
In Silico Prediction and Analysis of Unusual Lantibiotic Resistance Operons in the Genus Corynebacterium


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids and Oligonucleotides
2.2. BLASTP Analyses
2.3. Prediction and Analysis of Bacteriocin Gene Clusters
2.4. Phylogenetic Analyses of Corynebacterium Species
2.5. Molecular Biology Procedures
2.6. Determination of Minimal Inhibitory Concentration (MIC)
2.7. Radial Streak Assay
3. Results
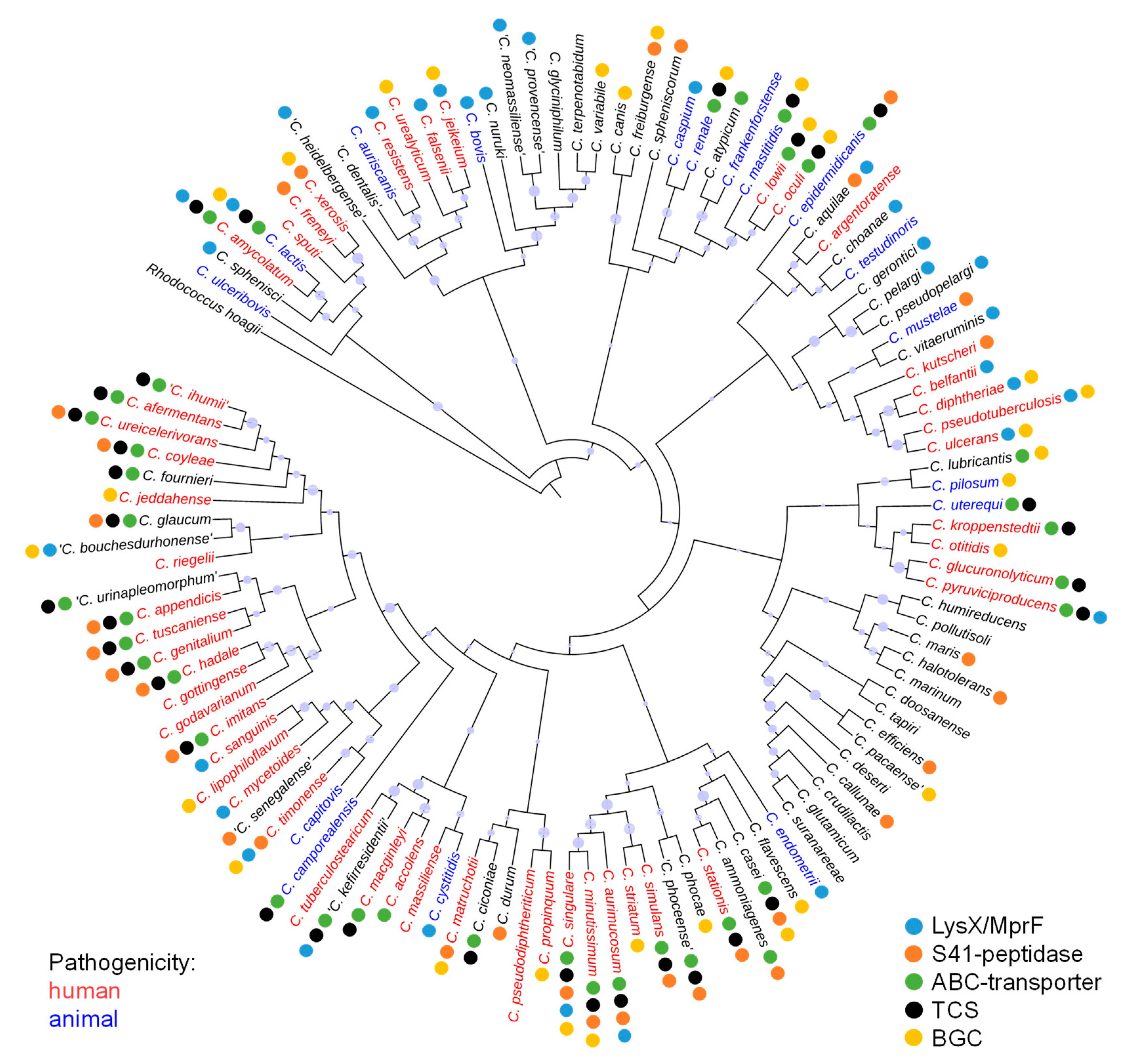
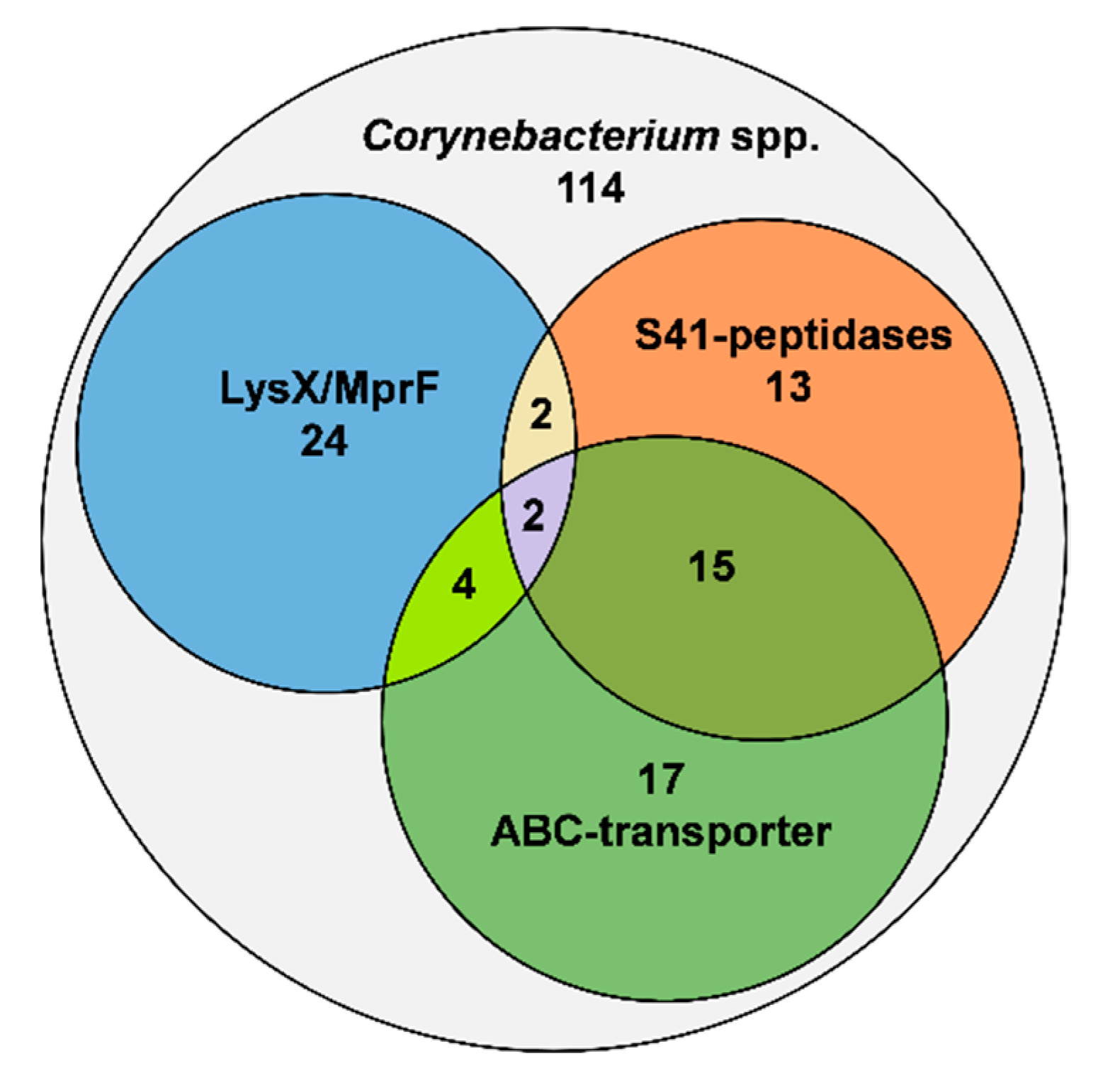
3.1. Distribution of Proteins Involved in Lantibiotic Resistance in the Genus Corynebacterium
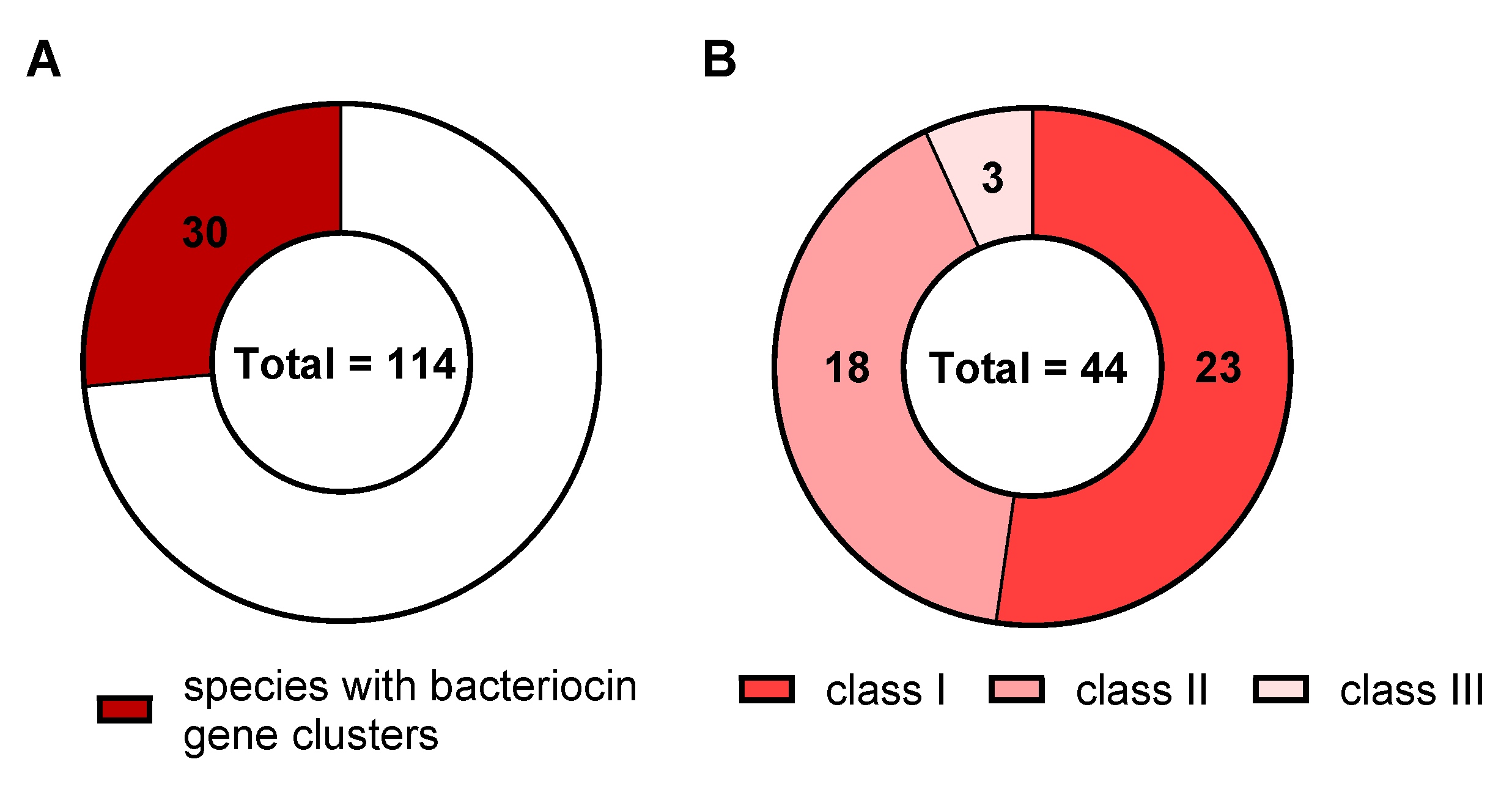
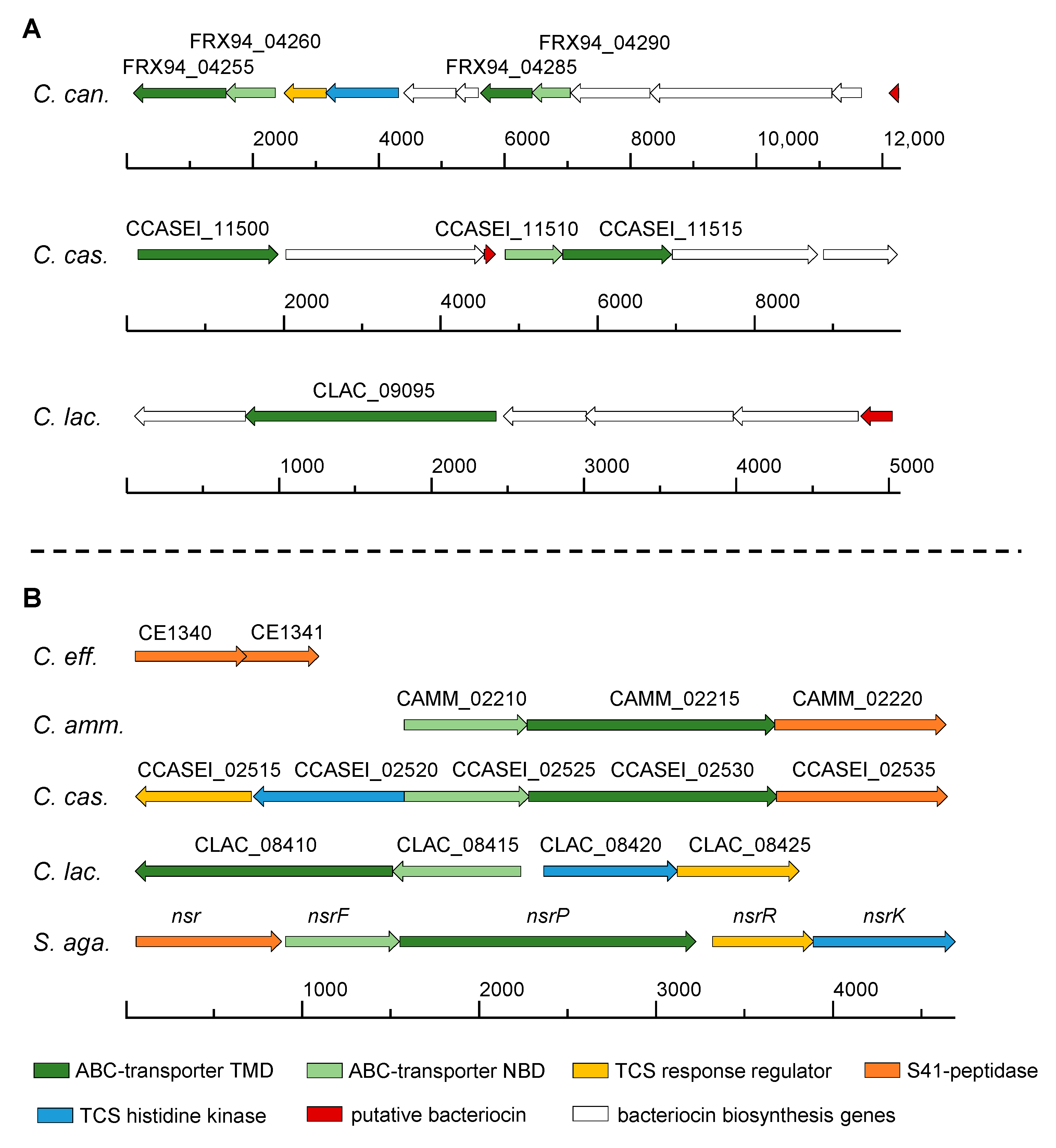
3.2. Distribution of Bacteriocin Gene Clusters in the Genus Corynebacterium
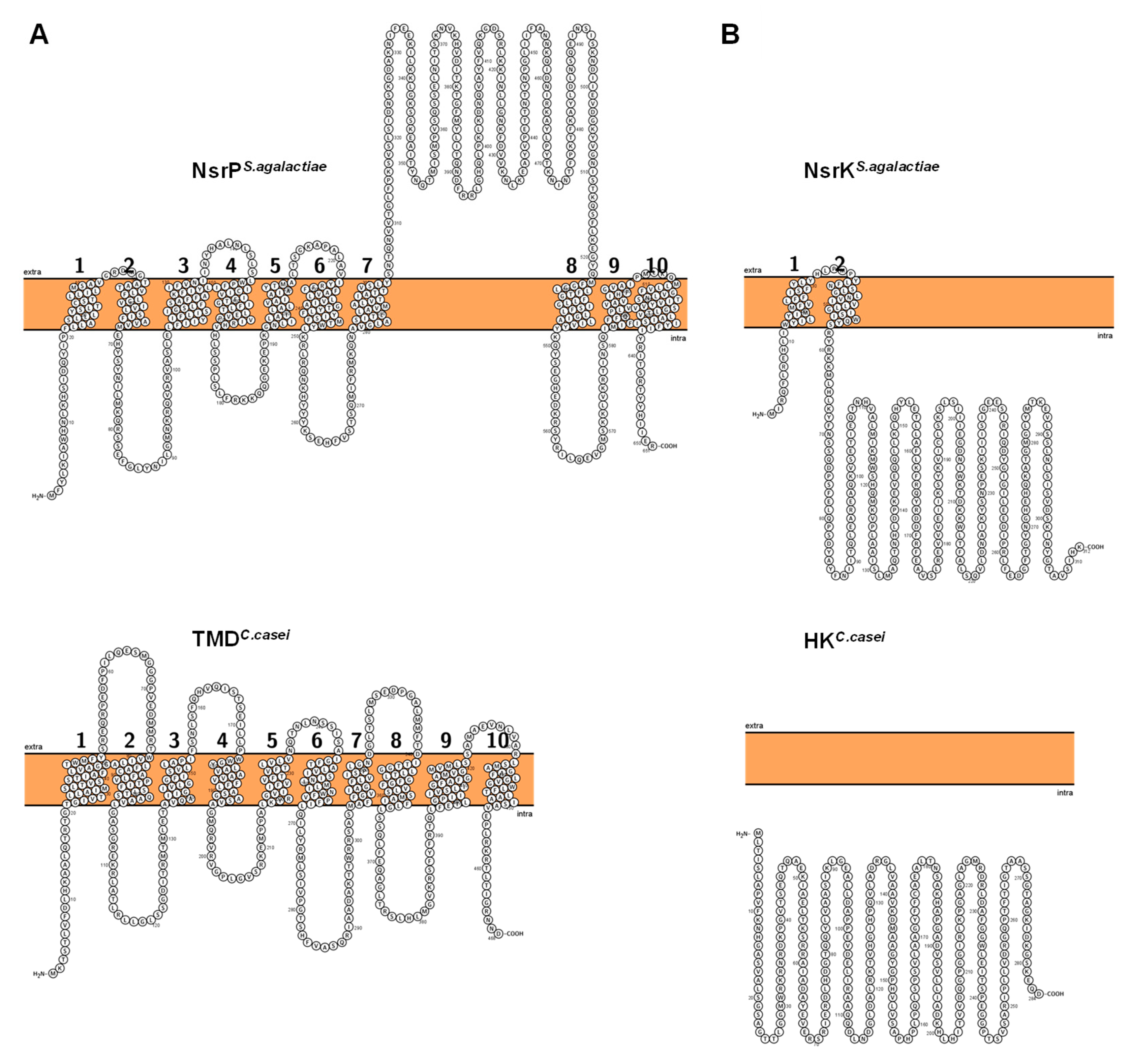
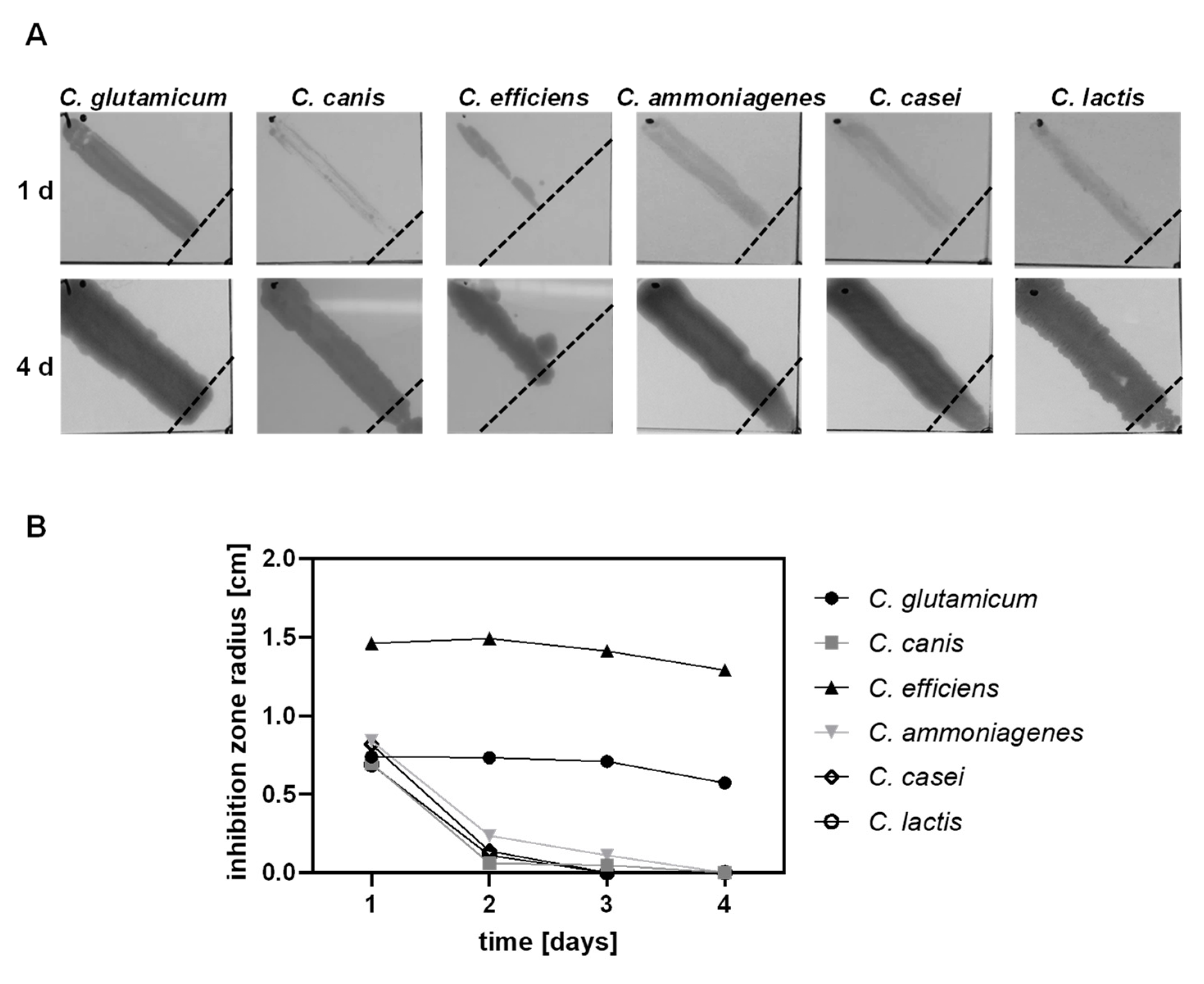
3.3. Analysis of Lantibiotic Resistance in Six Different Corynebacterium Species
3.4. Heterologous Expression of Putative Lantibiotic Resistance Operons in C. glutamicum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Na, Y.-A.; Kim, E.; Lee, H.-S.; Kim, P. The Actinobacterium Corynebacterium glutamicum, an Industrial Workhorse. J. Microbiol. Biotechnol. 2016, 26, 807–822. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, S.; Yonetani, Y.; Maruyama, A.; Teshiba, S. Production of riboflavin by metabolically engineered Corynebacterium ammoniagenes. Appl. Microbiol. Biotechnol. 2000, 53, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Hoskisson, P.A. Microbe Profile: Corynebacterium diphtheriae – an old foe always ready to seize opportunity. Microbiology 2018, 164, 865–867. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Dangel, A.; Peters, M.; Mühldorfer, K.; Braune, S.; Eisenberg, T.; Szentiks, C.A.; Rau, J.; Konrad, R.; Hörmansdorfer, S.; et al. Tox-positive Corynebacterium ulcerans in hedgehogs, Germany. Emerg. Microbes Infect. 2019, 8, 211–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, K.S.; White, J.M.; Lucenko, I.; Mercer, D.; Crowcroft, N.S.; Neal, S. Diphtheria in the Postepidemic. Emerg. Infect. Dis. 2012, 18, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Bernard, K. The Genus Corynebacterium and Other Medically Relevant Coryneform-Like Bacteria. J. Clin. Microbiol. 