
CRISPRi-Library-Guided Target Identification for Engineering Carotenoid Production by Corynebacterium glutamicum

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
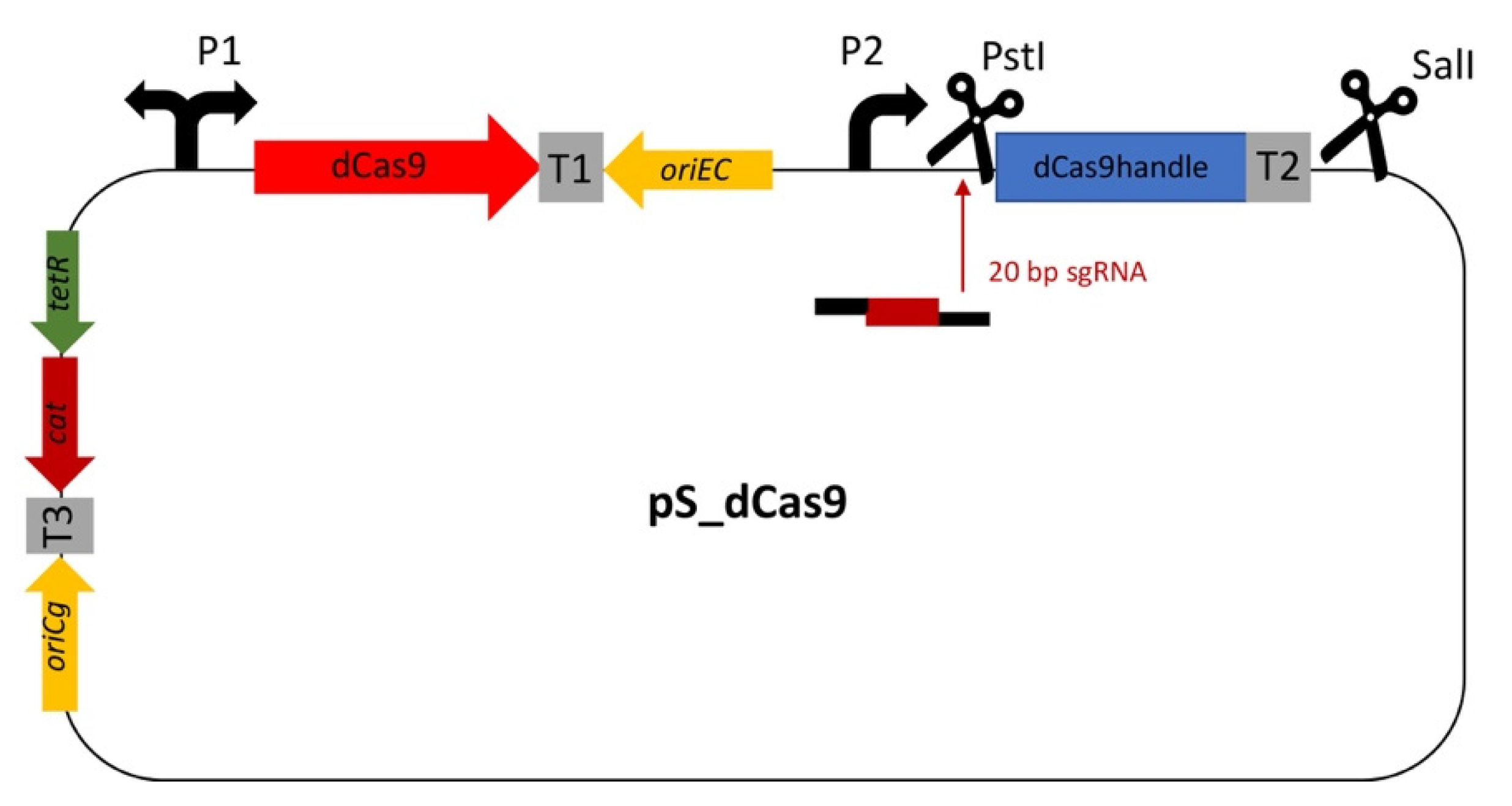
2.1. Construction of the CRISPRi Vector System
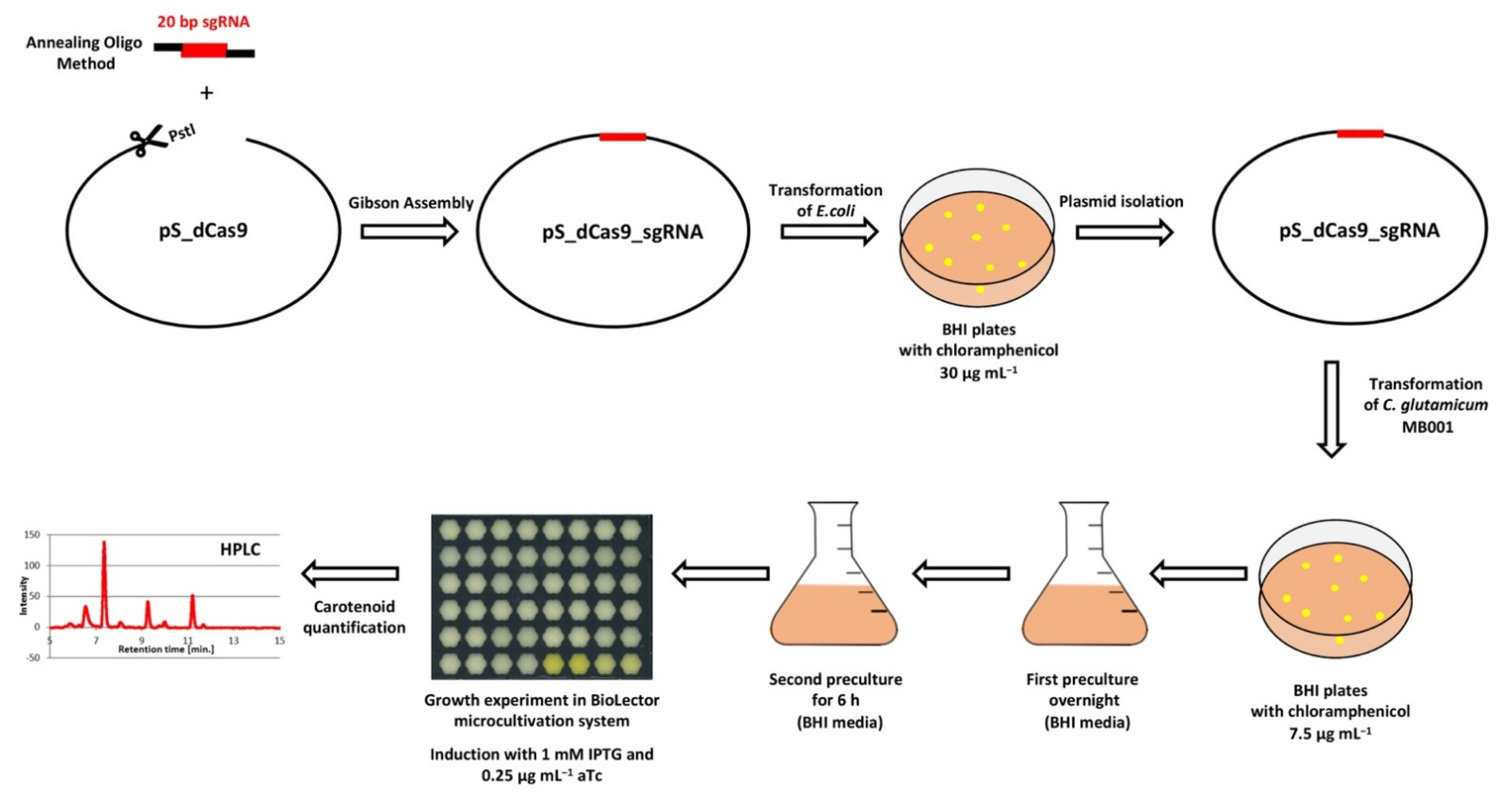
2.2. Construction of the CRISPRi Library
2.3. Construction of C. glutamicum Deletion Mutants
2.4. Quantification of the mRNA Levels of Targeted Cells by CRISPRi
2.5. Carotenoid Quantification
3. Results
3.1. Design and Initial Testing of a CRISPRi Library for Gene Repression in C. glutamicum
3.1.1. Construction of a Vector CRISPRi Library for C. glutamicum
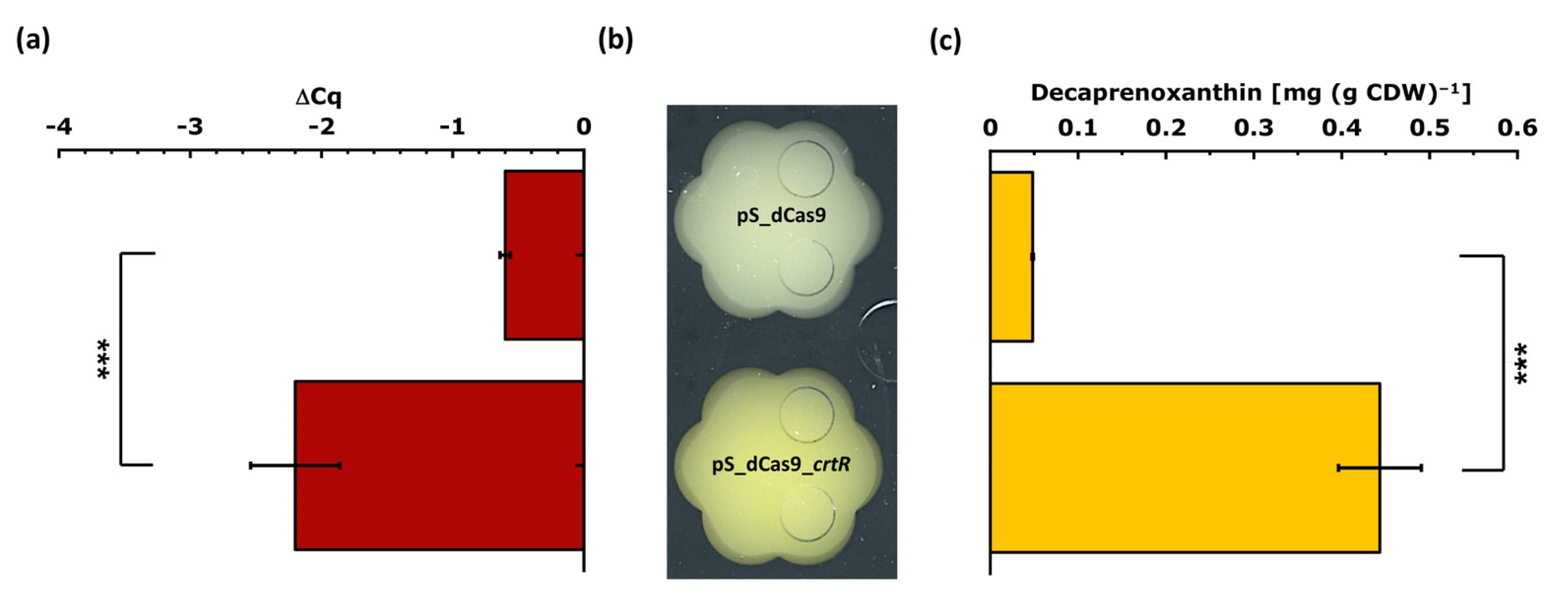
3.1.2. Testing of the CRISPRi Library Vector for the Repression of crtR
3.2. Characterization of a CRISPRi Library to Interrogate 74 Target Genes with Potential Relevance for Carotenogenesis in C. glutamicum
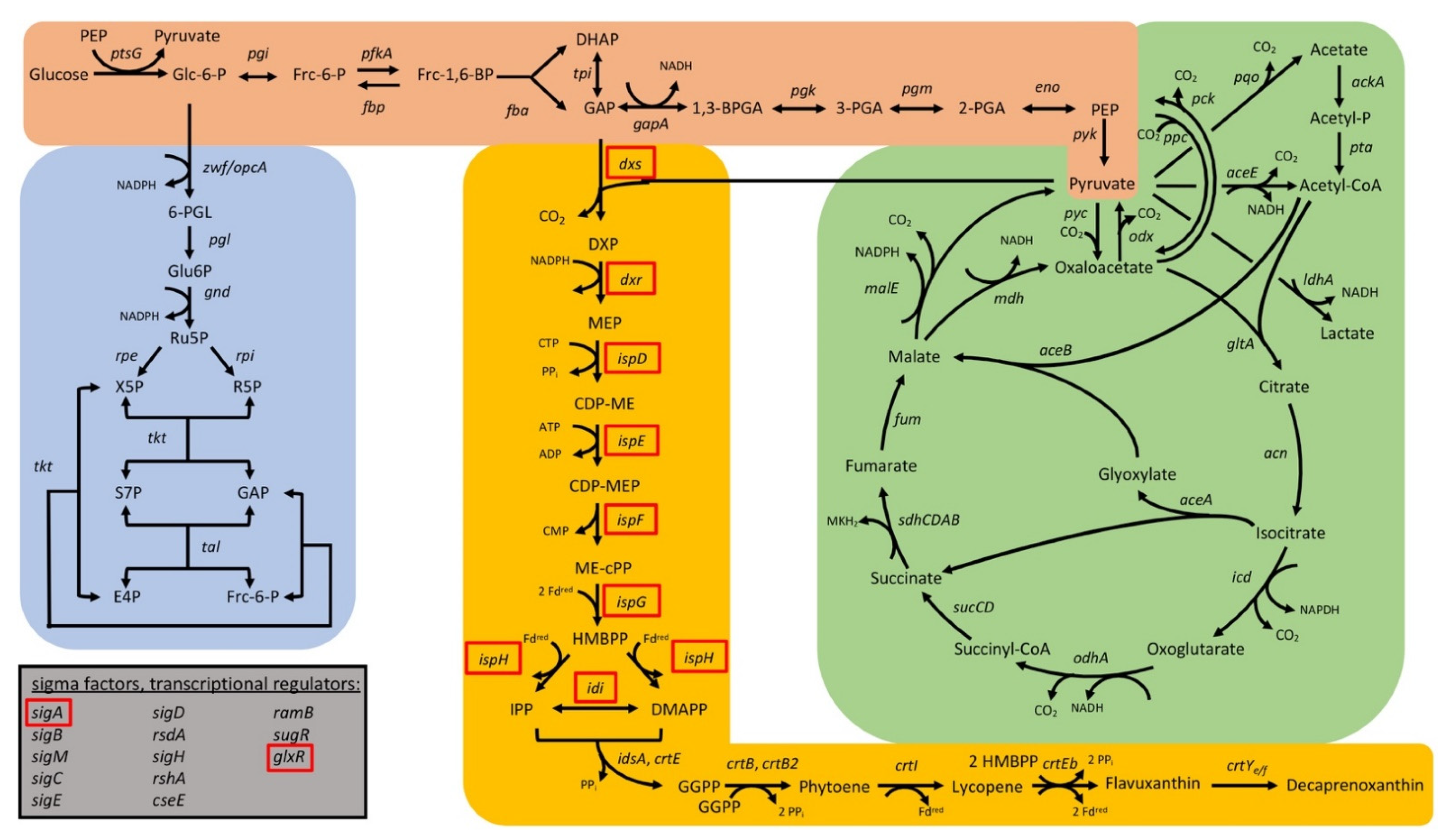
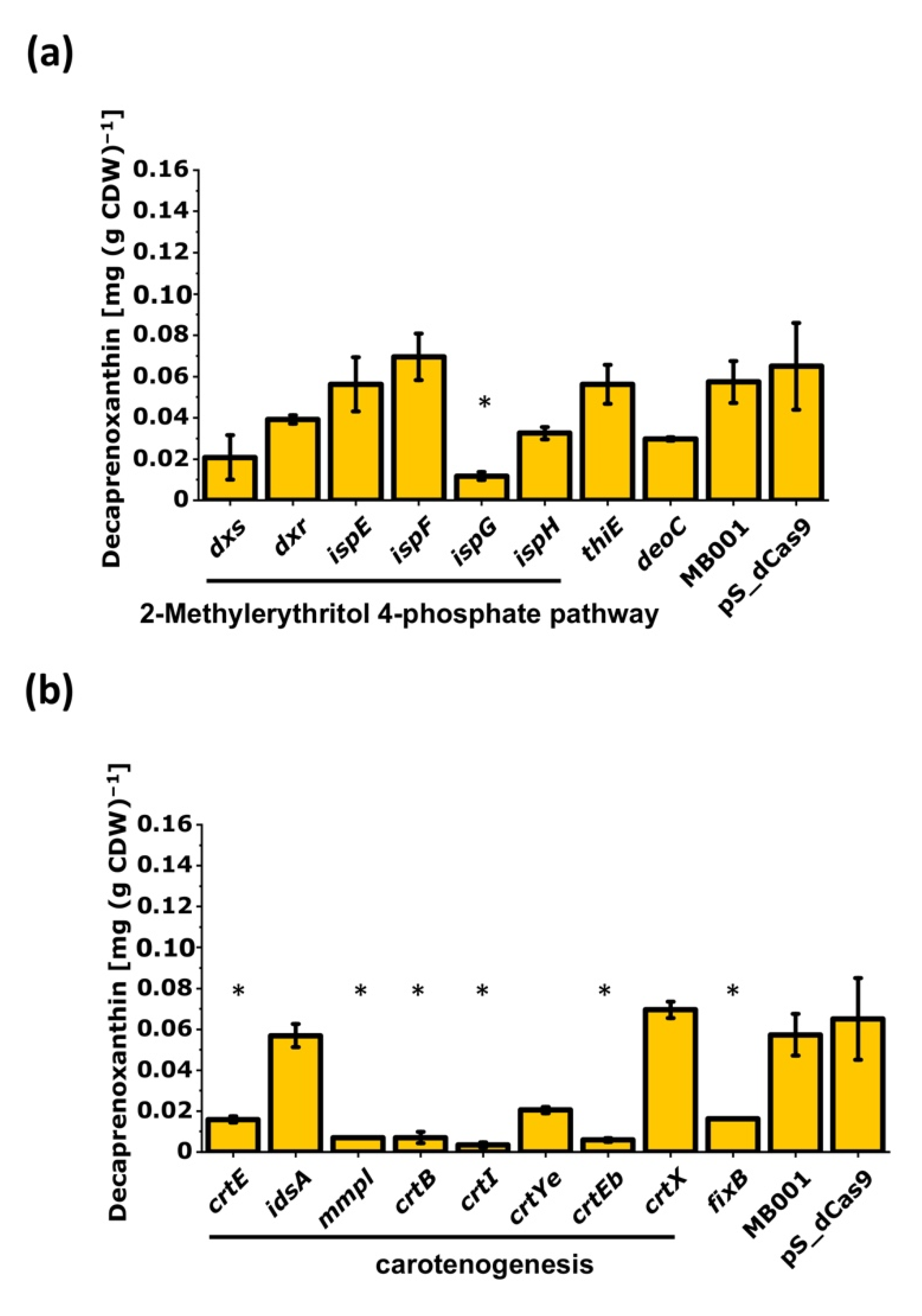
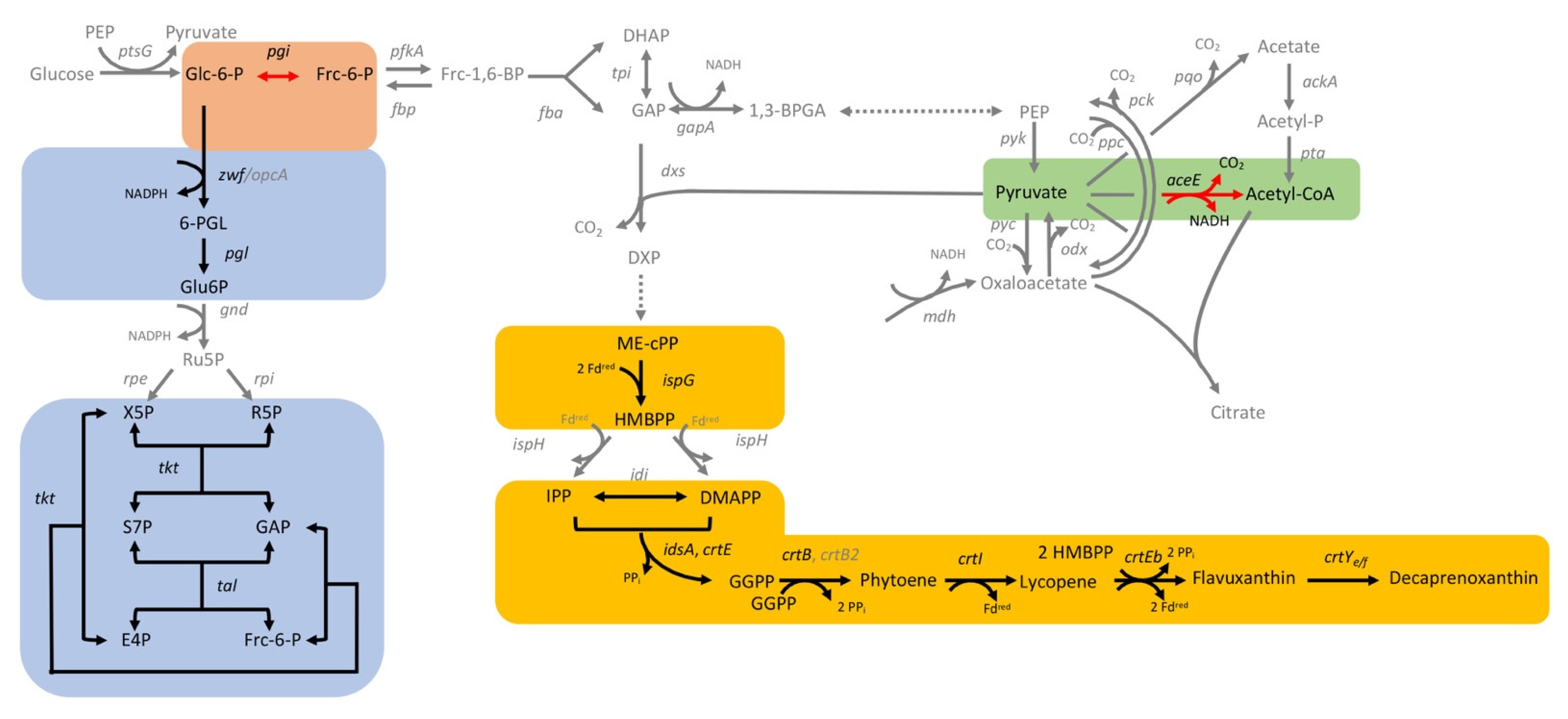
3.2.1. CRISPRi-Based Repression of Genes of the MEP Pathway and of Carotenogenesis-Reduced Decaprenoxanthin Pigmentation
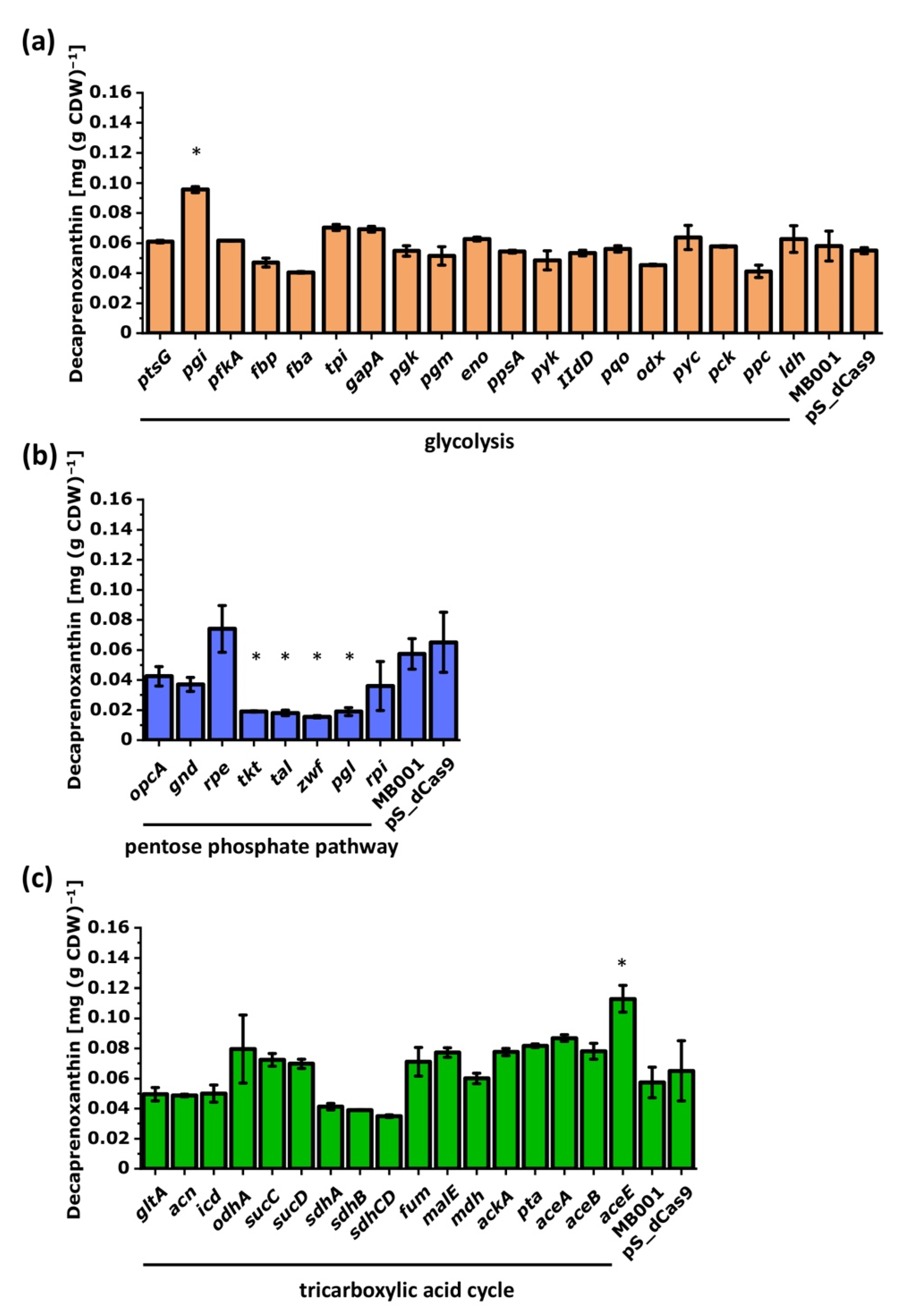
3.2.2. CRISPRi-Based Repression of Genes of the Central-Carbon-Metabolism-Identified Supply of GAP and Entry into the Pentose Phosphate Pathway as Potential Bottlenecks in Decaprenoxanthin Biosynthesis
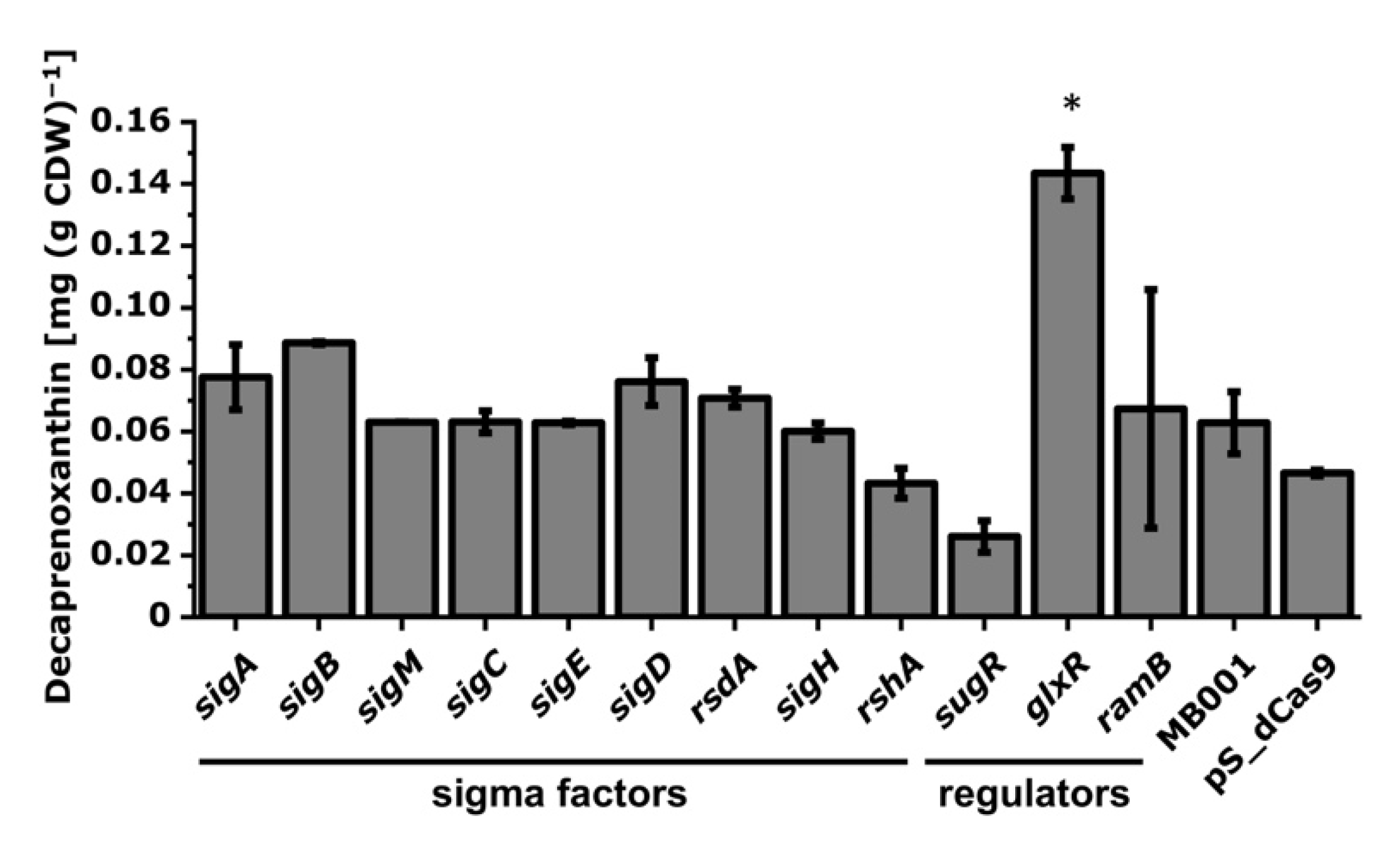
3.2.3. Interrogation of Regulatory Genes by CRISPRi with Respect to Carotenoid Biosynthesis in C. glutamicum
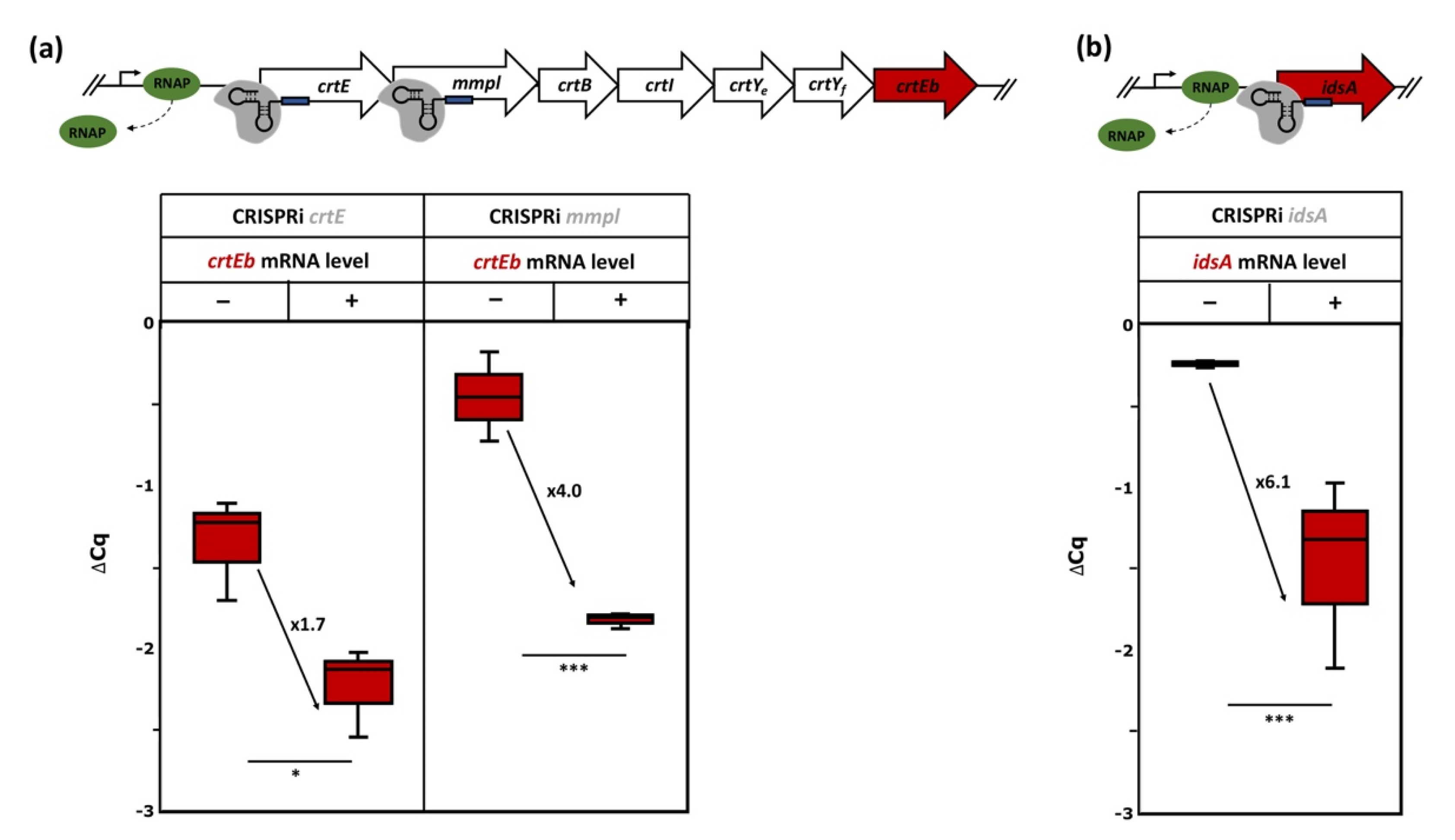
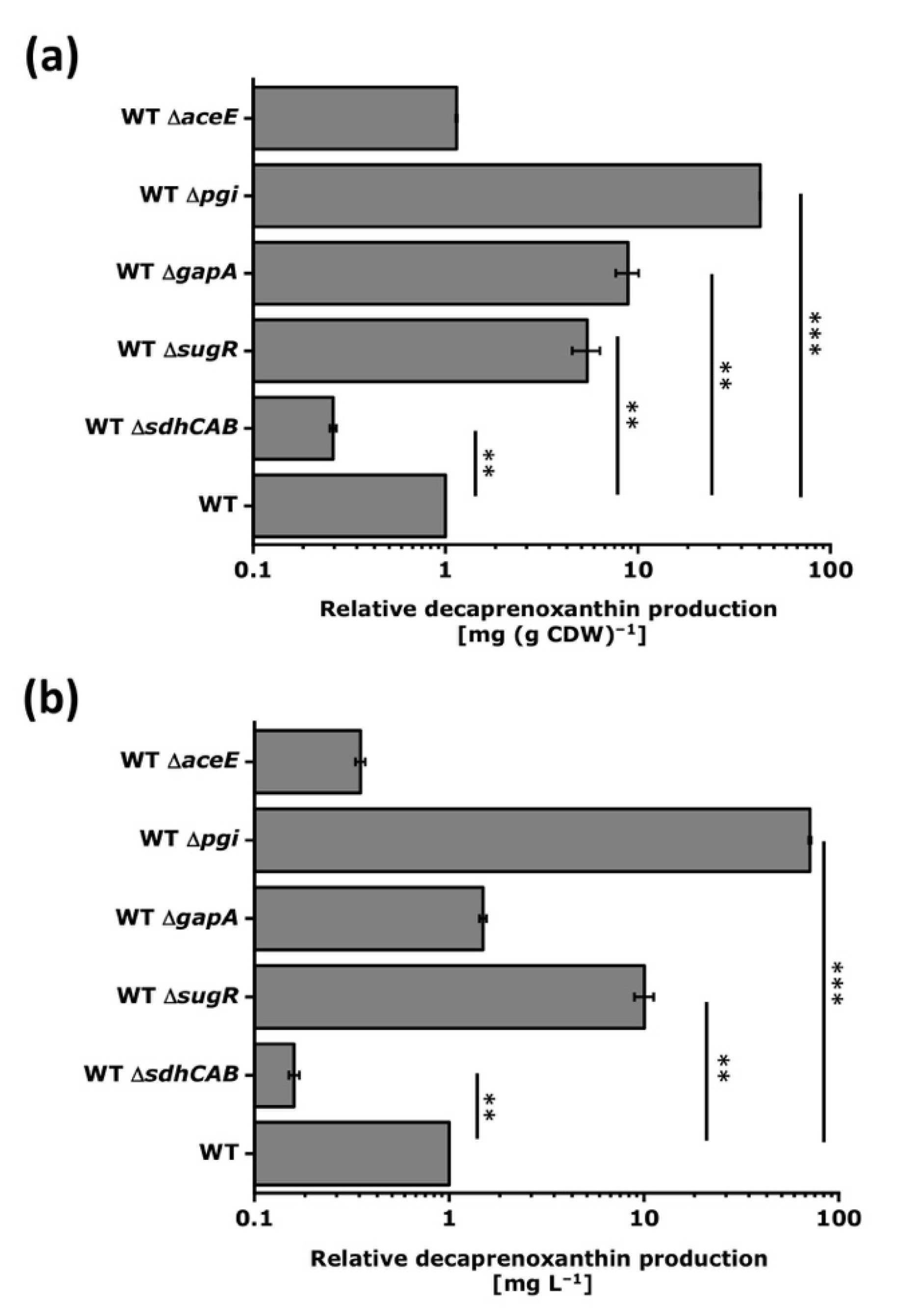
3.3. Deletion of Selected Target Genes Identified by CRISPRi Repression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adli, M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018, 9, 1911. [Google Scholar] [CrossRef]
- Lee, J.-H.; Wendisch, V.F. Production of amino acids—Genetic and metabolic engineering approaches. Bioresour. Technol. 2017, 245, 1575–1587. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the Immune System of Bacteria and Archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Larson, M.H.; Gilbert, L.A.; Wang, X.; Lim, W.A.; Weissman, J.S.; Qi, L.S. CRISPR interference (CRISPRi) for sequence-specific control of gene expression. Nat. Protoc. 2013, 8, 2180–2196. [Google Scholar] [CrossRef] [PubMed]
- Schultenkämper, K.; Brito, L.F.; Wendisch, V.F. Impact of CRISPR interference on strain development in biotechnology. Biotechnol. Appl. Biochem. 2020, 67, 7–21. [Google Scholar] [CrossRef]
- Li, S.; Jendresen, C.B.; Landberg, J.; Pedersen, L.E.; Sonnenschein, N.; Jensen, S.I.; Nielsen, A.T. Genome-wide CRISPRi-based identification of targets for decoupling growth from production. ACS Synth. Biol. 2020, 9, 1030–1040. [Google Scholar] [CrossRef]
- Zhao, H.; Sun, Y.; Peters, J.M.; Gross, C.A.; Garner, E.C.; Helmann, J.D. Depletion of undecaprenyl pyrophosphate phosphatases disrupts cell envelope biogenesis in Bacillus subtilis. J. Bacteriol. 2016, 198, 2925–2935. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.E.; Sburlati, A.; Hatzimanikatis, V.; Lee, K.; Renner, W.A.; Tsai, P.S. