat, M.; Boulouis, H.J.; Chomel, B.B. Bartonella spp. DNA associated with biting flies from California. Emerg. Infect. Dis. 2004, 10, 1311–1313. [Google Scholar] [CrossRef]
- Durden, L.A.; Ellis, B.A.; Banks, C.W.; Crowe, J.D.; Oliver, J.H., Jr. Ectoparasites of gray squirrels in two different habitats and screening of selected ectoparasites for bartonellae. J. Parasitol. 2004, 90, 485–489. [Google Scholar] [CrossRef]
- Rar, V.A.; Fomenko, N.V.; Dobrotvorsky, A.K.; Livanova, N.N.; Rudakova, S.A.; Fedorov, E.G.; Astanin, V.B.; Morozova, O.V. Tickborne pathogen detection, Western Siberia, Russia. Emerg. Infect. Dis. 2005, 11, 1708–1715. [Google Scholar] [CrossRef]
- Reeves, W.K.; Nelder, M.P.; Cobb, K.D.; Dasch, G.A. Bartonella spp. in deer keds, Lipoptena mazamae (Diptera: Hippoboscidae), from Georgia and South Carolina, USA. J. Wildl. Dis. 2006, 42, 391–396. [Google Scholar] [CrossRef]
- Reeves, W.K.; Szumlas, D.E.; Moriarity, J.R.; Loftis, A.D.; Abbassy, M.M.; Helmy, I.M.; Dasch, G.A. Louse-borne bacterial pathogens in lice (Phthiraptera) of rodents and cattle from Egypt. J. Parasitol. 2006, 92, 313–318. [Google Scholar] [CrossRef]
- Regier, Y.; Ballhorn, W.; Kempf, V.A. Molecular detection of Bartonella henselae in 11 Ixodes ricinus ticks extracted from a single cat. Parasit. Vectors 2017, 10, 105. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.L.; Lin, C.C.; Chomel, B.B.; Chuang, S.T.; Tsai, K.H.; Wu, W.J.; Huang, C.G.; Yu, J.C.; Sung, M.H.; Kass, P.H.; et al. Bartonella infection in shelter cats and dogs and their ectoparasites. Vector Borne Zoonotic Dis. 2011, 11, 1023–1030. [Google Scholar] [CrossRef]
- Lucey, D.; Dolan, M.J.; Moss, C.W.; Garcia, M.; Hollis, D.G.; Wegner, S.; Morgan, G.; Almeida, R.; Leong, D.; Greisen, K.S.; et al. Relapsing illness due to Rochalimaea henselae in immunocompetent hosts: Implication for therapy and new epidemiological associations. Clin. Infect. Dis. 1992, 14, 683–688. [Google Scholar] [CrossRef]
- Cotte, V.; Bonnet, S.; Le Rhun, D.; Le Naour, E.; Chauvin, A.; Boulouis, H.J.; Lecuelle, B.; Lilin, T.; Vayssier-Taussat, M. Transmission of Bartonella henselae by Ixodes ricinus. Emerg. Infect. Dis. 2008, 14, 1074–1080. [Google Scholar] [CrossRef]
- Reis, C.; Cote, M.; Le Rhun, D.; Lecuelle, B.; Levin, M.L.; Vayssier-Taussat, M.; Bonnet, S.I. Vector competence of the tick Ixodes ricinus for transmission of Bartonella birtlesii. PLoS Negl. Trop. Dis. 2011, 5, e1186. [Google Scholar] [CrossRef] [Green Version]
- Krol, N.; Militzer, N.; Stobe, E.; Nijhof, A.M.; Pfeffer, M.; Kempf, V.A.J.; Obiegala, A. Evaluating Transmission Paths for Three Different Bartonella spp. in Ixodes ricinus Ticks Using Artificial Feeding. Microorganisms 2021, 9, 901. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasites Vectors 2010, 3, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Satta, G.; Chisu, V.; Cabras, P.; Fois, F.; Masala, G. Pathogens and symbionts in ticks: A survey on tick species distribution and presence of tick-transmitted micro-organisms in Sardinia, Italy. J. Med. Microbiol. 2011, 60, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Wikswo, M.E.; Hu, R.; Metzger, M.E.; Eremeeva, M.E. Detection of Rickettsia rickettsii and Bartonella henselae in Rhipicephalus sanguineus ticks from California. J. Med. Entomol. 2007, 44, 158–162. [Google Scholar] [CrossRef]
