
Bacteriophages to Control Multi-Drug Resistant Enterococcus faecalis Infection of Dental Root Canals

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of E. faecalis from Root Canal
2.1.1. Patient Selection
2.1.2. Sampling Procedures
2.2. Biochemical Identification of E. faecalis
2.3. Antimicrobial Susceptibility Test
2.4. PCR Analysis of E. faecalis Genomic DNAs
2.5. Isolation of Bacteriophages Infecting E. faecalis
2.6. Bacteriophages Purification and Propagation
2.7. Determination of Host Range and Cross Infectivity of the Isolated Bacteriophages
2.8. Efficiency of Plating (EOP)
2.9. Determination of the Frequency of Bacteriophage Insensitive Mutants
2.10. Examination of Phage Morphology by Electron Microscopy
2.11. Assessment of Phage Lytic Activity in Biofilms
2.12. One-Step Growth Curve
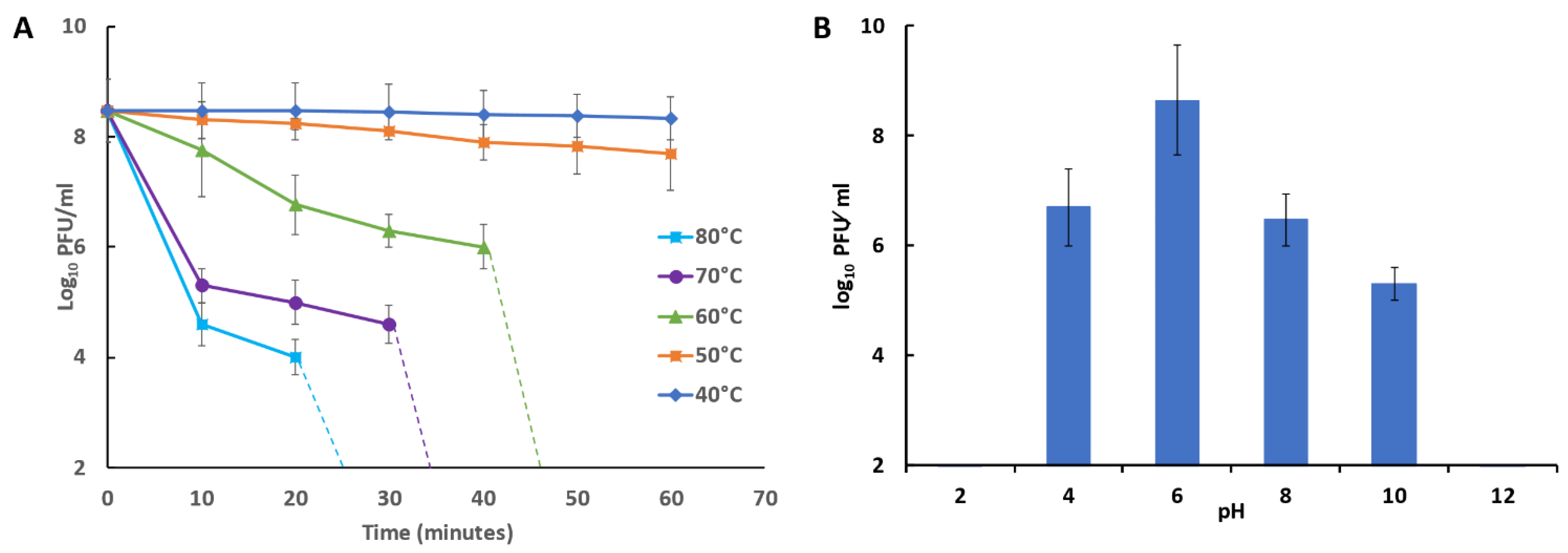
2.13. Bacteriophage Temperature and pH Stability
2.14. Isolation and Characterization of Nucleic Acids from Phage Particles
2.15. Phages Genome Sequencing
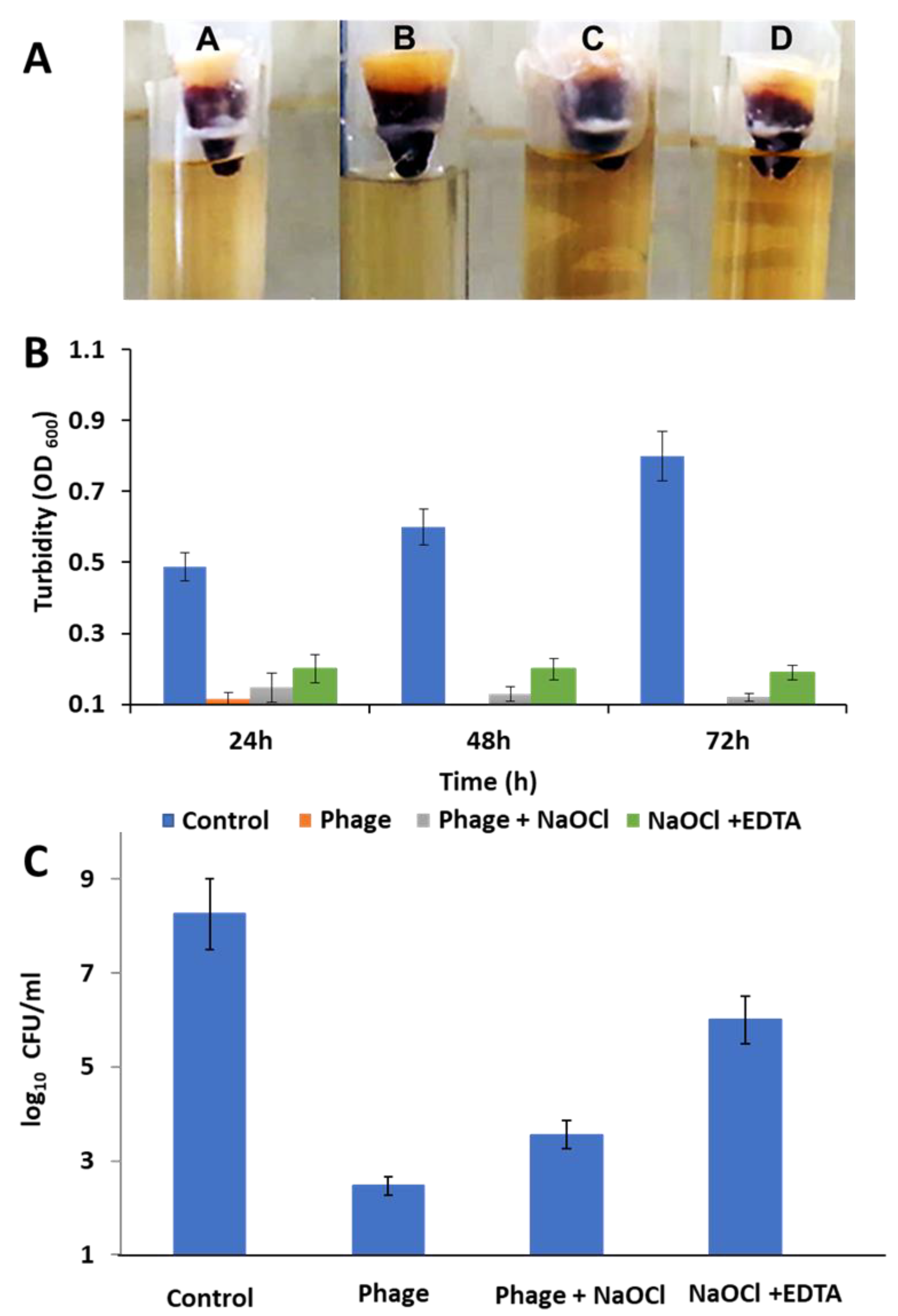
2.16. Ex Vivo Human Root Canal Model
2.16.1. Tooth Selection and Preparation
2.16.2. Tooth Contamination and Obturation
- Group A: saline irrigation as a negative control
- Group B: 200 µL of phage (108 PFU/mL) irrigation
- Group C: 2.5% NaOCl and 200 µL of phage (108 PFU/mL) irrigation
- Group D: 2.5% NaOCl and 17% EDTA irrigation
2.17. Statistical Analysis
3. Results
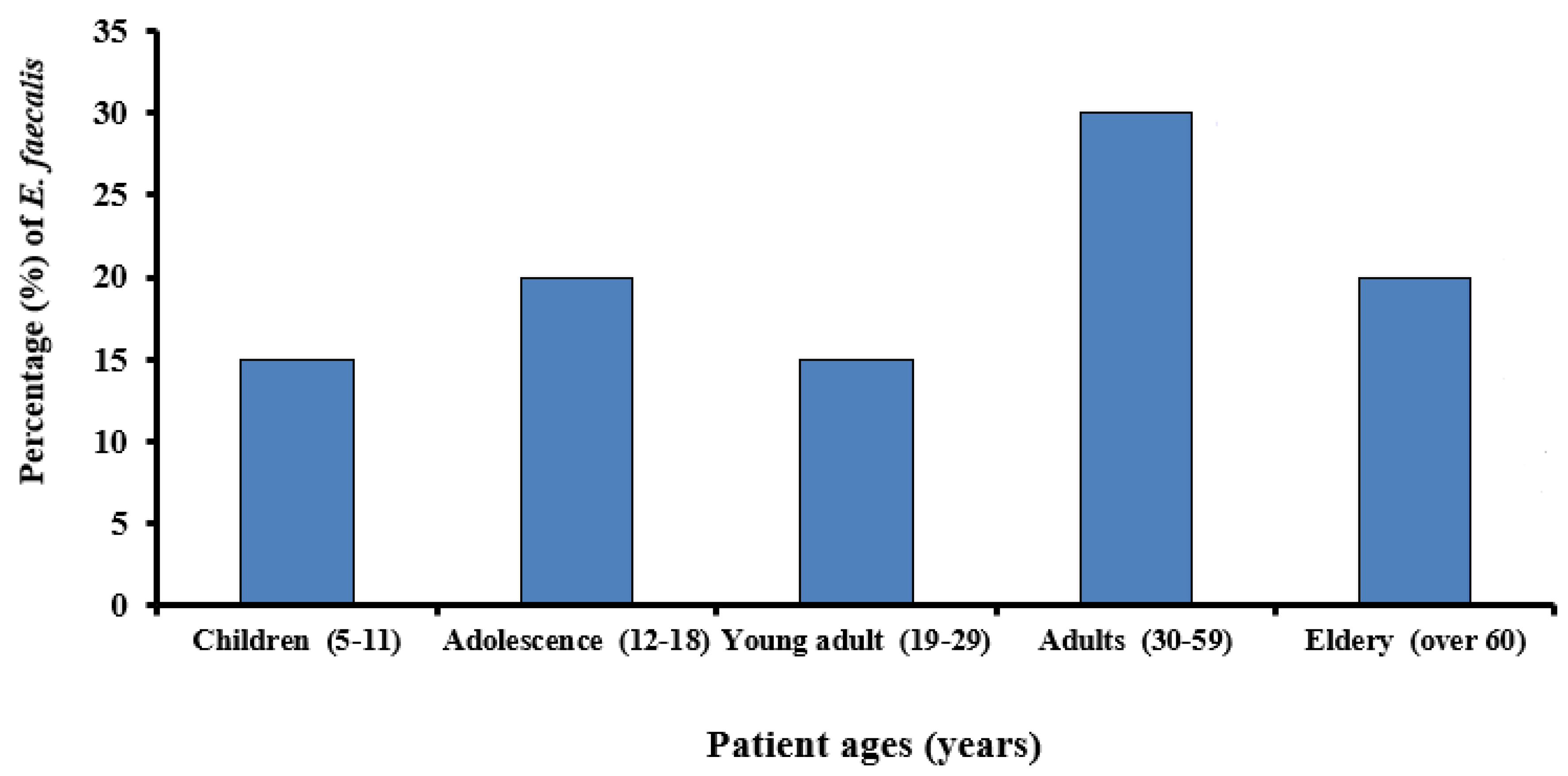
3.1. Prevalence of E. faecalis in Root Canals
3.2. Sensitivity of E. faecalis to Antibiotics
3.3. Survey of Virulence Determinants in E. faecalis Isolates
3.4. Isolation of Bacteriophages
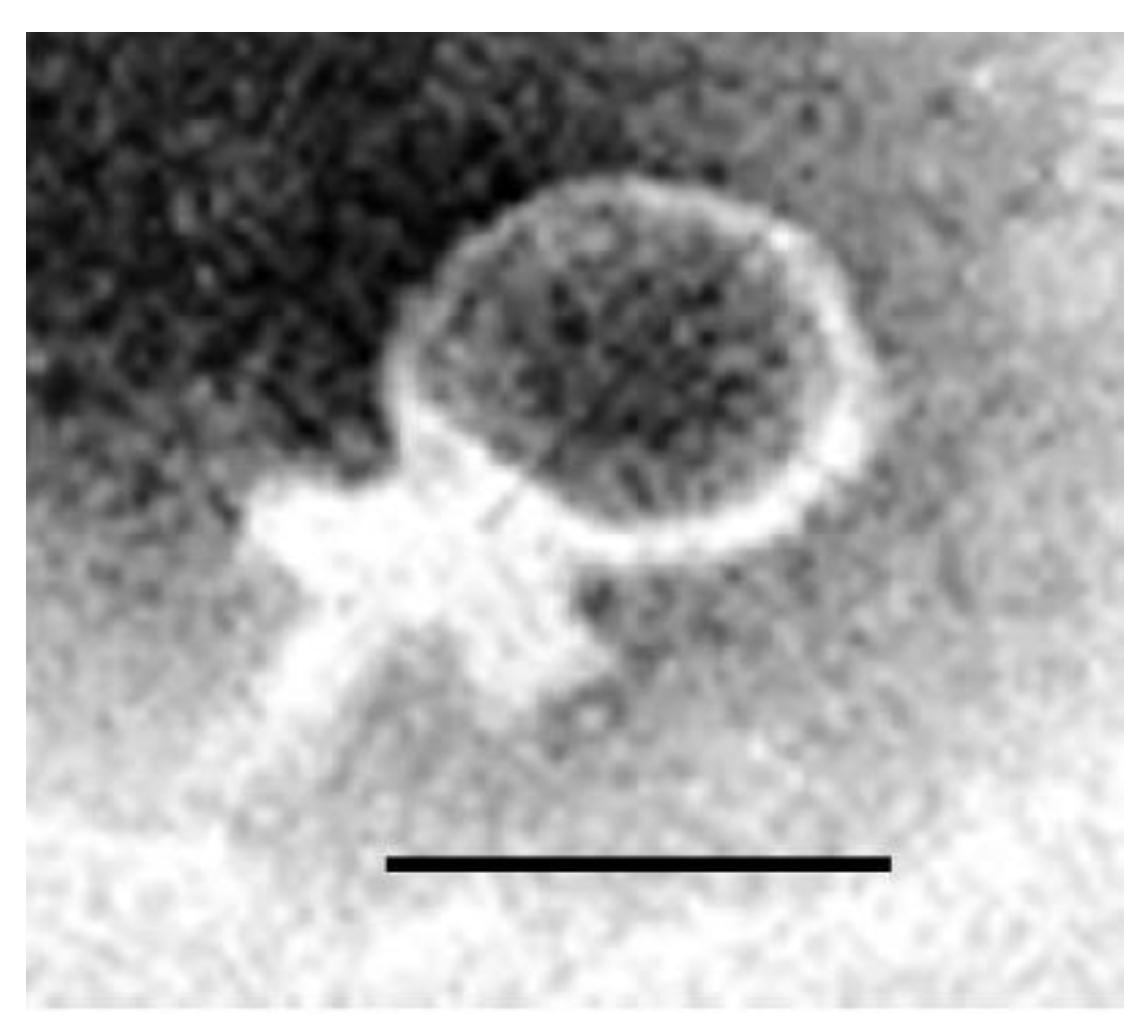
3.5. Morphological Characterization of Phages by Electron Microscopy
3.6. Host Range and EOP of vB_ZEFP Phage
3.7. Frequency of Bacteriophage Insensitive Mutants
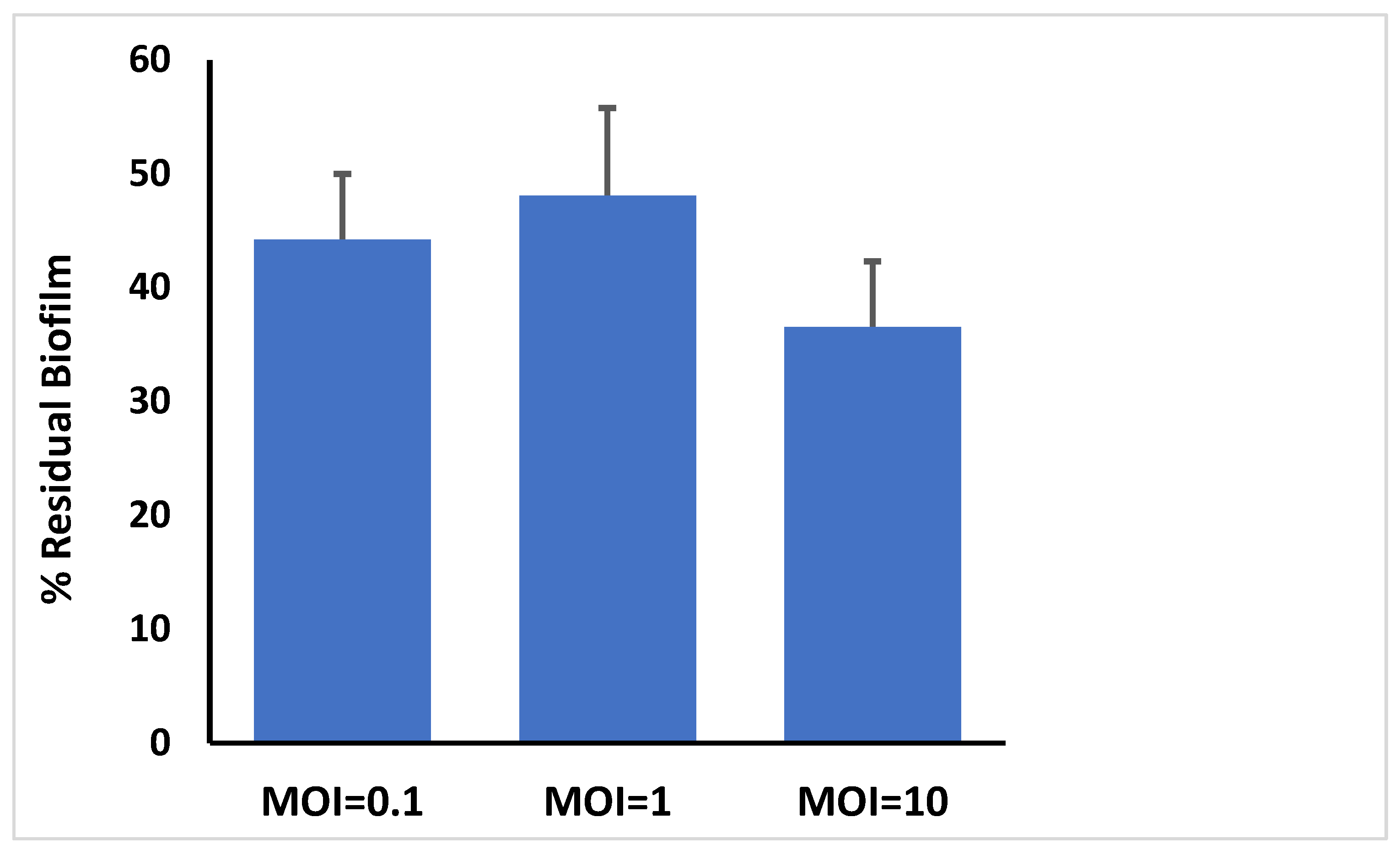
3.8. Bacteriophage Activity Against E. faecalis Established in Biofilms
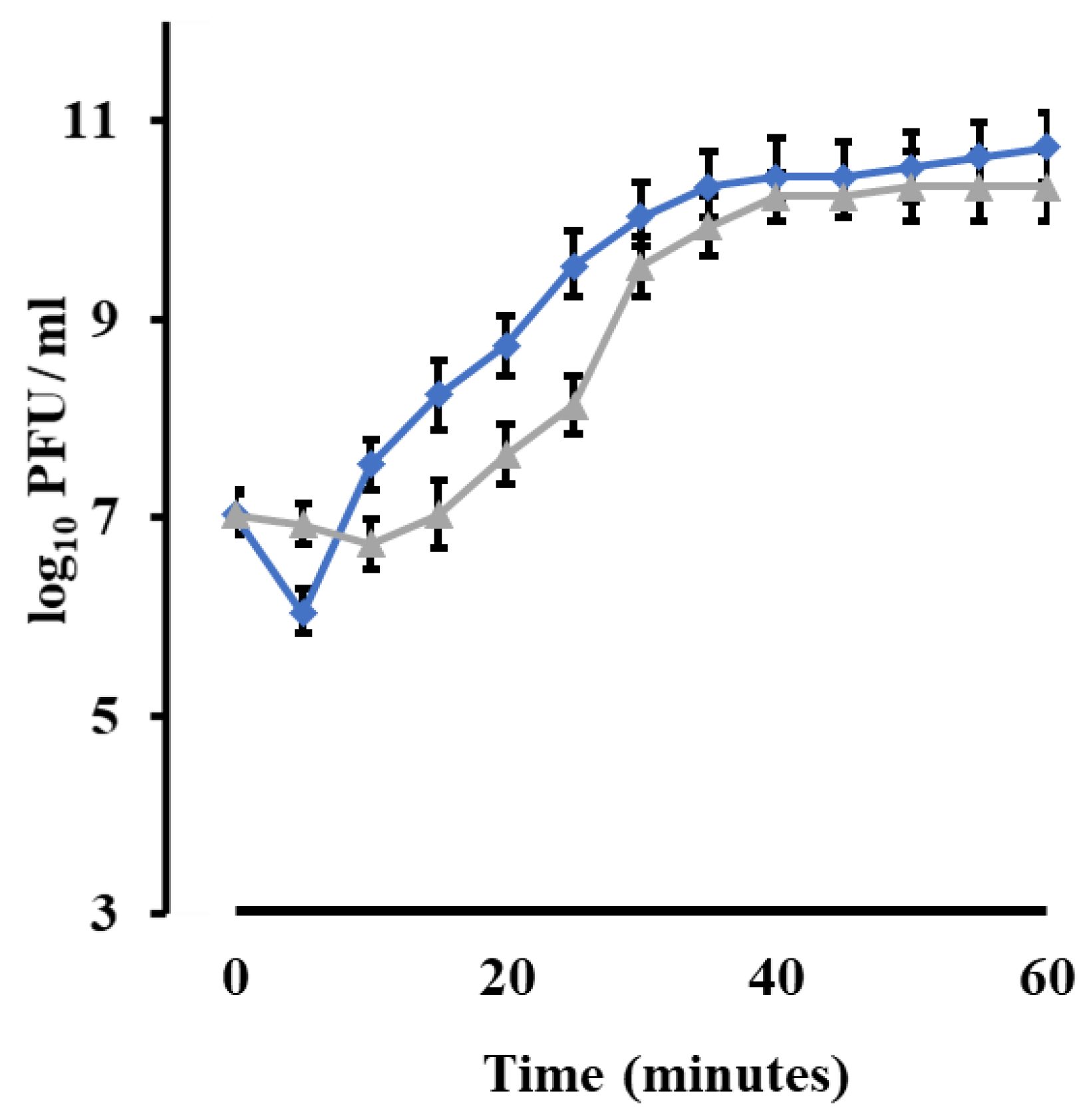
3.9. Phage Growth Characteristics
3.10. Bacteriophage Temperature and pH Stability
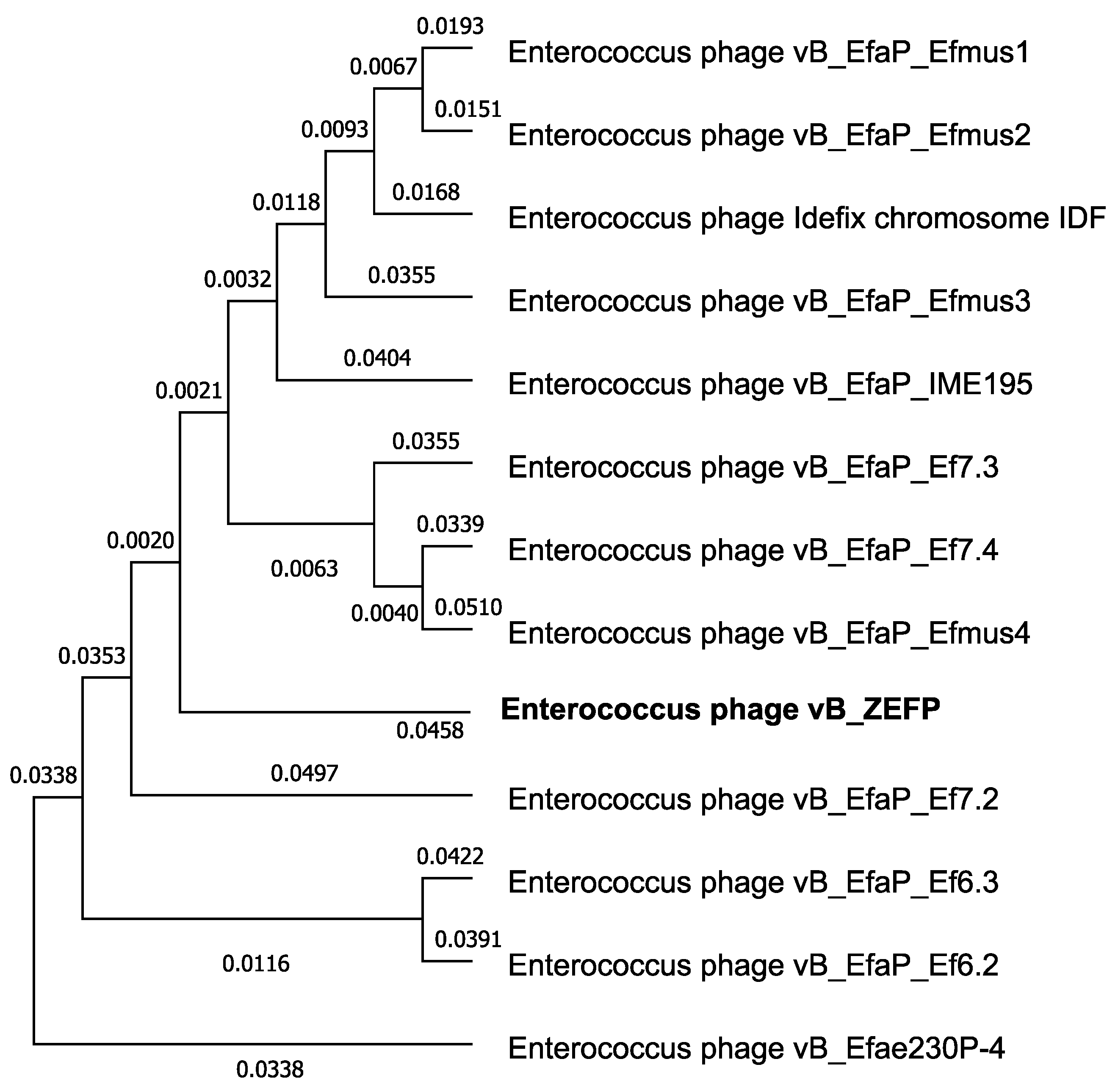
3.11. Phage Genome
3.12. Ex Vivo Human Root Canal Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dahl, A.; Bruun, N.E. Enterococcus faecalis infective endocarditis: Focus on clinical aspects. Expert Rev. Cardiovasc. Ther. 2013, 11, 1247–1257. [Google Scholar] [CrossRef]
