
Fusarium Species in Mangrove Soil in Northern Peninsular Malaysia and the Soil Physico-Chemical Properties

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Identification of Fusarium Isolates
2.2. Molecular Identification and Phylogenetic Analysis
2.2.1. DNA Extraction
2.2.2. PCR Amplification
2.2.3. Sequence and Phylogenetic Analyses
2.3. Physical and Chemical Analyses of Mangrove Soil
2.3.1. Soil Moisture
2.3.2. Soil Texture
2.3.3. Soil pH
2.3.4. Soil Salinity
2.3.5. Carbon Content
2.3.6. Nitrogen Content
2.4. Statistical Analyses
3. Results
3.1. Fusarium Isolates
3.2. PCR Amplification and Sequence Analysis of TEF-1α Gene
3.3. PCR Amplification and Sequence Analysis of β -Tubulin Gene and ITS Region
3.4. Phylogenetic Analysis of FSSC Isolates Based on Combined TEF-1α and ITS Sequences
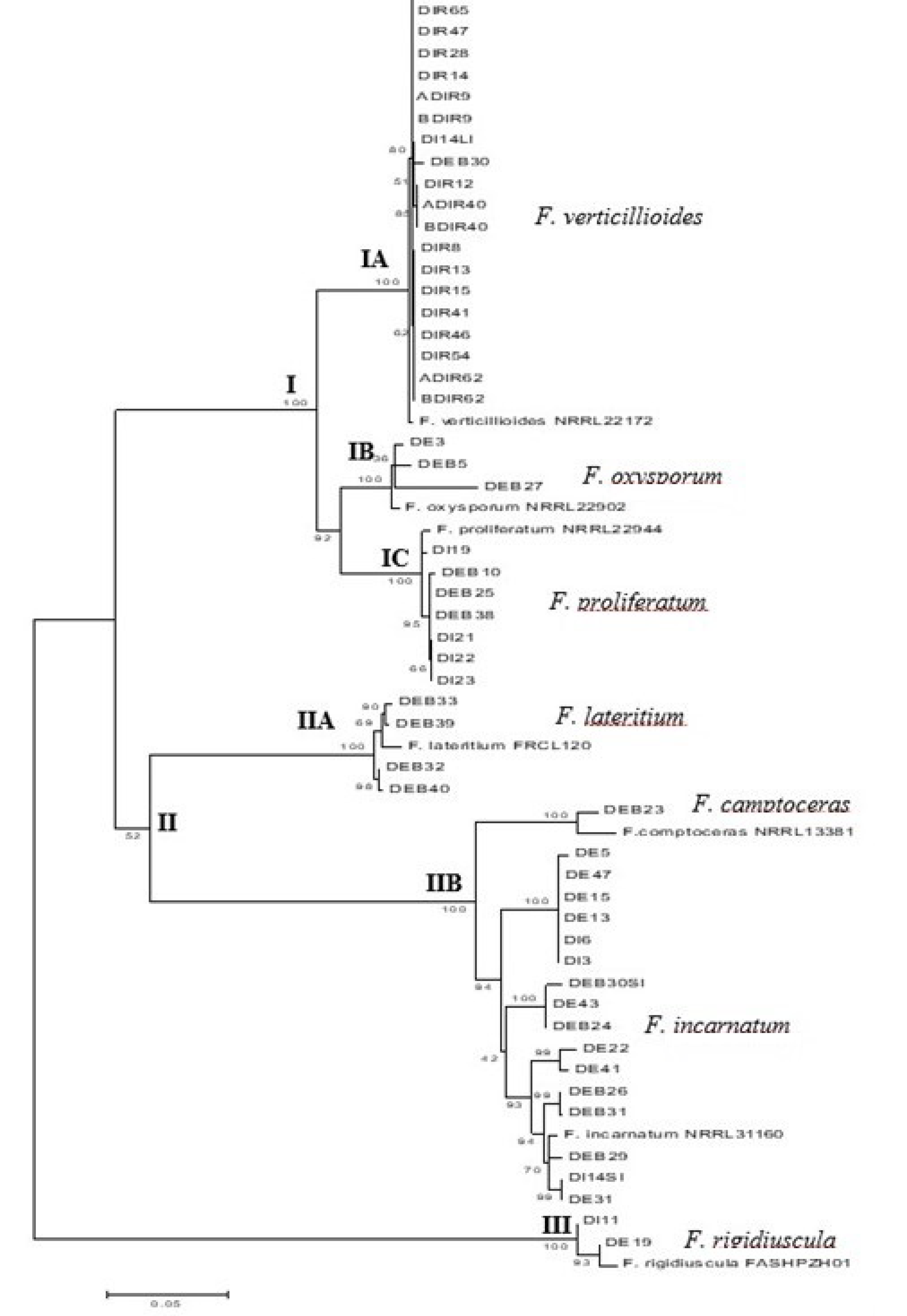
3.5. Phylogenetic Analysis of Fusarium Species Based on Combined TEF-1α and β -Tubulin Sequences
3.6. Soil Analysis
3.6.1. Soil Moisture
3.6.2. Soil Texture
3.6.3. Soil pH
3.6.4. Soil Salinity
3.6.5. Carbon Content
3.6.6. Nitrogen Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Hatmi, A.M.S.; Bonifaz, A.; De Hoog, G.S.; Vazquez-Maya, L.; Garcia-Carmona, K.; Meis, J.F.; Van Diepeningen, A.D. Keratitis by Fusarium temperatum, a novel opportunist. BMC Infect. Dis. 2014, 14, 588. [Google Scholar] [CrossRef] [Green Version]
- Burgess, L.W. General ecology of the Fusaria. In Fusarium: Diseases, Biology, and Taxonomy; Nelson, P.E., Toussoun, T.A., Cook, R.J., Eds.; University Park, The Pennsylvania State University: State College, PA, USA, 1981; pp. 225–235. [Google Scholar]
- Burgess, L.W.; Bryden, W.L. Fusarium: A ubiquitous fungus of global significance. Microbiol. Aust. 2012, 22–25. [Google Scholar] [CrossRef]
- Latiffah, Z.; Izzati, H.N.; Baharuddin, S. Fusarium species isolated from peat soil of Pondok Tanjung and Sungai Beriah, Perak. Malays. J. Microbiol. 2010, 6, 102–105. [Google Scholar] [CrossRef]
- Mandeel, Q.A.; Abbas, J.A. Survey of Fusarium species in an arid environment of Bahrain. Mycopathologia 1994, 127, 167–173. [Google Scholar] [CrossRef]
- Kommendahl, T.; Abbas, H.K.; Barnes, P.M.; Mirocha, C.J. Prevalance and toxigenicity of Fusarium species form soils of Norway near the Arctic Circle. Mycologia 1988, 80, 790–794. [Google Scholar] [CrossRef]
- Ananda, K.; Sridhar, K.R. Diversity of endophytic fungi in the roots of mangrove species on the west coast of India. Can. J. Microbiol. 2002, 48, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Lee, S.Y. Ecology of Mangrove Fungi and Their Role in Mutrient Cycling: What Gaps Occur in Our Knowledge? Hydrobiologia 1995, 295, 107–118. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Bebout, B.; Vlkmann-Kohlmeyer, B. Decomposition of mangrove wood by marine fungi and teredinids in Belize. Mar. Ecol. 1995, 16, 27–39. [Google Scholar] [CrossRef]