- Wechtaisong, W.; Bonnet, S.I.; Lien, Y.Y.; Chuang, S.T.; Tsai, Y.L. Transmission of Bartonella henselae within Rhipicephalus sanguineus: Data on the Potential Vector Role of the Tick. PLoS Negl. Trop. Dis. 2020, 14, e0008664. [Google Scholar] [CrossRef]
- Troughton, D.R.; Levin, M.L. Life cycles of seven ixodid tick species (Acari: Ixodidae) under standardized laboratory conditions. J. Med. Entomol. 2007, 44, 732–740. [Google Scholar] [CrossRef]
- Bonnet, S.; Jouglin, M.; Malandrin, L.; Becker, C.; Agoulon, A.; L’Hostis, M.; Chauvin, A. Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology 2007, 134, 197–207. [Google Scholar] [CrossRef]
- Lejal, E.; Moutailler, S.; Simo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasites Vectors 2019, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Rolain, J.M.; Franc, M.; Davoust, B.; Raoult, D. Molecular detection of Bartonella quintana, B. koehlerae, B. henselae, B. clarridgeiae, Rickettsia felis, and Wolbachia pipientis in cat fleas, France. Emerg. Infect. Dis. 2003, 9, 338–342. [Google Scholar] [CrossRef]
- Sato, S.; Kabeya, H.; Negishi, A.; Tsujimoto, H.; Nishigaki, K.; Endo, Y.; Maruyama, S. Molecular survey of Bartonella henselae and Bartonella clarridgeiae in pet cats across Japan by species-specific nested-PCR. Epidemiol. Infect. 2017, 145, 2694–2700. [Google Scholar] [CrossRef] [Green Version]
- Krober, T.; Guerin, P.M. In vitro feeding assays for hard ticks. Trends Parasitol. 2007, 23, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.C.J.; Edwards, G.A.; Martin, D.J.; Huang, H.; Crichton, M.L.; Kendall, M.A.F. Allometric scaling of skin thickness, elasticity, viscoelasticity to mass for micro-medical device translation: From mice, rats, rabbits, pigs to humans. Sci. Rep. 2017, 7, 15885. [Google Scholar] [CrossRef]
- Sojka, D.; Franta, Z.; Horn, M.; Caffrey, C.R.; Mares, M.; Kopacek, P. New insights into the machinery of blood digestion by ticks. Trends Parasitol. 2013, 29, 276–285. [Google Scholar] [CrossRef]
- Dai, J.; Narasimhan, S.; Zhang, L.; Liu, L.; Wang, P.; Fikrig, E. Tick histamine release factor is critical for Ixodes scapularis engorgement and transmission of the lyme disease agent. PLoS Pathog. 2010, 6, e1001205. [Google Scholar] [CrossRef]
- De la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Pena, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-Pathogen Interactions and Vector Competence: Identification of Molecular Drivers for Tick-Borne Diseases. Front. Cell. Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente, J.; Villar, M.; Cabezas-Cruz, A.; Estrada-Pena, A.; Ayllon, N.; Alberdi, P. Tick-Host-Pathogen Interactions: Conflict and Cooperation. PLoS Pathog. 2016, 12, e1005488. [Google Scholar] [CrossRef]
- Hu, R.; Hyland, K.E.; Markowski, D. Effects of Babesia microti infection on feeding pattern, engorged body weight, and molting rate of immature Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1997, 34, 559–564. [Google Scholar] [CrossRef]
- Lefcort, H.; Durden, L.A. The effect of infection with Lyme disease spirochetes (Borrelia burgdorferi) on the phototaxis, activity, and questing height of the tick vector Ixodes scapularis. Parasitology 1996, 113, 97–103. [Google Scholar] [CrossRef]
- Randolph, S.E. The effect of Babesia microti on feeding and survival in its tick vector, Ixodes trianguliceps. Parasitology 1991, 102, 9–16. [Google Scholar] [CrossRef]
