
Relative Contribution of Each Component of the French Ante-Mortem Surveillance System for Bovine Tuberculosis in Its Overall Sensitivity



Abstract
:1. Introduction
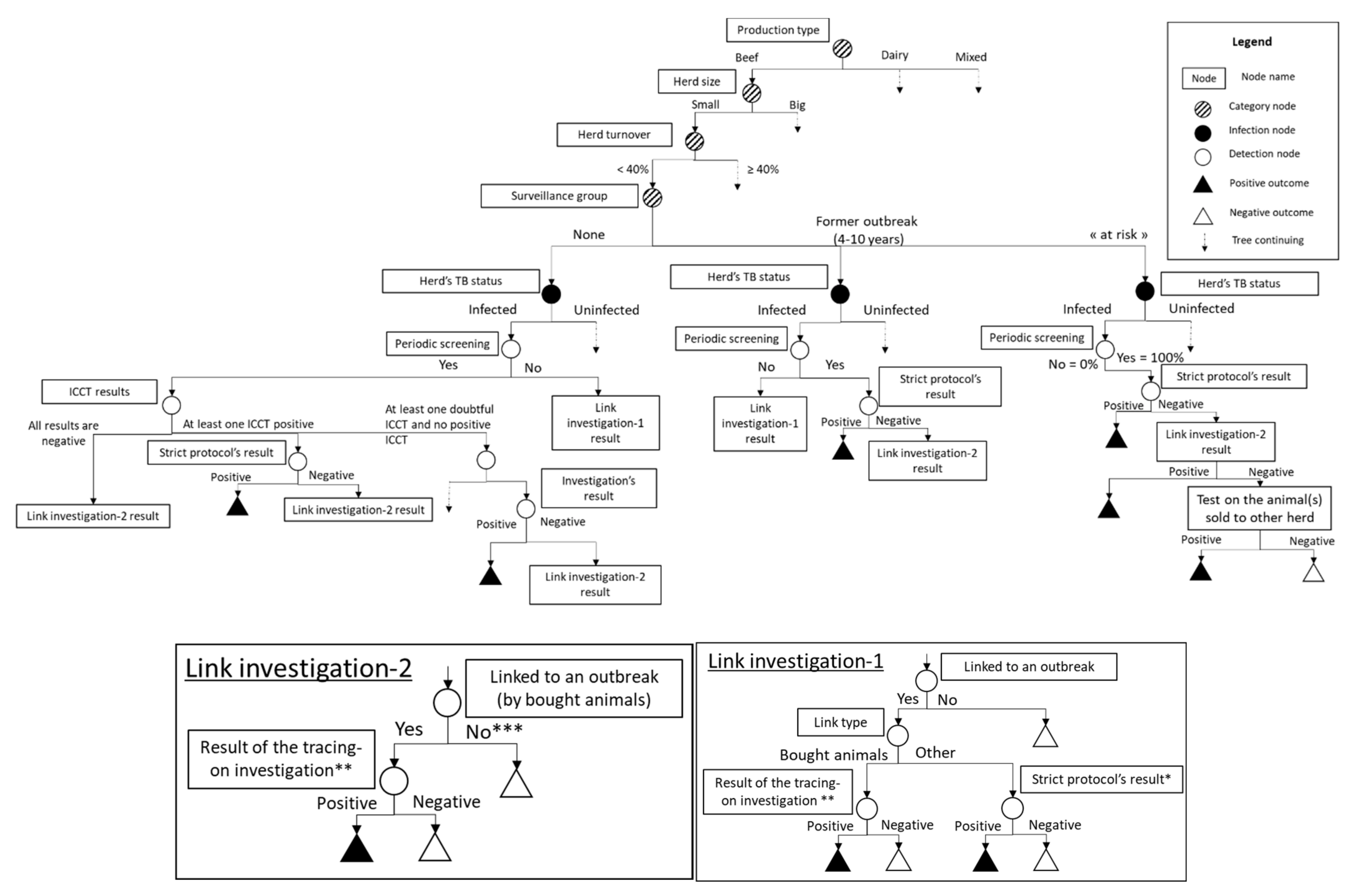
2. Materials and Methods
2.1. Category Nodes
2.2. Likelihood for a Herd to Be Classified “at Risk”
2.3. Probability for the Herd to Be a Former Outbreak Older Than 3 Years
2.4. Adjusted Relative Risks of Infection
2.5. Probability of Infection
2.6. Periodic Screening on Herds
2.6.1. Likelihood of a Herd Being Subject to Periodic Screening
2.6.2. Periodic Screening Protocol Used
2.6.3. Sensitivity and Specificity of Protocols
2.7. Epidemiological Investigations
2.7.1. Likelihood of a Herd Having a Downstream Link to a TB Outbreak
2.7.2. Investigation Protocol Used for the Downstream Link
2.7.3. Likelihood of a Herd Having an Upstream Link to a TB Outbreak
2.8. Screening of Exchanged Animals
2.9. Calculations: Likelihood of Detecting Infection in Areas
3. Results
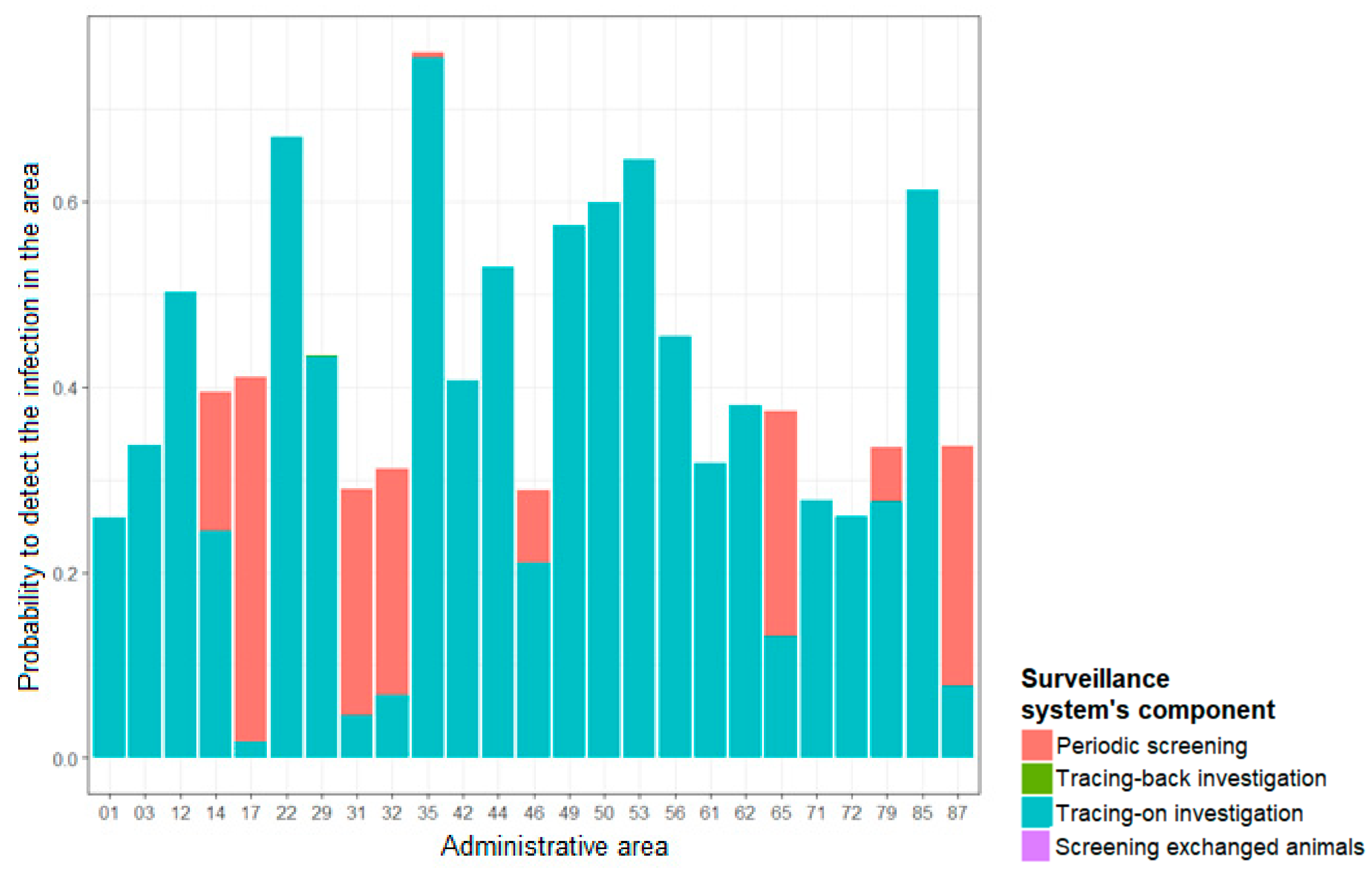
3.1. Effectiveness of the TB Ante-Mortem Surveillance System
3.2. Contribution of Each Component in the Overall Sensitivity of the Ante-Mortem Surveillance System
4. Discussion
4.1. Material and Method
4.2. Results
4.2.1. Effectiveness of the Ante-Mortem Surveillance System
4.2.2. Contribution of Each Surveillance System’s Component
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area’s Identification Number | Area’s Name | Number of Outbreaks between 2011 and 2016 | Prevalence Used for Modeling |
|---|---|---|---|
| 01 | Ain | [0–5[ | 0.0001 |
| 02 | Aisne | [0–5[ | 0.0001 |
| 03 | Allier | [0–5[ | 0.0001 |
| 04 | Alpes-de-Haute-Provence | [0–5[ | 0.0001 |
| 05 | Hautes-Alpes | [0–5[ | 0.0001 |
| 06 | Alpes-Maritimes | [0–5[ | 0.0001 |
| 07 | Ardèche | [0–5[ | 0.0001 |
| 08 | Ardennes | 10 et plus | 0.006 |
| 09 | Ariège | [5–10[ | 0.0002 |
| 10 | Aube | [0–5[ | 0.0001 |
| 11 | Aude | [0–5[ | 0.0001 |
| 12 | Aveyron | [0–5[ | 0.0001 |
| 13 | Bouches-du-Rhône | [0–5[ | 0.0001 |
| 14 | Calvados | [0–5[ | 0.0001 |
| 15 | Cantal | [0–5[ | 0.0001 |
| 16 | Charente | 10 et plus | 0.006 |
| 17 | Charente-Maritime | [5–10[ | 0.0002 |
| 18 | Cher | [0–5[ | 0.0001 |
| 19 | Corrèze | [0–5[ | 0.0001 |
| 2A | Corse-du-Sud | [0–5[ | 0.0001 |