- Dittmar, T.; Hertkorn, N.; Kattner, G.; Lara, R.J. Mangroves, a major source of dissolved organic carbon to the oceans. Glob. Biogeochem. Cycles 2006, 20, 1–7. [Google Scholar] [CrossRef]
- Kuthubutheen, A.J. Leaf surface fungi associated with Avicennia alba and Rhizophora mucronata in Malaysia. In Proceedings of the Asian Symposium on Mangrove Environment-Research and Management, Kuala Lumpur, Malaysia, 25–29 August 1980. [Google Scholar]
- Alias, S.A.; Zainuddin, N.; Jones, E.G. Biodiversity of marine fungi in Malaysian mangroves. Bot. Mar. 2010, 53, 545–554. [Google Scholar] [CrossRef]
- Latiffah, Z.; Kok, F.M.; Mei, H.H.; Maziah, Z.; Baharuddin, S. Fusarium species isolated from mangrove soil in Kampung Pantai Acheh, Balik Pulau, Pulau Pinang, Malaysia. Trop. Life Sci. Res. 2010, 21, 21–29. [Google Scholar] [PubMed]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Wiley-Blackwell: Oxford, UK, 2006. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.O.; Cigelnik, E.L.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 Types within a monophyletic lineage of the Fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Tamura, K.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K. Molecular phylogeny of the Nectria haematococca-Fusarium solani species complex. Mycologia 2000, 92, 919–938. [Google Scholar] [CrossRef]
- Proctor, R.H.; Desjardins, A.E.; Plattner, R.D.; Hohn, T.M. A polyketide synthase gene required for biosynthesis of fumonisin mycotoxins in Gibberella fujikuroi mating population A. Fungal Genet. Biol. 1999, 27, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.X.; Zhang, X.; Zhang, H.; Lu, Y.; Yu, Q.; Zhang, H.; Xie, Y. First report of dieback of mango caused by Fusarium decemcellulare in China. J. Phytopathol. 2013, 161, 735–738. [Google Scholar] [CrossRef]
- Geiser, D.M.; Ivey, M.L.; Hakiza, G.; Juba, J.H.; Miller, S.A. Gibberella xylarioides anamorph: Fusarium xylarioides), a causative agent of coffee wilt disease in Africa, is a previously unrecognized member of the G. fujikuroi species complex. Mycologia 2005, 97, 191–201. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.; Kang, S.; Park, B.; Robert, V.A.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic diversity of insecticolous fusaria inferred from multilocus DNA sequence data and their molecular identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef] [Green Version]