- Romashchenko, A.V.; Ratushnyak, A.S.; Zapara, T.A.; Tkachev, S.E.; Moshkin, M.P. The correlation between tick (Ixodes persulcatus Sch.) questing behaviour and synganglion neuronal responses to odours. J. Insect Physiol. 2012, 58, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Cote, M.; Paul, R.E.; Bonnet, S.I. Impact of feeding system and infection status of the blood meal on Ixodes ricinus feeding. Ticks Tick-Borne Dis. 2014, 5, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.L.; Chang, C.C.; Chuang, S.T.; Chomel, B.B. Bartonella species and their ectoparasites: Selective host adaptation or strain selection between the vector and the mammalian host? Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Blisnick, A.A.; Foulon, T.; Bonnet, S.I. Serine Protease Inhibitors in Ticks: An Overview of Their Role in Tick Biology and Tick-Borne Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.K.; Tsuji, N.; Miyoshi, T.; Alim, M.A.; Huang, X.; Hatta, T.; Fujisaki, K. The Kunitz-like modulatory protein haemangin is vital for hard tick blood-feeding success. PLoS Pathog. 2009, 5, e1000497. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; de la Fuente, J.; Cote, M.; Galindo, R.C.; Moutailler, S.; Vayssier-Taussat, M.; Bonnet, S.I. IrSPI, a tick serine protease inhibitor involved in tick feeding and Bartonella henselae infection. PLoS Negl. Trop. Dis. 2014, 8, e2993. [Google Scholar] [CrossRef]
- Hajdusek, O.; Sima, R.; Ayllon, N.; Jalovecka, M.; Perner, J.; de la Fuente, J.; Kopacek, P. Interaction of the tick immune system with transmitted pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Billeter, S.A.; Diniz, P.P.; Battisti, J.M.; Munderloh, U.G.; Breitschwerdt, E.B.; Levy, M.G. Infection and replication of Bartonella species within a tick cell line. Exp. Appl. Acarol. 2009, 49, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Bouhsira, E.; Franc, M.; Boulouis, H.J.; Jacquiet, P.; Raymond-Letron, I.; Lienard, E. Assessment of persistence of Bartonella henselae in Ctenocephalides felis. Appl. Environ. Microbiol. 2013, 79, 7439–7444. [Google Scholar] [CrossRef] [Green Version]
- Billeter, S.A.; Kasten, R.W.; Killmaster, L.F.; Breitschwerdt, E.B.; Levin, M.L.; Levy, M.G.; Kosoy, M.Y.; Chomel, B.B. Experimental infection by capillary tube feeding of Rhipicephalus sanguineus with Bartonella vinsonii subspecies berkhoffii. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 9–15. [Google Scholar] [CrossRef]
- Chauvin, A.; Moreau, E.; Bonnet, S.; Plantard, O.; Malandrin, L. Babesia and its hosts: Adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet. Res. 2009, 40, 37. [Google Scholar] [CrossRef] [Green Version]
- Burgdorfer, W.; Brinton, L.P. Mechanisms of transovarial infection of spotted fever Rickettsiae in ticks. Ann. N. Y. Acad. Sci. 1975, 266, 61–72. [Google Scholar] [CrossRef]
- Rosell, R.; Coons, L.B. The role of the fat body, midgut and ovary in vitellogenin production and vitellogenesis in the female tick, Dermacentor variabilis. Int. J. Parasitol. 1992, 22, 341–349. [Google Scholar] [CrossRef]
- Andreotti, R.; Perez de Leon, A.A.; Dowd, S.E.; Guerrero, F.D.; Bendele, K.G.; Scoles, G.A. Assessment of bacterial diversity in the cattle tick Rhipicephalus (Boophilus) microplus through tag-encoded pyrosequencing. BMC Microbiol. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Perveen, N.; Muzaffar, S.B.; Vijayan, R.; Al-Deeb, M.A. Microbial communities associated with the camel tick, Hyalomma dromedarii: 16S rRNA gene-based analysis. Sci. Rep. 2020, 10, 17035. [Google Scholar] [CrossRef]
