
An Update on Wastewater Multi-Resistant Bacteria: Identification of Clinical Pathogens Such as Escherichia coli O25b:H4-B2-ST131-Producing CTX-M-15 ESBL and KPC-3 Carbapenemase-Producing Klebsiella oxytoca


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotypic Characterization of Isolates from Wastewater
2.1.1. Sample Sites and Collection
2.1.2. Microbiological Analyses
2.1.3. Isolation of Resistant Gram-Negative Bacteria
2.1.4. Antibiotic Susceptibility Testing
2.2. Genomic Characterization of Isolates from Wastewater
2.2.1. DNA Extraction and Sequencing
2.2.2. Genotype Identification of Isolates
2.2.3. Detection of Antibiotic Resistance Genes
2.2.4. Phylogenetic Group, Molecular Typing, and Sequence Analysis
2.2.5. Whole-Genome Sequencing and Sequence Analysis
3. Results
3.1. Phenotypic Characterization
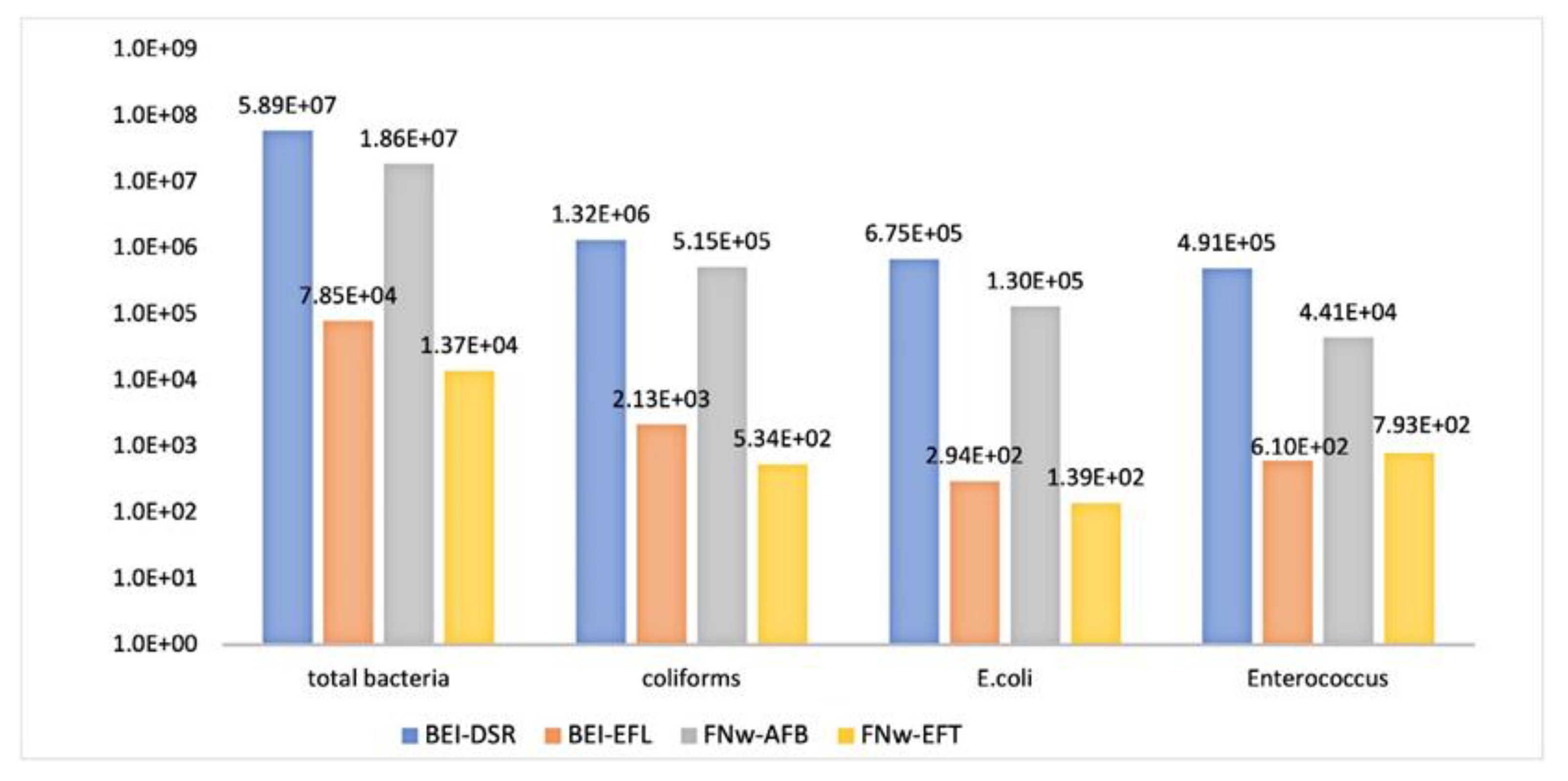
3.1.1. Enumeration of Wastewater Isolates
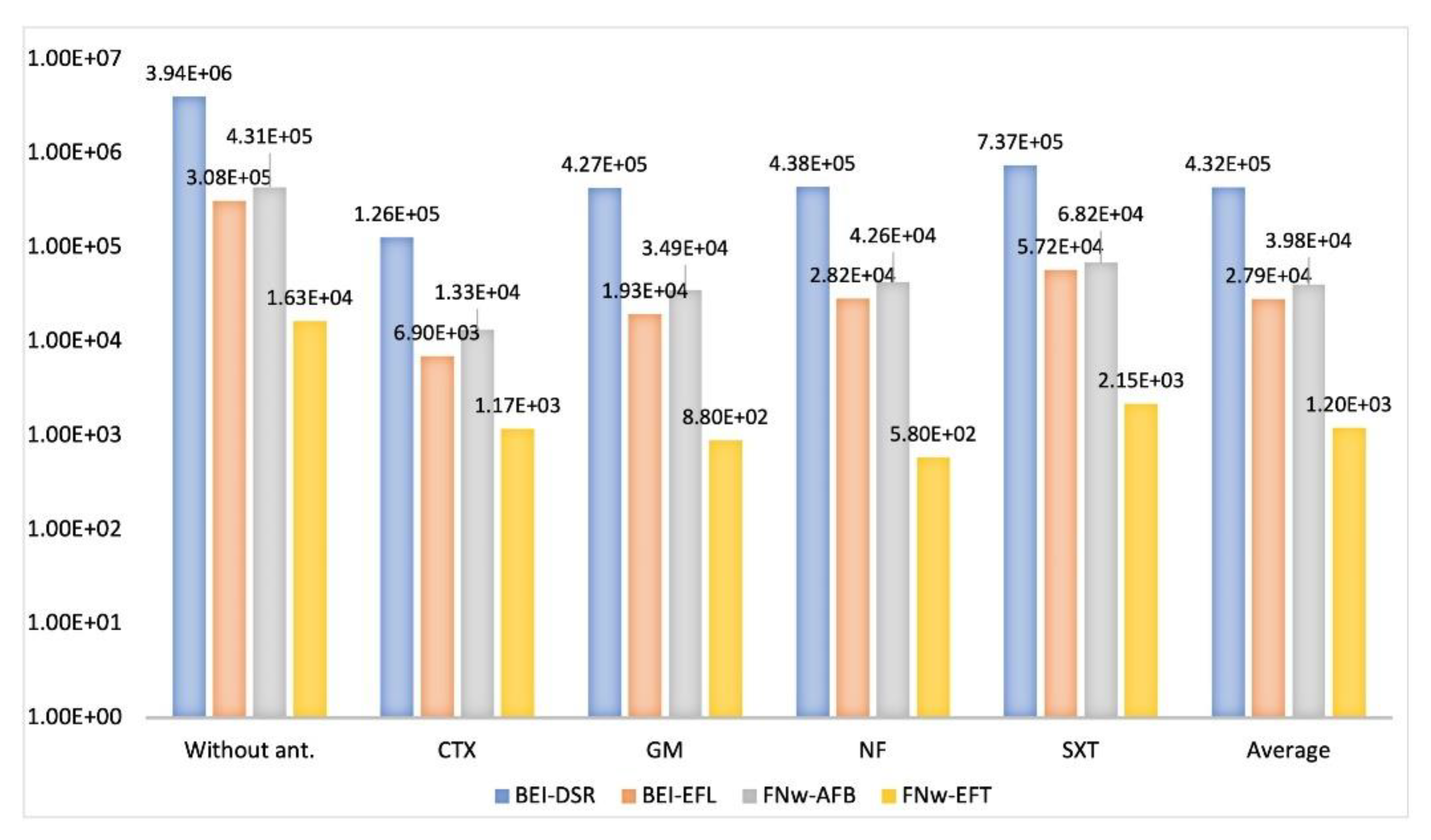
3.1.2. Enumeration of Antimicrobial-Resistant Gram-Negative Bacteria
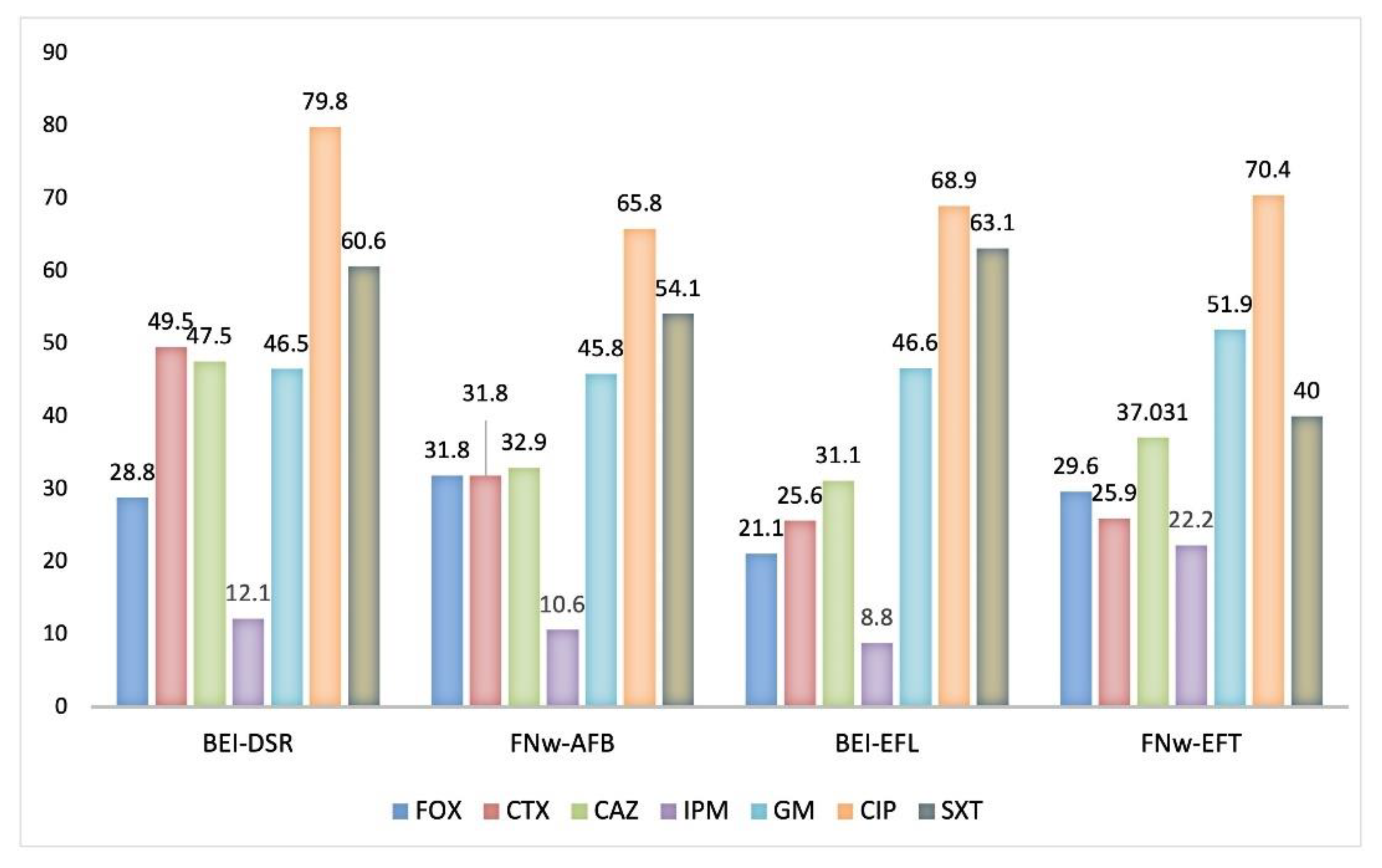
3.1.3. Antimicrobial Susceptibility Profiles
3.2. Genomic Characterization
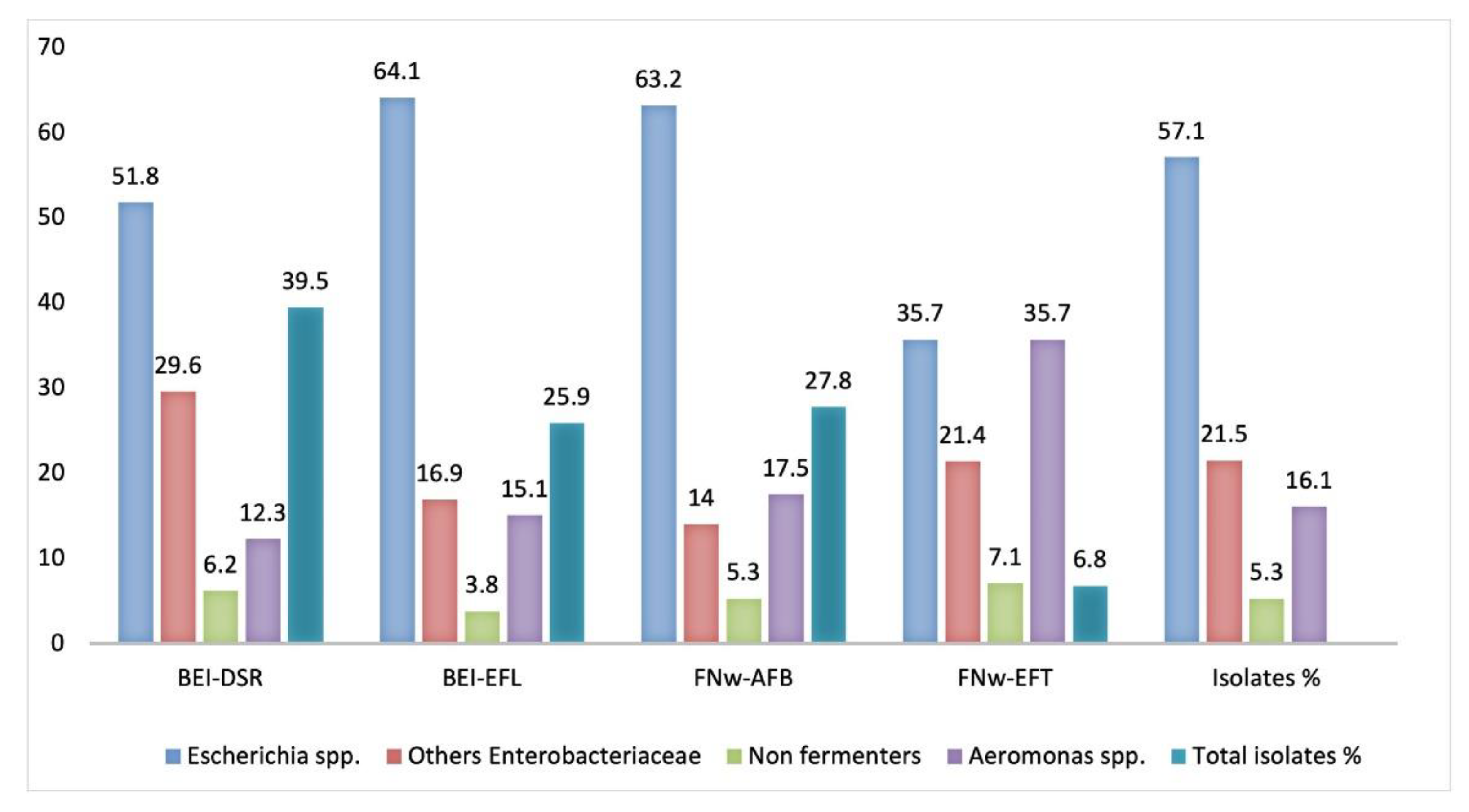
3.2.1. Identification of Isolates by 16S rRNA
3.2.2. Detection and Sequencing of Antibiotic Resistance Genes
3.2.3. Molecular Typing Characterization