- Rowell, D.L. Soil Science: Methods and Applications; Routledge: Harlow, Essex, UK, 1994. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle-size analysis. Methods Biogeochem. Wetl. 2018, 9, 383–411. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of Degtjareff method for determining soil organic matter, and proposed modification of the chromic acid tritation method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Ritieni, A.; Bottalico, A.; Corda, P. Occurrence and toxigenicity of Fusarium proliferatum from preharvest maize ear rot, and associated mycotoxins, in Italy. Plant Dis. 1995, 79, 727–731. [Google Scholar] [CrossRef]
- Nalim, F.A.; Samuels, G.J.; Wijesundera, R.L.; Geiser, D.M. New species from the Fusarium solani species complex derived from perithecia and soil in the Old World tropics. Mycologia 2011, 103, 1302–1330. [Google Scholar] [CrossRef]
- Bogale, M.; Steenkamp, E.; Wingfield, M.; Wingfield, B. Diverse Fusarium solani isolates colonise agricultural environments in Ethiopia. Eur. J. Plant Pathol. 2009, 124, 369–378. [Google Scholar] [CrossRef]
- Chehri, K. Molecular phylogeny of the Fusarium solani species complex (FSSC) isolated from soils in Iran. Botany 2014, 92, 815–820. [Google Scholar] [CrossRef]
- Nirenberg, H.I.; Brielmaier-Liebetanz, U. Nectria ipomoeae Halst., Anamorph: Fusarium striatum Sherb. an Passiflora edulis Sims. Nachr Dtsch Pflanzenschutzd 1996, 48, 270–274. [Google Scholar]
- Rossman, A.Y.; Samuels, G.J.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–248. [Google Scholar]
- Burgess, L.W.; Summerell, B.A. Mycogeography of Fusarium: Survey of Fusarium species from subtropical and semiarid grassland soils from Queensland, Australia. Mycol. Res. 1992, 96, 780–784. [Google Scholar] [CrossRef]
- Mandeel, Q.A. Biodiversity of the genus Fusarium in saline soil habitats. J. Basic Microbiol. 2006, 46, 480–494. [Google Scholar] [CrossRef]
- Nordahliawate, S.; Izzati, N.A.; Azlin, N.; Baharudin, S. Diversity of Fusarium species isolated from soil cultivated with cucurbits within east coast, Peninsular Malaysia. Pertanika J. Trop. Agric. Sci. 2012, 35, 381–386. [Google Scholar]
- Gott, K.P.; Burgess, L.W.; Balmas, V.; Duff, J. Mycogeography of Fusarium: Fusarium species in soils from Palm Valley, Central Australia. Australas. Plant Pathol. 1994, 23, 112–117. [Google Scholar] [CrossRef]
- Summerell, B.; Rugg, C.; Burgess, L. Mycogeography of Fusarium: Survey of Fusarium species associated with forest and woodland communities in north Queensland, Australia. Mycol. Res. 1993, 97, 1015–1019. [Google Scholar] [CrossRef]
- Summerell, B.A.; Leslie, J.F.; Liew, E.C.Y.; Laurence, M.H.; Bullock, S.; Petrovic, T.; Bentley, A.R.; Howard, C.G.; Peterson, S.A.; Walsh, J.L.; et al. Fusarium species associated with plants in Australia. Fungal Divers. 2011, 46, 1–27. [Google Scholar] [CrossRef]