| 2B | Haute-Corse | 10 et plus | 0.006 |
| 21 | Côte-d’Or | 10 et plus | 0.006 |
| 22 | Cotes-d’Armor | [0–5[ | 0.0001 |
| 23 | Creuse | [0–5[ | 0.0001 |
| 24 | Dordogne | 10 et plus | 0.006 |
| 25 | Doubs | [0–5[ | 0.0001 |
| 26 | Drome | [0–5[ | 0.0001 |
| 27 | Eure | [0–5[ | 0.0001 |
| 28 | Eure-et-Loir | [0–5[ | 0.0001 |
| 29 | Finistère | [0–5[ | 0.0001 |
| 30 | Gard | [0–5[ | 0.0001 |
| 31 | Haute-Garonne | [0–5[ | 0.0001 |
| 32 | Gers | [5–10[ | 0.0002 |
| 33 | Gironde | [0–5[ | 0.0001 |
| 34 | Hérault | [5–10[ | 0.0002 |
| 35 | Ille-et-Vilaine | [0–5[ | 0.0001 |
| 36 | Indre | [0–5[ | 0.0001 |
| 37 | Indre-et-Loire | [0–5[ | 0.0001 |
| 38 | Isère | [0–5[ | 0.0001 |
| 39 | Jura | [0–5[ | 0.0001 |
| 40 | Landes | 10 et plus | 0.006 |
| 41 | Loir-et-Cher | [0–5[ | 0.0001 |
| 42 | Loire | [5–10[ | 0.0002 |
| 43 | Haute-Loire | [0–5[ | 0.0001 |
| 44 | Loire-Atlantique | [0–5[ | 0.0001 |
| 45 | Loiret | [0–5[ | 0.0001 |
| 46 | Lot | [0–5[ | 0.0001 |
| 47 | Lot-et-Garonne | 10 et plus | 0.006 |
| 48 | Lozère | [0–5[ | 0.0001 |
| 49 | Maine-et-Loire | [0–5[ | 0.0001 |
| 50 | Manche | [5–10[ | 0.0002 |
| 51 | Marne | [0–5[ | 0.0001 |
| 52 | Haute-Marne | [0–5[ | 0.0001 |
| 53 | Mayenne | [5–10[ | 0.0002 |
| 54 | Meurthe-et-Moselle | [0–5[ | 0.0001 |
| 55 | Meuse | [0–5[ | 0.0001 |
| 56 | Morbihan | [0–5[ | 0.0001 |
| 57 | Moselle | [0–5[ | 0.0001 |
| 58 | Nièvre | [0–5[ | 0.0001 |
| 59 | Nord | [0–5[ | 0.0001 |
| 60 | Oise | [0–5[ | 0.0001 |
| 61 | Orne | [0–5[ | 0.0001 |
| 62 | Pas-de-Calais | [0–5[ | 0.0001 |
| 63 | Puy-de-Dôme | [0–5[ | 0.0001 |
| 64 | Pyrénées-Atlantiques | 10 et plus | 0.006 |
| 65 | Hautes-Pyrénées | [0–5[ | 0.0001 |
| 66 | Pyrénées-Orientales | [0–5[ | 0.0001 |
| 67 | Bas-Rhin | [0–5[ | 0.0001 |
| 68 | Haut-Rhin | [0–5[ | 0.0001 |
| 69 | Rhône + métropole de Lyon | [0–5[ | 0.0001 |
| 70 | Haute-Saône | [0–5[ | 0.0001 |
| 71 | Saône-et-Loire | [0–5[ | 0.0001 |
| 72 | Sarthe | [0–5[ | 0.0001 |
| 73 | Savoie | [0–5[ | 0.0001 |
| 74 | Haute-Savoie | [0–5[ | 0.0001 |
| 75 | Paris | [0–5[ | 0.0001 |
| 76 | Seine-Maritime | [5–10[ | 0.0002 |
| 77 | Seine-et-Marne | [0–5[ | 0.0001 |
| 78 | Yvelines | [0–5[ | 0.0001 |
| 79 | Deux-Sèvres | [5–10[ | 0.0002 |
| 80 | Somme | [0–5[ | 0.0001 |