3.2.4. Whole-Genome Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanderson, H.; Fricker, C.; Brown, R.S.; Majury, A.; Liss, S.N. Antibiotic resistance genes as an emerging environmental contaminant. Environ. Rev. 2016, 24, 205–218. [Google Scholar] [CrossRef]
- Caneiras, C.; Lito, L.; Mayoralas-Alises, S.; Diaz-Lobato, S.; Melo-Cristino, J.; Duarte, A. Virulence and resistance determinants of Klebsiella pneumoniae isolated from a Portuguese tertiary university hospital centre over a 31-year period. Enferm. Infecc. Microbiol. Clin. 2019, 37, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Perdigao, J.; Caneiras, C.; Elias, R.; Modesto, A.; Spadar, A.; Phelan, J.; Campino, S.; Clark, T.G.; Costa, E.; Saavedra, M.J.; et al. Genomic Epidemiology of Carbapenemase Producing Klebsiella pneumoniae Strains at a Northern Portuguese Hospital Enables the Detection of a Misidentified Klebsiella variicola KPC-3 Producing Strain. Microorganisms 2020, 8, 1986. [Google Scholar] [CrossRef] [PubMed]
- Manaia, C.M.; Rocha, J.; Scaccia, N.; Marano, R.; Radu, E.; Biancullo, F.; Cerqueira, F.; Fortunato, G.; Iakovides, I.C.; Zammit, I.; et al. Antibiotic resistance in wastewater treatment plants: Tackling the black box. Environ. Int. 2018, 115, 312–324. [Google Scholar] [CrossRef]
- Karkman, A.; Do, T.T.; Walsh, F.; Virta, M.P.J. Antibiotic-Resistance Genes in Waste Water. Trends Microbiol. 2018, 26, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tehrani, A.H.; Gilbride, K.A. A closer look at the antibiotic-resistant bacterial community found in urban wastewater treatment systems. Microbiologyopen 2018, 7, e00589. [Google Scholar] [CrossRef]
- Bouki, C.; Venieri, D.; Diamadopoulos, E. Detection and fate of antibiotic resistant bacteria in wastewater treatment plants: A review. Ecotoxicol. Environ. Saf. 2013, 91, 1–9. [Google Scholar] [CrossRef]
- Gilchrist, C.A.; Turner, S.D.; Riley, M.F.; Petri, W.A., Jr.; Hewlett, E.L. Whole-genome sequencing in outbreak analysis. Clin. Microbiol. Rev. 2015, 28, 541–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO. Water quality—Evaluation of membrane filters used for microbiological analyses. In International Standard ISO 7704; International Organization for Standardization: Geneva, Switzerland, 1985. [Google Scholar]
- ISO. Water Quality—General requirements and guidance for microbiological examinations by culture. In International Standard ISO 8199; International Organization for Standardization: Geneva, Switzerland, 1988. [Google Scholar]
- ISO. Water Quality-Enumeration of Escherichia coli and Coliform Bacteria—Part 1: Membrane Filtration Method for Waters with Low Bacterial Background Flora. In International Standard ISO 9308-1; International Organization for Standardization: Geneva, Switzerland, 2014. [Google Scholar]
- Water Research Australia Water for the wellbeing of all Australians. Log Removal Values in Wastewater Treatment. Factsheets April 2014. Available online: https://www.waterra.com.au/publications/factsheets/ (accessed on 1 February 2021).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Conceicao, T.; Brizio, A.; Duarte, A.; Lito, L.M.; Cristino, J.M.; Salgado, M.J. First description of CTX-M-15-producing Klebsiella pneumoniae in Portugal. Antimicrob. Agents Chemother. 2005, 49, 477–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naas, T.; Cuzon, G.; Villegas, M.V.; Lartigue, M.F.; Quinn, J.P.; Nordmann, P. Genetic structures at the origin of acquisition of the beta-lactamase blaKPC gene. Antimicrob. Agents Chemother. 2008, 52, 1257–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, G.; Korem, M.; Robicsek, A.; Engelstein, D.; Moses, A.E.; Block, C.; Strahilevitz, J. Changes in aac(6’)-Ib-cr prevalence and fluoroquinolone resistance in nosocomial isolates of Escherichia coli collected from 1991 through 2005. Antimicrob. Agents Chemother. 2009, 53, 1268–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Dhanji, H.; Upton, M.; Gibreel, T.; Fox, A.; Boyd, D.; Mulvey, M.R.; Nordmann, P.; Ruppe, E.; Sarthou, J.L.; et al. Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 2009, 64, 274–277. [Google Scholar] [CrossRef] [Green Version]
- Compeau, P.E.; Pevzner, P.A.; Tesler, G. How to apply de Bruijn graphs to genome assembly. Nat. Biotechnol. 2011, 29, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic. Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joensen, K.G.; Tetzschner, A.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and Easy In Silico Serotyping of Escherichia coli Isolates by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Leung, P.C.; Qian, P.; JI-Dong, G.U. Effects of UV, H2O2 and Fe3C on the Growth of Four Environmental Isolates of Aeromonas and Vibrio Species from a Mangrove Environment. Microbes Environ. 2004, 19, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Caneiras, C.; Calisto, F.; Jorge da Silva, G.; Lito, L.; Melo-Cristino, J.; Duarte, A. First Description of Colistin and Tigecycline-Resistant Acinetobacter baumannii Producing KPC-3 Carbapenemase in Portugal. Antibiotics 2018, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Adriana Osińska, E.K.; Monika, H.; Sebastian, N. Impact of type of wastewater treatment process on the antibiotic resistance of bacterial populations. E3S Web Conf. 2017, 17. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Report on Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: https://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 1 February 2021).
- Janda, J.M.; Abbott, S.L. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [Green Version]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging species infections and their significance in public health. Sci. World J. 2012, 2012, 625023. [Google Scholar] [CrossRef] [Green Version]
- Talagrand-Reboul, E.; Roger, F.; Kimper, J.L.; Colston, S.M.; Graf, J.; Latif-Eugenin, F.; Figueras, M.J.; Petit, F.; Marchandin, H.; Jumas-Bilak, E.; et al. Delineation of Taxonomic Species within Complex of Species: Aeromonas media and Related Species as a Test Case. Front. Microbiol. 2017, 8, 621. [Google Scholar] [CrossRef]
- Chen, P.L.; Lamy, B.; Ko, W.C. Aeromonas dhakensis, an Increasingly Recognized Human Pathogen. Front. Microbiol. 2016, 7, 793. [Google Scholar] [CrossRef]
- Carr, E.L.; Kampfer, P.; Patel, B.K.C.; Gurtler, V.; Seviour, R.J. Seven novel species of Acinetobacter isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 2003, 53, 953–963. [Google Scholar] [CrossRef] [Green Version]
- Al Atrouni, A.; Joly-Guillou, M.L.; Hamze, M.; Kempf, M. Reservoirs of Non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Karah, N.; Haldorsen, B.; Hegstad, K.; Simonsen, G.S.; Sundsfjord, A.; Samuelsen, O.; Norwegian Study Group of Acinetobacter. Species identification and molecular characterization of Acinetobacter spp. blood culture isolates from Norway. J. Antimicrob. Chemother. 2011, 66, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, M.A.; Ozer, E.; Bolon, M.K.; Hauser, A.R. Influence of ACB complex genospecies on clinical outcomes in a U.S. hospital with high rates of multidrug resistance. J. Infect. 2015, 70, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Chen, Y.; Jia, X.; Luo, Y.; Song, Q.; Zhao, W.; Wang, Y.; Liu, H.; Zheng, D.; Xia, Y.; et al. Dissemination and characterization of NDM-1-producing Acinetobacter pittii in an intensive care unit in China. Clin. Microbiol. Infect. 2012, 18, E506–E513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleicher, X.; Higgins, P.G.; Wisplinghoff, H.; Korber-Irrgang, B.; Kresken, M.; Seifert, H. Molecular epidemiology of Acinetobacter baumannii and Acinetobacter nosocomialis in Germany over a 5-year period (2005–2009). Clin. Microbiol. Infect. 2013, 19, 737–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsini, J.; Tam, E.; Hauser, N.; Rajayer, S. Polymicrobial Bacteremia Involving Comamonas testosteroni. Case Rep. Med. 2014, 2014, 578127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vela, A.I.; Collins, M.D.; Lawson, P.A.; Garcia, N.; Dominguez, L.; Fernandez-Garayzabal, J.F. Uruburuella suis gen. nov., sp. nov., isolated from clinical specimens of pigs. Int. J. Syst. Evol. Microbiol. 2005, 55, 643–647. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.P.; Pembroke, J.T. The Genus Ochrobactrum as Major Opportunistic Pathogens. Microorganisms 2020, 8, 1797. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, T.; Sharma, D.; Sharma, H.; Kandil, H.; Collier, S. Ochromobactrum intermedium: An emerging opportunistic pathogen-case of recurrent bacteraemia associated with infective endocarditis in a haemodialysis patient. New Microbes New Infect. 2017, 15, 14–15. [Google Scholar] [CrossRef] [Green Version]
- Petrella, S.; Clermont, D.; Casin, I.; Jarlier, V.; Sougakoff, W. Novel class A beta-lactamase Sed-1 from Citrobacter sedlakii: Genetic diversity of beta-lactamases within the Citrobacter genus. Antimicrob. Agents Chemother. 2001, 45, 2287–2298. [Google Scholar] [CrossRef] [Green Version]
- Caneiras, C.; Lito, L.; Melo-Cristino, J.; Duarte, A. Community- and Hospital-Acquired Klebsiella pneumoniae Urinary Tract Infections in Portugal: Virulence and Antibiotic Resistance. Microorganisms 2019, 7, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Russo, T.A. Extraintestinal pathogenic Escherichia coli: “the other bad E coli”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Reppin, F.C.; Cochet, S.; El Nemer, W.; Fritz, G.; Schmidt, G. High Affinity Binding of Escherichia coli Cytotoxic Necrotizing Factor 1 (CNF1) to Lu/BCAM Adhesion Glycoprotein. Toxins 2018, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Mao, B.H.; Chang, Y.F.; Scaria, J.; Chang, C.C.; Chou, L.W.; Tien, N.; Wu, J.J.; Tseng, C.C.; Wang, M.C.; Chang, C.C.; et al. Identification of Escherichia coli genes associated with urinary tract infections. J. Clin. Microbiol. 2012, 50, 449–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Cadena, E.; Mojica, M.F.; Castillo, N.; Correa, A.; Appel, T.M.; Garcia-Betancur, J.C.; Pallares, C.J.; Villegas, M.V. Genomic Analysis of CTX-M-Group-1-Producing Extraintestinal Pathogenic E. coli (ExPEc) from Patients with Urinary Tract Infections (UTI) from Colombia. Antibiotics 2020, 9, 899. [Google Scholar] [CrossRef] [PubMed]
- Glaubman, J.; Hofmann, J.; Bonney, M.E.; Park, S.; Thomas, J.M.; Kokona, B.; Ramos Falcon, L.I.; Chung, Y.K.; Fairman, R.; Okeke, I.N. Self-association motifs in the enteroaggregative Escherichia coli heat-resistant agglutinin 1. Microbiology 2016, 162, 1091–1102. [Google Scholar] [CrossRef]
- Jakes, K.S.; Finkelstein, A. The colicin Ia receptor, Cir, is also the translocator for colicin Ia. Mol. Microbiol. 2010, 75, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Nipic, D.; Podlesek, Z.; Budic, M.; Crnigoj, M.; Zgur-Bertok, D. Escherichia coli uropathogenic-specific protein, Usp, is a bacteriocin-like genotoxin. J. Infect. Dis 2013, 208, 1545–1552. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Grobbel, M.; Stamm, I.; Kopp, P.A.; Diehl, I.; Semmler, T.; Fruth, A.; Beutlich, J.; Guerra, B.; Wieler, L.H.; et al. Emergence of human pandemic O25:H4-ST131 CTX-M-15 extended-spectrum-beta-lactamase-producing Escherichia coli among companion animals. J. Antimicrob. Chemother. 2010, 65, 651–660. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Product | Antibiotic Resistance | BEI- DSR | BEI- EFL | FNw- AFB | FNw- EFT | Total Number of Resistance Genes |
|---|---|---|---|---|---|---|---|
| aacA4 | Aminoglycoside 6’-N-acetyltransferase | kanamycin | 1 | 1 | |||
| aadA1 | Streptomycin 3’-adenylyltransferase | streptomycin | 5 | 3 | 1 | 9 | |
| aadA2 | Streptomycin 3’-adenylyltransferase | streptomycin | 6 | 2 | 3 | 11 | |
| aadA5 | Streptomycin 3’-adenylyltransferase | streptomycin | 9 | 7 | 3 | 3 | 22 |
| qnrB-1 | DNA-gyrase and topoisomerase | ciprofloxacin | 2 | 1 | 3 | ||
| qnrB4 | DNA-gyrase and topoisomerase | ciprofloxacin | 1 | 1 | 3 | ||
| qnrB13 | DNA-gyrase and topoisomerase | ciprofloxacin | 1 | 1 | 2 | ||
| aac(6’)-Ib-cr5 | Aminoglycoside 6’-N-acetyltransferase | ciprofloxacin and kanamycin | 19 | 4 | 2 | 1 | 26 |
| dfrA1 | Dihydrofolate reductase | trimethoprim | 3 | 6 | 1 | 10 | |
| dfrA7 | Dihydrofolate reductase | trimethoprim | 3 | 3 | |||
| dfrA12 | Dihydrofolate reductase | trimethoprim | 6 | 2 | 2 | 10 | |
| dfrA15 | Dihydrofolate reductase | trimethoprim | 2 | 2 | |||
| dfrA17 | Dihydrofolate reductase | trimethoprim | 9 | 6 | 4 | 2 | 21 |
| dfrA27 | Dihydrofolate reductase | trimethoprim | 1 | 1 | |||
| arr-3 | Rifampicin ADP-ribosyltransferase | rifamycin | 1 | 1 | 2 | ||
| CTX-M-1 | Class A β-lactamase | cephalosporins | 1 | 1 | |||
| CTX-M-2 | Class A β-lactamase | cephalosporins | 1 | 1 | |||
| CTX-M-14 | Class A β-lactamase | cephalosporins | 2 | 4 | 2 | 8 | |
| CTX-M-15 | Class A β-lactamase | cephalosporins | 22 | 6 | 6 | 3 | 37 |
| CTX-M-27 | Class A β-lactamase | cephalosporins | 1 | 2 | 1 | 4 | |
| CTX-M-32 | Class A β-lactamase | cephalosporins | 1 | 1 | |||
| CTX-M-174 | Class A β-lactamase | cephalosporins | 1 | 1 | |||
| OXY-2 | Class A β-lactamase | cephalosporins | 1 | 1 | |||
| SED-1 | Class A β-lactamase | cephalosporins | 1 | 1 | |||
| KPC-3 | Class A β-lactamase | carbapenem | 1 | 1 | |||
| OXA-2 | Class A β-lactamase | oxacilin | 1 | 1 |
| Isolates | Origin | ESBL | ST | Serotype | Phylogroup |
|---|---|---|---|---|---|
| 5301 | BEI-EFL_10TM | CTX-M-15 | 131 | O25b | B2 |
| 5302 | BEI-EFL_12CTX | CTX-M-15 | 131 | O25b | B2 |
| 5262 | BEI-DSR_6TM | CTX-M-174 | 131 | O25b | B2 |
| 5318 | FNw-EFT_11GM | CTX-M-15 | 44 | O25b | A |
| 5281 | FNw-AFB_7GM | CTX-M-15 | 410 | O25b | A |
| 5064 | FNw-AFB_10CTX | CTX-M-15 | 10 | O25b | A |
| 5295 | BEI-EFL_9TM | CTX-M-14 | 23 | O25b | A |
| Isolates | Resistance Genes | Plasmids | Virulence Genes | |||
|---|---|---|---|---|---|---|
| β-Lactamases | Others Genes | Replicon | Offensive Factors | Iron Uptake | Fimbrial Types | |
| 5301 | CTX-M-15; OXA-1; EC5 | dfrA17; aadA5; qacEdelta1; sul1; aac(3)-IIa; catB3; aac(6’)-Ib-cr5; | FIA; FIB; FII; I1-I | CNF1; Iss; Hra1; Colicin Ia; Usp; senB | Enterobactin; Aerobactin; Yersinobactin; ChuA; sitA | CFA/1 (clade α); Type1(clade γ1); Yad (clade γ4); Yeh (clade γ4); Ybg (clade π); Yfc (clade π); P (clade π); Type IV |
| 5281 | CTX-M-15; TEM-1 | dfrA1; sat2; aadA1 | HI1A; HI1B; R | Enterobactin; Aerobactin | CFA/1 (clade α); Type1(clade γ1); Sfm (clade γ1); Type IV | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesquita, E.; Ribeiro, R.; Silva, C.J.C.; Alves, R.; Baptista, R.; Condinho, S.; Rosa, M.J.; Perdigão, J.; Caneiras, C.; Duarte, A. An Update on Wastewater Multi-Resistant Bacteria: Identification of Clinical Pathogens Such as Escherichia coli O25b:H4-B2-ST131-Producing CTX-M-15 ESBL and KPC-3 Carbapenemase-Producing Klebsiella oxytoca. Microorganisms 2021, 9, 576. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030576
Mesquita E, Ribeiro R, Silva CJC, Alves R, Baptista R, Condinho S, Rosa MJ, Perdigão J, Caneiras C, Duarte A. An Update on Wastewater Multi-Resistant Bacteria: Identification of Clinical Pathogens Such as Escherichia coli O25b:H4-B2-ST131-Producing CTX-M-15 ESBL and KPC-3 Carbapenemase-Producing Klebsiella oxytoca. Microorganisms. 2021; 9(3):576. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030576
Chicago/Turabian StyleMesquita, Elsa, Rita Ribeiro, Carla J. C. Silva, Rita Alves, Rita Baptista, Sílvia Condinho, Maria João Rosa, João Perdigão, Cátia Caneiras, and Aida Duarte. 2021. "An Update on Wastewater Multi-Resistant Bacteria: Identification of Clinical Pathogens Such as Escherichia coli O25b:H4-B2-ST131-Producing CTX-M-15 ESBL and KPC-3 Carbapenemase-Producing Klebsiella oxytoca" Microorganisms 9, no. 3: 576. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030576