- Molander, A.; Reit, C.; Dahlen, G.; Kvist, T. Microbiological status of root-filled teeth with apical periodontitis. Int. Endod. J. 1998, 31, 1–7. [Google Scholar] [CrossRef]
- Wang, Q.Q.; Zhang, C.F.; Chu, C.H.; Zhu, X.F. Prevalence of Enterococcus faecalis in saliva and filled root canals of teeth associated with apical periodontitis. Int. J. Oral Sci. 2012, 4, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siqueira, J.F., Jr.; Rocas, I.N.; Ricucci, D.; Hulsmann, M. Causes and management of post-treatment apical periodontitis. Br. Dent. J. 2014, 216, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebruegge, M.; Pantazidou, A.; Clifford, V.; Gonis, G.; Ritz, N.; Connell, T.; Curtis, N. The age-related risk of coexisting meningitis in children with urinary tract infection. PLoS ONE 2011, 6, e26576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, S.; Hufnagel, M.; Theilacker, C.; Huebner, J. Enterococcal infections: Host response, therapeutic, and prophylactic possibilities. Vaccine 2004, 22, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agudelo Higuita, N.I.; Huycke, M.M. Enterococcal disease, epidemiology, and implications for treatment. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Guzman Prieto, A.M.; van Schaik, W.; Rogers, M.R.; Coque, T.M.; Baquero, F.; Corander, J.; Willems, R.J.L. Global emergence and dissemination of enterococci as nosocomial pathogens: Attack of the clones? Front. Microbiol. 2016, 7, 788. [Google Scholar] [CrossRef] [Green Version]
- Al-Ahmad, A.; Müller, N.; Wiedmann-Al-Ahmad, M.; Sava, I.; Hübner, J.; Follo, M.; Schirrmeister, J.; Hellwig, E. Endodontic and salivary isolates of Enterococcus faecalis integrate into biofilm from human salivary bacteria cultivated in vitro. J. Endod. 2009, 35, 986–991. [Google Scholar] [CrossRef]
- Ran, S.; Wang, J.; Jiang, W.; Zhu, C.; Liang, J. Assessment of dentinal tubule invasion capacity of Enterococcus faecalis under stress conditions ex vivo. Int. Endod. J. 2015, 48, 362–372. [Google Scholar] [CrossRef]
- Bystrom, A.; Sundqvist, G. Bacteriologic evaluation of the efficacy of mechanical root canal instrumentation in endodontic therapy. Scand. J. Dent. Res. 1981, 89, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Sjogren, U.; Figdor, D.; Spangberg, L.; Sundqvist, G. The antimicrobial effect of calcium hydroxide as a short-term intracanal dressing. Int. Endod. J. 1991, 24, 119–125. [Google Scholar] [CrossRef]