2012, 50, 3152–3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nhan, T.-X.; Parienti, J.-J.; Badiou, G.; Leclercq, R.; Cattoir, V. Microbiological investigation and clinical significance of Corynebacterium spp. in respiratory specimens. Diagn. Microbiol. Infect. Dis. 2012, 74, 236–241. [Google Scholar] [CrossRef] [PubMed]
- McMullen, A.R.; Anderson, N.; Wallace, M.A.; Shupe, A.; Burnham, C.-A.D. When Good Bugs Go Bad: Epidemiology and Antimicrobial Resistance Profiles of Corynebacterium striatum, an Emerging Multidrug-Resistant, Opportunistic Pathogen. Antimicrob. Agents Chemother. 2017, 61, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cazanave, C.; Greenwood-Quaintance, K.E.; Hanssen, A.D.; Patel, R. Corynebacterium Prosthetic Joint Infection. J. Clin. Microbiol. 2012, 50, 1518–1523. [Google Scholar] [CrossRef] [Green Version]
- Díez-Aguilar, M.; Ruiz-Garbajosa, P.; Fernández-Olmos, A.; Guisado, P.; Del Campo, R.; Quereda, C.; Cantón, R.; Meseguer, M.A. Non-diphtheriae Corynebacterium species: An emerging respiratory pathogen. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 769–772. [Google Scholar] [CrossRef]
- Pardo, S.M.M.; Patel, R.H.; Ramsakal, A.; Greene, J. Disseminated Corynebacterium jeikeium Infection in Cancer Patients. Cureus 2020, 12, e8764. [Google Scholar] [CrossRef]
- Sierra, J.M.; Martinez-Martinez, L.; Vázquez, F.; Giralt, E.; Vila, J. Relationship between Mutations in the gyrA Gene and Quinolone Resistance in Clinical Isolates of Corynebacterium striatum and Corynebacterium amycolatum. Antimicrob. Agents Chemother. 2005, 49, 1714–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deguchi, H.; Kitazawa, K.; Kayukawa, K.; Kondoh, E.; Fukumoto, A.; Yamasaki, T.; Kinoshita, S.; Sotozono, C. The trend of resistance to antibiotics for ocular infection of Staphylococcus aureus, coagulase-negative staphylococci, and Corynebacterium compared with 10-years previous: A retrospective observational study. PLoS ONE 2018, 13, e0203705. [Google Scholar] [CrossRef]