- Karim, S.; Budachetri, K.; Mukherjee, N.; Williams, J.; Kausar, A.; Hassan, M.J.; Adamson, S.; Dowd, S.E.; Apanskevich, D.; Arijo, A.; et al. A study of ticks and tick-borne livestock pathogens in Pakistan. PLoS Negl. Trop. Dis. 2017, 11, e0005681. [Google Scholar] [CrossRef]
- Loong, S.K.; Lim, F.S.; Khoo, J.J.; Lee, H.Y.; Suntharalingam, C.; Ishak, S.N.; Mohd-Taib, F.S.; AbuBakar, S. Culturable pathogenic bacteria in ticks parasitizing farm animals and rodents in Malaysia. Trop. Biomed. 2020, 37, 803–811. [Google Scholar] [CrossRef]
- Duron, O.; Noel, V.; McCoy, K.D.; Bonazzi, M.; Sidi-Boumedine, K.; Morel, O.; Vavre, F.; Zenner, L.; Jourdain, E.; Durand, P.; et al. The Recent Evolution of a Maternally-Inherited Endosymbiont of Ticks Led to the Emergence of the Q Fever Pathogen, Coxiella burnetii. PLoS Pathog. 2015, 11, e1004892. [Google Scholar] [CrossRef] [Green Version]
- Tsementzi, D.; Castro Gordillo, J.; Mahagna, M.; Gottlieb, Y.; Konstantinidis, K.T. Comparison of closely related, uncultivated Coxiella tick endosymbiont population genomes reveals clues about the mechanisms of symbiosis. Environ. Microbiol. 2018, 20, 1751–1764. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ticks | B. henselae-Infected Group | Noninfected Group |
|---|---|---|
| Females (after 14 days of feeding) | n = 25 | n = 20 |
| No. of engorged ticks | 4 (16.0%) | 5 (25.0%) |
| No. of semi-engorged ticks | 14 (56.0%) | 10 (50.0%) |
| No. of unfed, inactive, or dead ticks | 7 (28.0%) | 5 (25.0%) |
| Attachment rate | 18 (72.0%) | 15 (75.0%) |
| Larvae (after 7 days of feeding) | n = 300 | n = 250 |
| No. of engorged ticks | 51 (17.0%) | 39 (15.6%) |
| No. of semi-engorged ticks | 28 (9.3%) | 85 (34.0%) |
| No. of unfed, inactive, or dead ticks | 221 (73.6%) | 126 (50.4%) |
| Attachment rate | 79 (26.3%) * | 124 (49.6%) * |
| Adult Ticks | No. of B. henselae-Positive Samples/No. of Tested Samples (%) | |
|---|---|---|
| B. henselae-Infected Group | Noninfected Group | |
| At the end of feeding | ||
| Males (MG) | 5/8 (62.5) | 0/3 (0) |
| Males (SG) | 3/8 (37.5) | 0/3 (0) |
| Males (CC) | 0/8 (0) | 0/3 (0) |
| Females (MG) | 8/10 (80) | 0/3 (0) |
| Females (SG) | 6/10 (60) | 0/3 (0) |
| Females (OV) | 0/10 (0) | 0/3 (0) |
| Females (CC) | 3/10 (30) | 0/3 (0) |
| Post oviposition | ||
| Females (MG) | 0/3 (0) | 0/2 (0) |
| Females (SG) | 1/3 (33.3) | 0/2 (0) |
| Females (OV) | 0/3 (0) | 0/2 (0) |
| Females (CC) | 0/3 (0) | 0/2 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wechtaisong, W.; Bonnet, S.I.; Chomel, B.B.; Lien, Y.-Y.; Chuang, S.-T.; Tsai, Y.-L. Investigation of Transovarial Transmission of Bartonella henselae in Rhipicephalus sanguineus sensu lato Ticks Using Artificial Feeding. Microorganisms 2021, 9, 2501. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122501
Wechtaisong W, Bonnet SI, Chomel BB, Lien Y-Y, Chuang S-T, Tsai Y-L. Investigation of Transovarial Transmission of Bartonella henselae in Rhipicephalus sanguineus sensu lato Ticks Using Artificial Feeding. Microorganisms. 2021; 9(12):2501. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122501
Chicago/Turabian StyleWechtaisong, Wittawat, Sarah I. Bonnet, Bruno B. Chomel, Yi-Yang Lien, Shih-Te Chuang, and Yi-Lun Tsai. 2021. "Investigation of Transovarial Transmission of Bartonella henselae in Rhipicephalus sanguineus sensu lato Ticks Using Artificial Feeding" Microorganisms 9, no. 12: 2501. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122501