| 81 | Tarn | [0–5[ | 0.0001 |
| 82 | Tarn-et-Garonne | [0–5[ | 0.0001 |
| 83 | Var | [0–5[ | 0.0001 |
| 84 | Vaucluse | [0–5[ | 0.0001 |
| 85 | Vendée | [0–5[ | 0.0001 |
| 86 | Vienne | [0–5[ | 0.0001 |
| 87 | Haute-Vienne | [5–10[ | 0.0002 |
| 88 | Vosges | [0–5[ | 0.0001 |
| 89 | Yonne | [0–5[ | 0.0001 |
| 90 | Territoire de Belfort | [0–5[ | 0.0001 |
| 91 | Essonne | [0–5[ | 0.0001 |
| 92 | Hauts-de-Seine | [0–5[ | 0.0001 |
| 93 | Seine-Saint-Denis | [0–5[ | 0.0001 |
| 94 | Val-de-Marne | [0–5[ | 0.0001 |
| 95 | Val-d’Oise | [0–5[ | 0.0001 |
Appendix A.1. Calculations A1. Deduction of a Modeling of the Raw Sensitivity and Specificity of the ICCT from French Data
Appendix A.2. Calculations A2. Method for Estimating the Probabilities of Obtaining a Positive or Negative Final Result in a Herd for Each of the Regulatory Protocols for Periodic Screening, Conditional on the Results of the ICCT1
- Results of the Investigation Using the “Strict” Protocol Conditional on Obtaining at Least One Positive ICCT1 in the Herd
- Results of the “compliant slow path” and “compliant quick path” protocols conditional on obtaining at least one non-negative ICCT1 but no positive ICCT1





References
- Robert, J.; Boulahbal, F.; Trystram, D.; Truffot-Pernot, C.; De Benoist, A.C.; Vincent, V.; Jarlier, V.; Grosset, J. A national survey of human Mycobacterium bovis infection in France. Network of Microbiology Laboratories in France. Int. J. Tuberc. Lung Dis. 1999, 3, 711–714. [Google Scholar] [PubMed]
- De la Rua-Domenech, R. Human Mycobacterium bovis infection in the United Kingdom: Incidence, risks, control measures and review of the zoonotic aspects of bovine tuberculosis. Tuberculosis 2006, 86, 77–109. [Google Scholar] [CrossRef] [PubMed]
- Thoen, C.; LoBue, P.; de Kantor, I. The importance of Mycobacterium bovis as a zoonosis. Vet. Microbiol. 2006, 112, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Poirier, V.; Rivière, J.; Bouveret, A.; Gardon, S.; Dufour, B. Cost-effectiveness assessment of three components of the bovine tuberculosis surveillance system by intradermal tuberculin testing in French cattle farms by a scenario tree approach. Prev. Vet. Med. 2019, 166, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Guétin-Poirier, V.; Rivière, J.; Dufour, B. Cost-effectiveness of two different protocols for animal tracing investigations of bovine tuberculosis outbreaks in France. Prev. Vet. Med. 2020, 175, 104868. [Google Scholar] [CrossRef] [PubMed]
- Guétin-Poirier, V.; Crozet, G.; Gardon, S.; Dufour, B.; Rivière, J. Integrating data of veterinarians’ practices in assessing the cost effectiveness of three components of the bovine tuberculosis surveillance system by intradermal tuberculin testing in French cattle farms through a scenario-tree approach. Res. Vet. Sci. 2020, 128, 242–260. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.M.; Martin, S.; Thorburn, M.A.; Eves, J.A.; Hammond, R.F. A case-control study on the association of selected risk factors with the occurrence of bovine tuberculosis in the Republic of Ireland. Prev. Vet. Med. 1996, 27, 75–87. [Google Scholar] [CrossRef]
- Munroe, F.A.; Dohoo, I.R.; McNab, W.B.; Spangler, L. Risk factors for the between-herd spread of Mycobacterium bovis in Canadian cattle and cervids between 1985 and 1994. Prev. Vet. Med. 1999, 41, 119–133. [Google Scholar] [CrossRef]
- Reilly, L.; Courtenay, O. Husbandry practices, badger sett density and habitat composition as risk factors for transient and persistent bovine tuberculosis on UK cattle farms. Prev. Vet. Med. 2007, 80, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Porphyre, T.; Stevenson, M.A.; McKenzie, J. Risk factors for bovine tuberculosis in New Zealand cattle farms and their relationship with possum control strategies. Prev. Vet. Med. 2008, 86, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Villaescusa, A.; Medley, G.; Mason, S.; Green, L. Risk factors for herd breakdown with bovine tuberculosis in 148 cattle herds in the south west of England. Prev. Vet. Med. 2010, 95, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessell, P.R.; Orton, R.; White, P.C.L.; Hutchings, M.R.; Kao, R.R. Risk factors for bovine Tuberculosis at the national level in Great Britain. BMC Vet. Res. 2012, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Lopez, B.; Barasona, J.A.; Gortazar, C.; Rodríguez-Prieto, V.; Sánchez-Vizcaíno, J.M.; Vicente, J. Farm-level risk factors for the occurrence, new infection or persistence of tuberculosis in cattle herds from South-Central Spain. Prev. Vet. Med. 2014, 116, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Broughan, J.M.; Judge, J.; Ely, E.; Delahay, R.J.; Wilson, G.; Clifton-Hadley, R.S.; Goodchild, A.V.; Bishop, H.; Parry, J.E.; Downs, S.H. A review of risk factors for bovine tuberculosis infection in cattle in the UK and Ireland. Epidemiol. Infect. 2016, 144, 2899–2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, L.; Gordon, A.; Abernethy, D.; Stevens, K. Bovine tuberculosis in Northern Ireland: Risk factors associated with time from post-outbreak test to subsequent herd breakdown. Prev. Vet. Med. 2014, 116, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humblet, M.-F.; Boschiroli, M.L.; Saegerman, C. Classification of worldwide bovine tuberculosis risk factors in cattle: A stratified approach. Vet. Res. 2009, 40, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humblet, M.-F.; Gilbert, M.; Govaerts, M.; Fauville-Dufaux, M.; Walravens, K.; Saegerman, C. New Assessment of Bovine Tuberculosis Risk Factors in Belgium Based on Nationwide Molecular Epidemiology. J. Clin. Microbiol. 2010, 48, 2802–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bénet, J.-J.; Dufour, B. Étude historique du risqué de résurgence de tuberculose bovine en France de 1992 à 2006. Épidémiologie Santé Anim 2009, 56, 107–115. [Google Scholar]
- Poirier, V. Elevages Foyers de Tuberculose Bovine Concernés Par l’abattage Partiel En France Depuis Son Autorisation Sur Tout Le Territoire En 2014: Typologie Des Élevages Concernés, Efficacité Du Dispositif; Ecole Nationale Vétérinaire d’Alfort: Maisons-Alfort, France, 2017. [Google Scholar]
- Crozet, G.; Dufour, B.; Rivière, J. Investigation of field intradermal tuberculosis test practices performed by veterinarians in France and factors that influence testing. Res. Vet. Sci. 2019, 124, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Delavenne, C.; Pandolfi, F.; Girard, S.; Réveillaud, É.; Boschiroli, M.-L.; Dommergues, L.; Garapin, F.; Keck, N.; Martin, F.; Moussu, M.; et al. Tuberculose Bovine: Bilan et Evolution de la Situation Epidémiologique Entre 2015 et 2016 en France Métropolitaine. Publication Anticipée du Bulletin Epidémiologique de l’Anses. 2019. Available online: https://be.anses.fr/sites/default/files/O-034_2019-06-05_Tub-Bilan_Delavenne_VF_par-ant.pdf (accessed on 16 March 2021).
- Palisson, A.; Courcoul, A.; Durand, B. Role of Cattle Movements in Bovine Tuberculosis Spread in France between 2005 and 2014. PLoS ONE 2016, 11, e0152578. [Google Scholar] [CrossRef] [PubMed]
- Avis de l’Anses Evaluation Du Risque Dans Le Cadre Du Dispositif de Surveillance de La Tuberculose Bovine. Anses. 2019. Available online: https://www.anses.fr/fr/system/files/SABA2018SA0186Ra.pdf (accessed on 16 March 2021).





| Size | Turnover | Surveillance Group | RRi |
|---|---|---|---|
| Small | <40% | At risk | Crisky |
| Former outbreak > 3 years | Crisky | ||
| None | 1 | ||
| ≥40% | At risk | Chigh turnover × Crisky | |
| Former outbreak > 3 years | Chigh turnover × Crisky | ||
| None | Chigh turnover | ||
| Big | <40% | At risk | Cbig × Crisky |
| Former outbreak > 3 years | Cbig × Crisky | ||
| None | Cbig | ||
| ≥40% | At risk | Cbig × Chigh turnover × Crisky | |
| Former outbreak > 3 years | Cbig × Chigh turnover × Crisky | ||
| None | Cbig × Chigh turnover |
| Overestimated Parameters | Underestimated Parameters |
|---|---|
| + probability for a herd to be “at risk” + for some areas, proportion of herds subject to periodic screening + probability for animals sold from “at risk” herds to be screened |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guétin-Poirier, V.; Rivière, J.; Dufour, B. Relative Contribution of Each Component of the French Ante-Mortem Surveillance System for Bovine Tuberculosis in Its Overall Sensitivity. Microorganisms 2021, 9, 643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030643
Guétin-Poirier V, Rivière J, Dufour B. Relative Contribution of Each Component of the French Ante-Mortem Surveillance System for Bovine Tuberculosis in Its Overall Sensitivity. Microorganisms. 2021; 9(3):643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030643
Chicago/Turabian StyleGuétin-Poirier, Valentine, Julie Rivière, and Barbara Dufour. 2021. "Relative Contribution of Each Component of the French Ante-Mortem Surveillance System for Bovine Tuberculosis in Its Overall Sensitivity" Microorganisms 9, no. 3: 643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030643