- Otsuka, Y.; Ohkusu, K.; Kawamura, Y.; Baba, S.; Ezaki, T.; Kimura, S. Emergence of multidrug-resistant Corynebacterium striatum as a nosocomial pathogen in long-term hospitalized patients with underlying diseases. Diagn. Microbiol. Infect. Dis. 2006, 54, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2012, 30, 108–160. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Ben Said, L.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45. [Google Scholar] [CrossRef]
- Delves-Broughton, J.; Blackburn, P.; Evans, R.J.; Hugenholtz, J. Applications of the bacteriocin, nisin. Antonie van Leeuwenhoek 1996, 69, 193–202. [Google Scholar] [CrossRef]
- Brötz, H.; Josten, M.; Wiedemann, I.; Schneider, U.; Götz, F.; Bierbaum, G.; Sahl, H.-G. Role of lipid-bound peptidoglycan precursors in the formation of pores by nisin, epidermin and other lantibiotics. Mol. Microbiol. 1998, 30, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasper, H.E.; Kramer, N.E.; Smith, J.L.; Hillman, J.D.; Zachariah, C.; Kuipers, O.P.; De Kruijff, B.; Breukink, E. An Alternative Bactericidal Mechanism of Action for Lantibiotic Peptides That Target Lipid II. Science 2006, 313, 1636–1637. [Google Scholar] [CrossRef] [Green Version]
- Riley, M.A.; Wertz, J.E. Bacteriocins: Evolution, Ecology, and Application. Annu. Rev. Microbiol. 2002, 56, 117–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhatib, Z.; Lagedroste, M.; Zaschke, J.; Wagner, M.; Abts, A.; Fey, I.; Kleinschrodt, D.; Smits, S.H.J. The C-terminus of nisin is important for the ABC transporter Nis FEG to confer immunity in Lactococcus lactis. MicrobiologyOpen 2014, 3, 752–763. [Google Scholar] [CrossRef]
- Alkhatib, Z.; Lagedroste, M.; Fey, I.; Kleinschrodt, D.; Abts, A.; Smits, S.H.J. Lantibiotic Immunity: Inhibition of Nisin Mediated Pore Formation by NisI. PLoS ONE 2014, 9, e102246. [Google Scholar] [CrossRef] [PubMed]
- Draper, L.A.; Cotter, P.D.; Hill, C.; Ross, R.P. Lantibiotic Resistance. Microbiol. Mol. Biol. Rev. 2015, 79, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Neuhaus, F.C.; Baddiley, J. A Continuum of Anionic Charge: Structures and Functions of d-Alanyl-Teichoic Acids in Gram-Positive Bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 686–723. [Google Scholar] [CrossRef] [Green Version]
- Roy, H.; Ibba, M. RNA-dependent lipid remodeling by bacterial multiple peptide resistance factors. Proc. Natl. Acad. Sci. USA 2008, 105, 4667–4672. [Google Scholar] [CrossRef] [Green Version]
- Roy, H. Tuning the properties of the bacterial membrane with aminoacylated phosphatidylglycerol. IUBMB Life 2009, 61, 940–953. [Google Scholar] [CrossRef] [Green Version]
- Peschel, A.; Jack, R.W.; Otto, M.; Collins, L.V.; Staubitz, P.; Nicholson, G.; Kalbacher, H.; Nieuwenhuizen, W.F.; Jung, G.; Tarkowski, A.; et al. Staphylococcus aureus Resistance to Human Defensins and Evasion of Neutrophil Killing via the Novel Virulence Factor Mprf Is Based on Modification of Membrane Lipids with l-Lysine. J. Exp. Med. 2001, 193, 1067–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, E.; Stankowska, D.; Zhang, J.; Fol, M.; Cheng, Q.-J.; Lun, S.; Bishai, W.R.; Rajagopalan, M.; Chatterjee, D.; Madiraju, M.V. The Two-Domain LysX Protein of Mycobacterium tuberculosis Is Required for Production of Lysinylated Phosphatidylglycerol and Resistance to Cationic Antimicrobial Peptides. PLoS Pathog. 2009, 5, e1000534. [Google Scholar] [CrossRef]
- Hachmann, A.-B.; Angert, E.R.; Helmann, J.D. Genetic Analysis of Factors Affecting Susceptibility of Bacillus subtilis to Daptomycin. Antimicrob. Agents Chemother. 2009, 53, 1598–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBride, S.M.; Sonenshein, A.L. Identification of a Genetic Locus Responsible for Antimicrobial Peptide Resistance inClostridium difficile. Infect. Immun. 2010, 79, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemens, R.; Zaschke-Kriesche, J.; Khosa, S.; Smits, S.H.J. Insight into Two ABC Transporter Families Involved in Lantibiotic Resistance. Front. Mol. Biosci. 2018, 4, 91. [Google Scholar] [CrossRef] [Green Version]
- Dintner, S.; Heermann, R.; Fang, C.; Jung, K.; Gebhard, S. A Sensory Complex Consisting of an ATP-binding Cassette Transporter and a Two-component Regulatory System Controls Bacitracin Resistance in Bacillus subtilis. J. Biol. Chem. 2014, 289, 27899–27910. [Google Scholar] [CrossRef] [Green Version]
- Hiron, A.; Falord, M.; Valle, J.; Débarbouillé, M.; Msadek, T. Bacitracin and nisin resistance in Staphylococcus aureus: A novel pathway involving the BraS/BraR two-component system (SA2417/SA2418) and both the BraD/BraE and VraD/VraE ABC transporters. Mol. Microbiol. 2011, 81, 602–622. [Google Scholar] [CrossRef]