- McMullen, M.P.; Stack, R.W. Effects of isolation techniques and media on the differential isolation of Fusarium species. Phytopathology 1983, 73, 458–462. [Google Scholar] [CrossRef]
- Latiffah, Z.; Mohd Zariman, M.; Baharuddin, S. Diversity of Fusarium species in cultivated soils in Penang. Malays. J. Microbiol. 2007, 3, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, A.; Gilardi, G.; Gullino, M.L. Seed transmission of Fusarium oxysporum f. sp. lactucae. Phytoparasitica 2004, 32, 61–65. [Google Scholar] [CrossRef]
- Xia, J.W.; Sandoval-Denis, M.; Crous, P.W.; Zhang, X.G.; Lombard, L. Numbers to names—restyling the Fusarium incarnatum-equiseti species complex. Persoonia 2019, 43, 186–221. [Google Scholar] [CrossRef] [PubMed]
- Bushula, V.S. Native Fusarium Species from Indigenous Fynbos Soils of the Western Cape. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2008. Available online: http://hdl.handle.net/10019.1/2437 (accessed on 1 November 2020).
- Karim, N.F.A.; Mohd, M.; Nor, N.M.I.M.; Zakaria, L. Saprophytic and potentially pathogenic fusarium species from peat soil in Perak and Pahang. Trop. Life Sci. Res. 2016, 27, 1–20. [Google Scholar] [PubMed]
- Sarquis, M.M.; Borba, C.M. Fusarium species in sandy soil from Ipanema Beach. J. Basic Microbiol. 1997, 37, 425–429. [Google Scholar] [CrossRef]
- Jeschke, N.; Nelson, P.E.; Marasas, W.F.O. Fusarium species isolated from soil samples collected at different altitudes in the Transkei, southern Africa. Mycologia 1990, 82, 727–733. [Google Scholar] [CrossRef]
- Onyike, N.B.N.; Nelson, P.E. The distribution of Fusarium species in soils planted to millet and sorghum in Lesotho, Nigeria and Zimbabwe. Mycopathologia 1993, 121, 105–114. [Google Scholar] [CrossRef]
- Steiman, R.; Guiraud, P.; Sage, L.; Seigle-Murandi, F. Soil mycoflora from the Dead Sea Oases of Ein Gedi and Einot Zuqim (Israel). Antonie Leeuwenhoek 1997, 72, 261–270. [Google Scholar] [CrossRef]
- Chehri, K. First report of three new Fusarium species isolated from garden soil in southern Iran. Rostaniha 2015, 16, 81–87. [Google Scholar]
- Kara, Ö.; Bolat, I. Influence of soil compaction on microfungal community structure in two soil types in Bartin Province, Turkey. J. Basic Microbiol. 2007, 47, 394–399. [Google Scholar] [CrossRef]
- Panda, T.; Panda, B.; Prasad, B.; Mishra, N. Influence of soil environment and surface vegetation on soil micro flora in a coastal Sandy Belt of Orissa, India. J. Hum. Ecol. 2009, 27, 69–73. [Google Scholar] [CrossRef]
- Hoorman, J.J.; Islam, R. Understanding Soils Microbes and Nutrient Recycling; Agriculture and Natural Resources, Ohio State University Extension: Columbus, OH, USA, 2010; Available online: http://ohioline.osu.edu/factsheet/SAG-16 (accessed on 1 November 2020).