- Vivacqua-Gomes, N.; Gurgel-Filho, E.D.; Gomes, B.P.; Ferraz, C.C.; Zaia, A.A.; Souza-Filho, F.J. Recovery of Enterococcus faecalis after single- or multiple-visit root canal treatments carried out in infected teeth ex vivo. Int. Endod. J. 2005, 38, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.S.; Queiroz, L.L.; Campos, P.A.; Batistao, D.W.; Silva, H.A.; Agostini, G.G.; Filho, P.P.G.; Ribas, R.N. The nares as a CA-MRSA reservoir in the healthy elderly. Rev. Soc. Bras. Med. Trop. 2015, 48, 614–616. [Google Scholar] [CrossRef] [Green Version]
- Coque, T.M.; Patterson, J.E.; Steckelberg, J.M.; Murray, B.E. Incidence of hemolysin, gelatinase, and aggregation substance among enterococci isolated from patients with endocarditis and other infections and from feces of hospitalized and community-based persons. J. Infect. Dis. 1995, 171, 1223–1229. [Google Scholar] [CrossRef]
- Lowe, A.M.; Lambert, P.A.; Smith, A.W. Cloning of an Enterococcus faecalis endocarditis antigen: Homology with adhesins from some oral streptococci. Infect. Immun. 1995, 63, 703–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, T.J.; Gasson, M.J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.C.; Jonas, D.; Huber, I.; Karygianni, L.; Wölber, J.; Hellwig, E.; Arweiler, N.; Vach, K.; Wittmer, A.; Al-Ahmad, A. Enterococcus faecalis from food, clinical specimens, and oral sites: Prevalence of virulence factors in association with biofilm formation. Front. Microbiol. 2016, 6, 1534. [Google Scholar] [CrossRef] [Green Version]
- Huycke, M.M.; Spiegel, C.A.; Gilmore, M.S. Bacteremia caused by hemolytic, high-level gentamicin-resistant Enterococcus faecalis. Antimicrob. Agents Chemother. 1991, 35, 1626–1634. [Google Scholar] [CrossRef] [Green Version]
- Chow, J.W.; Thal, L.A.; Perri, M.B.; Vazquez, J.A.; Donabedian, S.M.; Clewell, D.B.; Zervos, M.J. Plasmid-associated hemolysin and aggregation substance production contribute to virulence in experimental enterococcal endocarditis. Antimicrob. Agents Chemother. 1993, 37, 2474–2477. [Google Scholar] [CrossRef] [Green Version]
- Ike, Y.; Hashimoto, H.; Clewell, D.B. Hemolysin of Streptococcus faecalis subspecies zymogenes contributes to virulence in mice. Infect. Immun. 1984, 45, 528–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Ho, K. Bacteriophage therapy for bacterial infections: Rekindling a memory from the pre-antibiotics era. Perspect. Biol. Med. 2001, 44, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Rashel, M.; Uchiyama, J.; Sakurai, S.; Ujihara, T.; Kuroda, M.; Imai, S.; Ikeuchi, M.; Tani, T.; Fujieda, M.; et al. Bacteriophage therapy: A revitalized therapy against bacterial infectious diseases. J. Infect. Chemother. 2005, 11, 211–219. [Google Scholar] [CrossRef]
- Lederberg, J. Smaller fleas… ad infinitum: Therapeutic bacteriophage redux. Proc. Natl. Acad. Sci. USA 1996, 93, 3167–3168. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.R.; March, J.B. Bacteriophages and biotechnology: Vaccines, gene therapy and antibacterials. Trends Biotechnol. 2006, 24, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Ly-Chastain, M.H. The factors affecting effectiveness of treatment in phages therapy. Front. Microbiol. 2014, 5, 51. [Google Scholar]
- Capparelli, R.; Nocerino, N.; Iannaccone, M.; Ercolini, D.; Parlato, M.; Chiara, M.; Iannelli, D. Bacteriophage therapy of Salmonella enterica: A fresh appraisal of bacteriophage therapy. J. Infect. Dis. 2010, 201, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Hughes, K.A.; Sutherland, I.W.; Jones, M.V. Biofilm susceptibility to bacteriophage attack: The role of phage-borne polysaccharide depolymerase. Microbiology 1998, 144, 3039–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häusler, T. Viruses Versus Superbugs: A Solution to the Antibioticscrisis? 2nd ed.; Palgrave Macmillan: New York, NY, USA, 2006. [Google Scholar]
- Khalifa, L.; Shlezinger, M.; Beyth, S.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Phage therapy against Enterococcus faecalis in dental root canals. J. Oral Microbiol. 2016, 8, 32157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlton, R.M. Phage therapy: Past history and future prospects. Arch. Immunol. Ther. Exp. 1999, 47, 267–274. [Google Scholar]
- El-Shibiny, A.; El-Sahhar, S.; Adel, M. Phage applications for improving food safety and infection control in Egypt. J. Appl. Microbiol. 2017, 123, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Peciuliene, V.; Balciuniene, I.; Eriksen, H.M.; Haapasalo, M. Isolation of Enterococcus faecalis in previously root-filled canals in a Lithuanian population. J. Endod. 2000, 26, 593–595. [Google Scholar] [CrossRef]
- Zoletti, G.O.; Siqueira, J.F., Jr.; Santos, K.R.N. Identification of Enterococcus faecalis in root-filled teeth with or without periradicular lesions by culture-dependent and independent approaches. J. Endod. 2006, 32, 722–728. [Google Scholar] [CrossRef]
- Johnson, W.; Kulild, J. Obturation of the cleaned and shaped root canal system. In Cohen’s Pathways of the Pulp, 10th ed.; Berman, L., Hargreaves, K., Eds.; Mosby: St. Louis, MO, USA, 2011; pp. 349–388. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Dilution of Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Approved Standard, 10th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Vankerckhoven, V.; Van-Autgaerden, T.; Vael, C.; Lammens, C.; Chapelle, S.; Rossi, R. Development of multiplex PCR for the detection of asa1, gelE, cylA, esp, and hyl genes in Enterococci and survey for virulence determinants among European hospital isolates of Enterococcus faecium. J. Clin. Microbiol. 2004, 42, 4473–4479. [Google Scholar] [CrossRef] [Green Version]
- Creti, R.; Imperi, M.; Bertuccini, L.; Fabretti, F.; Orefici, G.; Di Rosa, R.; Baldassarri, L. Survey for virulence determinants among Enterococcus faecalis isolated from different sources. J. Med. Microbiol. 2004, 53, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Adams, H. Methods of study of bacterial viruses. In Bacteriophages; Interscience Publishers: London, UK, 1959; pp. 447–448. [Google Scholar]
- Abdel-Haliem, M.E.F.; Askora, A. Isolation and characterization of bacteriophages of Helicobacter pylori isolated from Egypt. Future Virol. 2013, 8, 821–826. [Google Scholar] [CrossRef]
- Viazis, S.; Akhtar, M.; Feirtag, J.; Brabban, A.D.; Diez-Gonzalez, F. Isolation and characterization of lytic bacteriophages against enterohaemorrhagic Escherichia coli. J. App. Microbiol. 2011, 110, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005. [Google Scholar] [CrossRef] [Green Version]
- Pajunen, M.; Kiljunen, S.; Skurnik, M. Bacteriophage phiYeO3-12, specific for Yersinia enterocolitica serotype O:3, is related to coli phage T3 and T7. J. Bacteriol. 2000, 182, 5114–5120. [Google Scholar] [CrossRef] [Green Version]
- Hammerl, J.A.; Jäckel, C.; Alter, T.; Janzcyk, P.; Stingl, K.; Knüver, M.T.; Hertwig, S. Reduction of Campylobacter jejuni in broiler chicken by successive application of Group II and Group III phages. PLoS ONE 2014, 9, e114785. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A completely reimplemented MPI bioinformatics toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2011, 28, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Desper, R.; Gascuel, O. Theoretical foundation of the balanced minimum evolution method of phylogenetic inference and its relationship to weighted least-squares tree fitting. Mol Biol. Evol. 2004, 21, 587–598. [Google Scholar] [CrossRef]
- Wolanek, G.A.; Loushine, R.J.; Weller, R.N.; Kimbrough, W.F.; Volkmann, K.R. In vitro bacterial penetration of endodontically treated teeth coronally sealed with a dentin bonding agent. J. Endod. 2001, 27, 354–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.A.; Beyth, N.; Hazan, R. Targeting Enterococcus faecalis biofilms with phage therapy. Appl. Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef] [Green Version]
- Distel, J.W.; Hatton, J.F.; Gillespie, M.J. Biofilm formation in medicated root canals. J. Endod. 2002, 28, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Kafil, H.S.; Mobarez, A.M. Spread of enterococcal surface protein in antibiotic resistant Enterococcus faecium and Enterococcus faecalis isolates from urinary tract infections. Open Microbiol. 2015, 9, 14–17. [Google Scholar] [CrossRef]
- Kafil, H.S.; Mobarez, A.M.; Moghadam, M.F.; Hashemi, Z.S.; Yousefi, M. Gentamicin induces efaA expression and biofilm formation in Enterococcus faecalis. Microb. Pathog. 2016, 92, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xing, S.; Zhao, F.; Li, P.; Mi, Z.; Shi, T.; Liu, H.; Tong, Y. Characterization and genome analysis of novel phage vB_EfaP_IME195 infecting Enterococcus faecalis. Virus Genes 2018, 54, 804. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Zhang, X.; Sun, Q.; Wang, J.; Mi, Z.; Pei, G.; Huang, Y.; An, X.; Fu, K.; Zhou, L.; et al. Complete genome sequence of a novel, virulent Ahjdlikevirus bacteriophage that infects Enterococcus faecium. Arch. Virol. 2017, 162, 3843–3847. [Google Scholar] [CrossRef] [PubMed]
- Bolocan, A.S.; Upadrasta, A.; de Almeida Bettio, P.H.; Clooney, A.G.; Draper, L.A.; Ross, R.P.; Hill, C. Evaluation of phage therapy in the context of Enterococcus faecalis and its associated diseases. Viruses 2019, 11, 366. [Google Scholar] [CrossRef] [Green Version]
- Hyman, P. Phages for phage therapy: Isolation, characterization, and host range breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wei, X.; Ling, J.; Wang, W.; Huang, X. Biofilm formation capability of Enterococcus faecalis cells in starvation phase and its susceptibility to sodium hypochlorite. J. Endod. 2010, 36, 630–635. [Google Scholar] [CrossRef]
- Shen, Y.; Stojicic, S.; Haapasalo, M. Antimicrobial efficacy of chlorexhidine against bacteria in biofilms at different stages of development. J. Endod. 2011, 37, 657–661. [Google Scholar] [CrossRef]
- Pan, J.; Sun, K.; Liang, Y.; Sun, P.; Yang, X.; Wang, J.; Zhang, J.; Zhu, W.; Fang, J.; Becker, K.H. Cold plasma therapy of a tooth root canal infected with Enterococcus faecalis biofilms in vitro. J. Endod. 2013, 39, 105–110. [Google Scholar] [CrossRef]
- Zhang, W.; Mi, Z.; Yin, X.; Fan, H.; An, X.; Zhang, Z.; Chen, J.; Tong, Y. Characterization of Enterococcus faecalis phage IME-EF1 and its endolysin. PLoS ONE 2013, 8, e80435. [Google Scholar] [CrossRef]
- Lee, D.; Im, J.; Na, H.; Ryu, S.; Yun, C.-H.; Han, S.H. The novel Enterococcus phage vB_EfaS_HEf13 has broad lytic activity against clinical isolates of Enterococcus faecalis. Front. Microbiol. 2019, 10, 2877. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | β-Lactams | Aminoglycosides | Quinolones | Tetracycline | Macrolides | Streptogramins | Glycopeptide | Oxalidinone | Macrobid | MAR Index | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PEN G | AMP | GEN | STR | CIP | LVX | TET | TGC | ERY | Q-D | VAN | LZD | NIT | ||
| EF.1 | S | S | S | S | S | S | S | S | S | S | S | S | S | 0 |
| EF.2 | S | S | R | R | R | R | R | S | R | R | S | S | S | 0.54 |
| EF.3 | S | S | S | S | S | S | S | S | S | S | S | S | S | 0 |
| EF.4 | R | R | R | R | R | R | R | S | R | R | S | S | R | 0.77 |
| EF.5 | R | R | S | S | R | R | S | S | S | S | S | S | S | 0.28 |
| EF.6 | S | S | R | R | R | R | R | S | S | R | S | S | S | 0.42 |
| EF.7 | S | S | S | S | S | S | S | S | S | S | S | S | S | 0 |
| EF.8 | R | R | S | S | R | R | R | S | R | R | S | S | S | 0.5 |
| EF.9 | R | S | R | R | S | S | S | S | R | S | S | S | S | 0.28 |
| EF.10 | S | S | S | R | R | R | S | S | R | R | S | S | S | 0.35 |
| EF.11 | R | S | S | S | S | S | R | S | R | S | S | S | S | 0.21 |
| EF12 | R | S | S | R | R | R | S | S | S | S | S | S | R | 0.35 |
| EF.13 | s | S | R | S | R | R | S | S | S | R | S | S | S | 0.28 |
| EF.14 | S | S | R | S | S | S | R | S | R | R | S | S | R | 0.35 |
| EF.15 | S | S | S | S | S | S | S | S | S | S | S | S | S | 0 |
| EF.16 | R | R | R | R | S | S | S | S | R | S | S | S | S | 0.35 |
| EF.17 | R | S | R | S | R | S | S | S | S | S | S | S | S | 0.21 |
| EF.18 | S | S | R | R | S | S | R | S | R | R | S | S | S | 0.35 |
| EF.19 | R | R | S | S | R | S | S | S | S | S | S | S | S | 0.28 |
| EF.20 | R | R | S | S | R | R | R | S | S | S | S | S | R | 0.42 |
| Total R % | 10 (50%) | 6 (30%) | 9 (45%) | 8 (40%) | 11 (55%) | 9 (45%) | 8 (40%) | 0 | 9 (45%) | 8 (40%) | 0 | 0 | 4 (20%) | |
| CDS | ORF Location (nt) | Putative Function |
|---|---|---|
| 1 | 172–342 | hypothetical protein |
| 2 | 344–712 | hypothetical protein |
| 3 | 786–1136 | single-stranded DNA-binding protein |
| 4 | 1204–1383 | hypothetical protein |
| 5 | 1380–1544 | hypothetical protein |
| 6 | 1557–1721 | hypothetical protein |
| 7 | 1734–1886 | hypothetical protein |
| 8 | 1870–2292 | hypothetical protein |
| 9 | 2285–3526 | terminase large subunit |
| 10 | 3539–5887 | DNA polymerase |
| 11 | 5947–6117 | hypothetical protein |
| 12 | 6117–6323 | hypothetical protein |
| 13 | 6314–6487 | hypothetical protein |
| 14 | 6488–6727 | transcriptional regulator |
| 15 | 6724–7221 | HNH homing endonuclease |
| 16 | 7221–7406 | putative transmembrane protein |
| 17 | 7406–8824 | CHAP domain-containing lysin |
| 18 | 8943–9389 | DNA binding protein |
| 19 | 9406–10182c | peptidoglycan recognition and hydrolase |
| 20 | 10183–10575c | hypothetical protein |
| 21 | 10575–12329c | tail fiber protein |
| 22 | 12331–13119c | distal tail protein |
| 23 | 13131–14696c | phosphodiesterase |
| 24 | 14710–15246c | hypothetical protein |
| 25 | 15296–16342c | phage capsid and scaffold |
| 26 | 16358–17575c | phage capsid and scaffold |
| 27 | 17577–17747c | capsid related protein |
| 28 | 17842–18177c | hypothetical protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Telbany, M.; El-Didamony, G.; Askora, A.; Ariny, E.; Abdallah, D.; Connerton, I.F.; El-Shibiny, A. Bacteriophages to Control Multi-Drug Resistant Enterococcus faecalis Infection of Dental Root Canals. Microorganisms 2021, 9, 517. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030517
El-Telbany M, El-Didamony G, Askora A, Ariny E, Abdallah D, Connerton IF, El-Shibiny A. Bacteriophages to Control Multi-Drug Resistant Enterococcus faecalis Infection of Dental Root Canals. Microorganisms. 2021; 9(3):517. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030517
Chicago/Turabian StyleEl-Telbany, Mohamed, Gamal El-Didamony, Ahmed Askora, Eman Ariny, Dalia Abdallah, Ian F. Connerton, and Ayman El-Shibiny. 2021. "Bacteriophages to Control Multi-Drug Resistant Enterococcus faecalis Infection of Dental Root Canals" Microorganisms 9, no. 3: 517. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030517