- Reiners, J.; Lagedroste, M.; Ehlen, K.; Leusch, S.; Zaschke-Kriesche, J.; Smits, S.H.J. The N-terminal Region of Nisin Is Important for the BceAB-Type ABC Transporter NsrFP from Streptococcus agalactiae COH1. Front. Microbiol. 2017, 8, 1643. [Google Scholar] [CrossRef] [Green Version]
- Dintner, S.; Staron, A.; Berchtold, E.; Petri, T.; Mascher, T.; Gebhard, S. Coevolution of ABC Transporters and Two-Component Regulatory Systems as Resistance Modules against Antimicrobial Peptides in Firmicutes Bacteria. J. Bacteriol. 2011, 193, 3851–3862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, J.M.; Edwards, A.N.; McBride, S.M. The Clostridium difficile cpr Locus Is Regulated by a Noncontiguous Two-Component System in Response to Type A and B Lantibiotics. J. Bacteriol. 2013, 195, 2621–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.; Teng, K.; Wang, J.; Zhao, F.; Zhang, J.; Zhong, J. Identification of Key Residues in the NisK Sensor Region for Nisin Biosynthesis Regulation. Front. Microbiol. 2017, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Khosa, S.; Alkhatib, Z.; Smits, S.H. NSR from Streptococcus agalactiae confers resistance against nisin and is encoded by a conserved nsr operon. Biol. Chem. 2013, 394, 1543–1549. [Google Scholar] [CrossRef]
- Collins, M.D. Transfer of Brevibacterium ammoniagenes (Cooke and Keith) to the Genus Corynebacterium as Corynebacterium ammoniagenes comb. nov. Int. J. Syst. Bacteriol. 1987, 37, 442–443. [Google Scholar] [CrossRef] [Green Version]
- Wiertz, R.; Schulz, S.C.; Müller, U.; Kämpfer, P.; Lipski, A. Corynebacterium frankenforstense sp. nov. and Corynebacterium lactis sp. nov., isolated from raw cow milk. Int. J. Syst. Evol. Microbiol. 2013, 63, 4495–4501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fudou, R.; Jojima, Y.; Seto, A.; Yamada, K.; Kimura, E.; Nakamatsu, T.; Hiraishi, A.; Yamanaka, S. Corynebacterium efficiens sp. nov., a glutamic-acid-producing species from soil and vegetables. Int. J. Syst. Evol. Microbiol. 2002, 52, 1127–1131. [Google Scholar] [CrossRef] [Green Version]
- Brennan, N.M.; Brown, R.; Goodfellow, M.; Ward, A.C.; Beresford, T.P.; Simpson, P.J.; Fox, P.F.; Cogan, T.M. Corynebacterium mooreparkense sp. nov. and Corynebacterium casei sp. nov., isolated from the surface of a smear-ripened cheese. Int. J. Syst. Evol. Microbiol. 2001, 51, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funke, G.; Englert, R.; Frodl, R.; Bernard, K.A.; Stenger, S. Corynebacterium canis sp. nov., isolated from a wound infection caused by a dog bite. Int. J. Syst. Evol. Microbiol. 2010, 60, 2544–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, J.; Treptow, C.; Eggeling, L.; Sahm, H. Regulation of Enzymes of Lysine Biosynthesis in Corynebacterium glutamicum. Microbiology 1988, 134, 3221–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.I.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes11Edited by F. Cohen. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Heel, A.J.; De Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Van Der Rest, M.E.; Lange, C.; Molenaar, D. A heat shock following electroporation induces highly efficient transformation of Corynebacterium glutamicum with xenogeneic plasmid DNA. Appl. Microbiol. Biotechnol. 1999, 52, 541–545. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Coman, M.; Verdenelli, M.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Boyko, N.; Cresci, A. In vitro evaluation of antimicrobial activity of Lactobacillus rhamnosus IMC 501®, Lactobacillus paracasei IMC 502® and SYNBIO® against pathogens. J. Appl. Microbiol. 2014, 117, 518–527. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat Methods. 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peschel, A.; Otto, M.; Jack, R.W.; Kalbacher, H.; Jung, G.; Götz, F. Inactivation of the dlt Operon inStaphylococcus aureus Confers Sensitivity to Defensins, Protegrins, and Other Antimicrobial Peptides. J. Biol. Chem. 1999, 274, 8405–8410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abachin, E.; Poyart, C.; Pellegrini, E.; Milohanic, E.; Fiedler, F.; Berche, P.; Trieu-Cuot, P. Formation of D-alanyl-lipoteichoic acid is required for adhesion and virulence of Listeria monocytogenes. Mol. Microbiol. 2002, 43, 1–14. [Google Scholar] [CrossRef]
- Sun, Z.; Zhong, J.; Liang, X.; Liu, J.; Chen, X.; Huan, L. Novel Mechanism for Nisin Resistance via Proteolytic Degradation of Nisin by the Nisin Resistance Protein NSR. Antimicrob. Agents Chemother. 2009, 53, 1964–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosa, S.; Lagedroste, M.; Smits, S.H.J. Protein Defense Systems against the Lantibiotic Nisin: Function of the Immunity Protein NisI and the Resistance Protein NSR. Front. Microbiol. 2016, 7, 504. [Google Scholar] [CrossRef]
- Keiler, K.C.; Sauer, R.T. Identification of Active Site Residues of the Tsp Protease. J. Biol. Chem. 1995, 270, 28864–28868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takala, T.M.; Saris, P.E.J. C terminus of NisI provides specificity to nisin. Microbiology 2006, 152, 3543–3549. [Google Scholar] [CrossRef] [Green Version]
- Kallenberg, F.; Dintner, S.; Schmitz, R.; Gebhard, S. Identification of Regions Important for Resistance and Signalling within the Antimicrobial Peptide Transporter BceAB of Bacillus subtilis. J. Bacteriol. 2013, 195, 3287–3297. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, B.; Mahoney, R. Inactivation of Nisin by Alpha-Chymotrypsin. J. Dairy Sci. 1969, 52, 1448–1450. [Google Scholar] [CrossRef]
- Fontana, M.; Bastos, M.; Brandelli, A. Peptide-like substances as antimicrobial barriers to Corynebacterium sp. adhesion to silicone catheters. Lett. Appl. Microbiol. 2007, 45, 324–329. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Reference |
|---|---|---|
| Corynebacterium glutamicum | ATCC 13032 | ATCC a |
| Corynebacterium ammoniagenes | DSM 20306 | [42] |
| Corynebacterium lactis | RW3-42 | [43] |
| Corynebacterium efficiens | DSM 44549 | [44] |
| Corynebacterium casei | DSM 44701 | [45] |
| Corynebacterium canis | DSM 45402 | [46] |
| Plasmid | Characteristics | |
| pJC1 | E. coli/C. glutamicum shuttle vector; | [47] |
| pJC_nsrPFKRClac | pJC1-derivative with nsr-like operon of C. lactis RW3-42 | This study |
| pJC_nsrRKFPCcas | pJC1-derivative with nsr-like operons of C. casei DSM 44701 lacking the peptidase gene nsrX | This study |
| pJC_nsrRKFPXCcas | pJC1-derivative with nsr-like operons of C. casei DSM 44701; | This study |
| pJC_nsrFPCamm | pJC1-derivative with nsr-like operon of C. ammoniagenes DSM 20306 lacking the peptidase gene nsrX; | This study |
| pJC_nsrFPXCamm | pJC1-derivative with nsr-like operon of C. ammoniagenes DSM 20306; | This study |
| Species | Strain | BGC | LanI | CprABC | BceAB | NSR | HK | RR | LysX |
|---|---|---|---|---|---|---|---|---|---|
| C. glutamicum | ATCC 13032 | - | - | - | - | - | - | - | - |
| C. ammoniagenes | DSM 20306 | - | - | - | + | + | - | - | - |
| C. lactis | RW3-42 | + | - | - | + | - | + | + | + |
| C. efficiens | DSM 44549 | - | - | - | - | (+) a | - | - | - |
| C. casei | DSM 44701 | + | - | - | + | + | + | + | - |
| C. canis | DSM 45402 | + | - | - | - | - | - | - | - |
| Strain | MIC [µg/mL] | |
|---|---|---|
| Uninduced | Induced | |
| C. glutamicum ATCC 13032 | 1.25 | 1.25 |
| C. canis DSM 45402 | 5.00 | 5.00 |
| C. efficiens DSM 44549 | 0.31 | 0.16 |
| C. ammoniagenes DSM 20306 | 1.25 | 5.00 |
| C. casei DSM 44701 | 0.63 | 1.25 |
| C. lactis RW3-42 | 12.50 | 25.00 |
| Strain | MIC [µg/mL] | |
|---|---|---|
| Uninduced | Induced | |
| C. glutamicum ATCC 13032/pJC1 | 1.25 | 1.25 |
| C. glutamicum ATCC 13032/pJC1_nsrPFKRClac | 1.25 | 1.25 |
| C. glutamicum ATCC 13032/pJC1_nsrRKFPCcas | 2.50 | 2.50 |
| C. glutamicum ATCC 13032/pJC1_nsrRKFPXCcas | 5.00 | 5.00 |
| C. glutamicum ATCC 13032/pJC1_nsrFPCamm | 1.25 | 1.25 |
| C. glutamicum ATCC 13032/pJC1_nsrFPXCamm | 5.00 | 5.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldbeck, O.; Weixler, D.; Eikmanns, B.J.; Riedel, C.U. In Silico Prediction and Analysis of Unusual Lantibiotic Resistance Operons in the Genus Corynebacterium. Microorganisms 2021, 9, 646. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030646
Goldbeck O, Weixler D, Eikmanns BJ, Riedel CU. In Silico Prediction and Analysis of Unusual Lantibiotic Resistance Operons in the Genus Corynebacterium. Microorganisms. 2021; 9(3):646. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030646
Chicago/Turabian StyleGoldbeck, Oliver, Dominik Weixler, Bernhard J. Eikmanns, and Christian U. Riedel. 2021. "In Silico Prediction and Analysis of Unusual Lantibiotic Resistance Operons in the Genus Corynebacterium" Microorganisms 9, no. 3: 646. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030646