- Swer, H.; Dkhar, M.S.; Kayang, H. Fungal population and diversity in organically amended agricultural soils of Meghalaya, India. J. Org. Syst. 2011, 6, 3–12. [Google Scholar]
- Fargasova, A. The influence of various nitrogen-sources on the growth, conidiation and pigment production of the brown mutant Trichoderma-VIRIDE M-108. Biologia 1992, 47, 453–464. [Google Scholar]
- Joshi, I.J.; Chauhan, R.K.S. Distribution of soil microfungi in various soil types of Chambal Ravines. Proc. Indian Natl. Sci. Acad. 1982, 48, 525–533. [Google Scholar]
- Rousk, J.; Brookes, P.; Bååth, E. Contrasting soil pH effects on fungal and growth suggest functional redundancy in carbon mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Acosta, B.; Bashan, Y.; Hernandez-Saavedra, N.Y.; Ascencio, F.; De La Cruz-Agüero, G. Seasonal seawater temperature as the major determinant for populations of culturable bacteria in the sediments of an intact mangrove in an arid region. FEMS Microbiol. Ecol. 2006, 55, 311–321. [Google Scholar] [CrossRef]
- Thatoi, H.N.; Behera, B.C.; Dangar, T.K.; Mishra, R.R. Microbial biodiversity in mangrove soils of Bhitarkanika, Odisha, India. Int. J. Environ. Biol. 2012, 2, 50–58. [Google Scholar]
- Wheeler, K.A.; Hurdman, B.F.; Pitt, J. Influence of pH on the growth of some toxigenic species of Aspergillus, Penicillium and Fusarium. Int. J. Food Microbiol. 1991, 12, 141–149. [Google Scholar] [CrossRef]
- Nevarez, L.; Vasseur, V.; Le Madec, A.; Le Bras, M.; Coroller, L.; Leguérinel, I.; Barbier, G. Physiological traits of Penicillium glabrum strain LCP 08.5568, a filamentous fungus isolated from bottled aromatised mineral water. Int. J. Food Microbiol. 2009, 130, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Nazim, K.; Ahmed, M.; Shaukat, S.S.; Khan, M.U.; Rao, T.A.; Ali, Q.M.; Sherwani, S.K. Distribution and diversity of airborne microflora under mangrove forest at Sandspit Area Karachi, Pakistan. Sci. Technol. Dev. 2012, 31, 305–311. [Google Scholar]
- Maina, P.; Okoth, S.; Monda, E. Impact of land use on distribution and diversity of Fusarium spp. in Taita Taveta, Kenya. Trop. Subtrop. Agroecosyst. 2009, 11, 323–335. [Google Scholar]
- Banakar, S.P.; Thippeswamy, B.; Thirumalesh, B.V.; Naveenkumar, K.J. Diversity of soil fungi in dry deciduous forest of Bhadra Wildlife sanctuary, western Ghats of southern India. J. For. Res. 2012, 23, 631–640. [Google Scholar] [CrossRef]
- Oritsejafor, J. Influence of moisture and pH on growth and survival of Fusarium oxysporum f.sp. elaeidis in soil. Trans. Br. Mycol. Soc. 1986, 87, 511–517. [Google Scholar] [CrossRef]
- Gauthier, G.M.; Keller, N.P. Crossover fungal pathogens: The biology and pathogenesis of fungi capable of crossing kingdoms to infect plants and humans. Fungal Genet. Biol. 2013, 61, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Nucci, M.; Anaissie, E. Fusarium infections in immunocompromised patients. Clin. Microbiol. Rev. 2007, 20, 695–704. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Gueidan, C.; Crous, P.W.; Geiser, D.M. Novel multilocus sequence typing scheme reveals high genetic diversity of human pathogenic members of the Fusarium incarnatum-F. equiseti and F. chlamydosporum species complexes within the United States. J. Clin. Microbiol. 2009, 47, 3851–3861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backhouse, D.; Burgess, L.W. Mycogegraphy of Fusarium climatic: Analysis of the distribution within Australia of Fusarium species in section Gibbosum. Mycol. Res. 1995, 99, 1218–1224. [Google Scholar] [CrossRef]
- Smith, S.N. An overview of ecological and habitat aspects in the genus Fusarium with special emphasis on the soil-borne pathogenic forms. Plant Pathol. Bull. 2007, 16, 97–120. [Google Scholar]



{kind=link}
{kind=link}
| Species/Code | Accession No. | Geographic Origin | References |
|---|---|---|---|
| F. verticillioides | NRRL22172 | Germany | O’Donnell et al. [19] |
| F. oxysporum | NRRL22902 | USA | O’Donnell et al. [19] |
| F. proliferatum | NRRL22944 | Germany | O’Donnell et al. [19] |
| F. camptoceras | NRRL13381 | - | Proctor et al. [20] |
| F. incarnatum | NRRL31160 | - | Proctor et al. [20] |
| F. rigidiuscula | FASHPZH01 | - | Qi et al. [21] |
| F. lateritium | FRCL120 | Africa | Geiser et al. [22] |
| F. chlamydosporum | NRRL 52797 | India | O’Donnell et al. [23] |
| Code | Accession Number | Fusarium solani | Geographic Origin |
|---|---|---|---|
| NRRL22389 | AF178340 | Fusarium sp. | Maryland, USA |
| NRRL22142 | AF178347 | Fusarium solani f. sp. cucurbitae MPV | California, USA |
| NRRL 22141 | AF178329 | Fusarium solani f. sp. cucurbitae MPV | New Zealand |
| NRRL22161 | AF178330 | Fusarium solani f. sp. robiniae MPVII | Japan |
| NRRL22586 | AF178353 | Fusarium solani f. sp. robiniae MPVII | Virginia, USA |
| NRRL22157 | AF178359 | Fusarium solani f. sp. mori MPVIII | Japan |
| NRRL22230 | AF178358 | Fusarium solani f. sp. mori MPVIII | Japan |
| NRRL22278 | AF178337 | Fusarium solani f. sp. pisi MPVI | - |
| NRRL22820 | AF178355 | Fusarium solani f. sp. pisi MPVI | Indiana, USA |
| NRRL22579 | AF178352 | Fusarium sp. | Indonesia |
| NRRL22163 | AF178382 | Fusarium solani f. sp. xanthoxyli MPIV | Japan |
| NRRL22277 | AF178336 | Fusarium solani f. sp. xanthoxyli MPIV | Japan |
| NRRL22402 | AF178344 | Fusarium solani f. sp. batatas | North Carolina, USA |
| NRRL22400 | AF178343 | Fusarium solani f. sp. batatas | North Carolina, USA |
| NRRL22101 | AF178333 | Fusarium striatum | Panama |
| NRRL22166 | AF178350 | Neocosmospora vasinfecta | Illinois, USA |
| NRRL22436 | AF178348 | Neocosmospora vasinfecta | South Africa |
| NRRL22468 | AF178349 | Neocosmospora vasinfecta | Guinea |
| NRRL22178 | AF178334 | Fusarium sp. | Venezuela |
| NRRL22570 | AF178360 | Fusarium sp. | Brazil |
| NRRL22098 | AF178327 | Fusarium solani f. sp. cucurbitae MPI | - |
| NRRL22153 | AF178346 | Fusarium solani f. sp. cucurbitae MPI | - |
| NRRL20438 | AF178332 | Fusarium ambrosium | India |
| NRRL22354 | AF178338 | Fusarium sp. | French Guiana |
| NRRL22396 | AF178342 | Fusarium sp. | French Guiana |
| NRRL22395 | AF178341 | Fusarium sp. | Venezuela |
| NRRL22412 | AF178351 | Fusarium sp. | French Guiana |
| NRRL22387 | AF178339 | Fusarium sp. | French Guiana |
| NRRL22574 | AF178345 | Fusarium sp. | Guatemala |
| NRRL22158 | AF178331 | Fusarium martii-phaseoli | New York, USA |
| NRRL22275 | AF178335 | Fusarium martii-phaseoli | - |
| NRRL22823 | AF178356 | Fusarium solani f. sp. glycines | Indiana, USA |
| NRRL22825 | AF178357 | Fusarium solani f. sp. glycines | Indiana, USA |
| NRRL22632 | AF178354 | Nectria plagianthi | New Zealand |
| NRRL22090 | AF178326 | Nectria illudens | New Zealand |
| NRRL22316 | AF178361 | Fusarium staphyleae | New Jersey, USA |
| State (Location) | Sampling Site | Soil Texture | Carbon Content (mg/g) | Nitrogen Content (mg/g) | Soil pH | Soil Moisture (%) | Soil Salinity (ppt) | Fusarium Species (Number of Isolates) | Total |
|---|---|---|---|---|---|---|---|---|---|
| Penang (Pantai Acheh, Balik Pulau | PG1 | Loamy sand | 71.8 ± 0.9 | 2.3 ± 0.1 | 7.33 ± 0.01 | 2.59 | 17.39 ± 4.7 | F. solani (3) F. lateritium (2) F. proliferatum (1) | 48 (35%) |
| PG2 | Sandy loam | 60.96 ± 1.2 | 1.8 ± 0 | 7.53 ± 0.39 | 3.71 | 16.49 ± 1.4 | F. solani (1) F. longipes (1) | ||
| PG3 | Sandy loam | 49.52 ± 0.6 | 1.9 ± 0 | 7.29 ± 0.01 | 3.07 | 21.36 ± 0.6 | F. solani (22) F. verticillioides (18) | 18 (13%) | |
| Penang (Pulau Burung, Seberang Perai) Pinang (Pulau Burung, Seberang Perai) | PB1 | Sandy loam | 138 ± 1.6 | 4.0±0 | 6.22±0.19 | 3.64 | 16.72 ± 0.5 | F. lateritium (2) | |
| PB2 | Sandy loam | 84.5 ± 0.3 | 1.33 ± 0.4 | 7.14 ± 0.06 | 2.43 | 16.24 ± 0.05 | F. oxysporum (1) | ||
| PB3 | Sandy loam | 113 ± 8.6 | 3.4 ± 0.2 | 7.02 ± 0.06 | 3.3 | 19.08 ± 2.9 | F. solani (4) F. verticillioides (1) F. incarnatum (5) F. proliferatum (2) F. oxysporum (1) F. chlamydosporum (1) F. camptoceras (1) | ||
| Kedah Kampung Pantai Merdeka | KPM1 | Sandy loam | 31.93 ± 0.7 | 1.7 ± 0.2 | 6.61 ± 0.01 | 3.6 ± 0.5 | 18.91 ± 4.3 | F. solani (4) | 13 (9%) |
| KPM2 | Sandy loam | 21.03 ± 1.2 | 1.33 ± 0.1 | 6.12 ± 0.05 | 1.83 ± 0 | 18.31 ± 0.1 | F. solani (5) F. incarnatum (3) F. oxysporum (1) | ||
| KPM3 | Loam | 19.45 ± 0.8 | 1.6 ± 0.2 | 5.97 ± 0.01 | 2.14 ± 0.3 | 16.52 ± 0.22 | 0 | ||
| Kedah Kampung Batu Lintang | KBL1 | Silt loam | 35.58 ± 0.8 | 2.46 ± 0.1 | 5.72 ± 0.02 | 4.82 ± 0.4 | 10.61 ± 0.5 | F. solani (13) F. incarnatum (3) F. rigidiuscula (1) | 17 (12%) |
| KBL2 | Sandy loam | 47.36 ± 2.7 | 2.46 ± 0.1 | 5.4 ± 0.03 | 3.9 ± 0.2 | 17.62 ± 0.07 | 0 | ||
| KBL3 | Sandy loam | 32.016 ± 1 | 2.0 ± 0 | 5.27 ± 0.09 | 2.77 ± 0.1 | 13.37 ± 0.02 | F. merismoides (1) | ||
| Kedah Semeling1 Sungai Merbok | S1SM1 | Clay loam | 28.61 ± 2.5 | 2.6 ± 0 | 5.02 ± 0.01 | 1.62 ± 0.4 | 11.99 ± 0.6 | F. solani (1) | 2 (1.4%) |
| S1SM2 | Sandy loam | 58. ± 27 ± 1 | 1.6 ± 0 | 5.33 ± 0.02 | 3.08 ± 0.4 | 13.2 ± 0.22 | 0 | ||
| S1SM3 | Sandy loam | 28.86 ± 0.1 | 1.8±0.1 | 5.36±0 | 1.7±0.08 | 10.25±0.04 | F. rigidiuscula (1) | ||
| Kedah Semeling2 Sungai Merbok | S2SM 1 | Sandy clay loam | 45.59 ± 0.4 | 2.1 ± 0.11 | 5.31 ± 0.01 | 2.66 ± 0.2 | 12.78 ± 0.6 | F. solani (4) F. incarnatum (2) | 20 (14%) |
| S2SM 2 | Sandy clay loam | 42.24 ± 1.5 | 2.4 ± 0 | 5.11 ± 0 | 2.66 ± 0.2 | 11.89 ± 0.14 | F. solani (5) F. verticillioides (1) F. incarnatum (2) | ||
| S2SM 3 | Sandy loam | 27.69 ± 7.5 | 2 ± 0 | 5.57 ± 0.03 | 1.82 ± 0.2 | 10.02 ± 0.15 | F. solani (1) F. longipes (1) | ||
| Kedah Segantang Garam h Segantang Garam | KSG1 | Loam | 20.4 ± 0.13 | 2.06 ± 0.1 | 5.98 ± 0.07 | 4.21 ± 0.1 | 13.02 ± 0.3 | F. solani (1) | 18 (13%) |
| KSG2 | Silt loam | 42.75 ± 0.7 | 1.6 ± 0 | 6.12 ± 0.01 | 2.94 ± 0.0 | 11.36 ± 0.8 | F. solani (10) F. incarnatum (1) F. proliferatum (2) F. longipes (1) | ||
| KSG3 | Silt loam | 26.85 ± 0.5 | 2.06 ± 0.1 | 6.133 ± 0 | 4.0±0.7 | 13.33±0.3 | F. proliferatum (3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed Zubi, W.S.; Mohd, M.H.; Mohamed Nor, N.M.I.; Zakaria, L. Fusarium Species in Mangrove Soil in Northern Peninsular Malaysia and the Soil Physico-Chemical Properties. Microorganisms 2021, 9, 497. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030497
Mohamed Zubi WS, Mohd MH, Mohamed Nor NMI, Zakaria L. Fusarium Species in Mangrove Soil in Northern Peninsular Malaysia and the Soil Physico-Chemical Properties. Microorganisms. 2021; 9(3):497. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030497
Chicago/Turabian StyleMohamed Zubi, Wafa S., Masratul Hawa Mohd, Nik Mohd Izham Mohamed Nor, and Latiffah Zakaria. 2021. "Fusarium Species in Mangrove Soil in Northern Peninsular Malaysia and the Soil Physico-Chemical Properties" Microorganisms 9, no. 3: 497. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030497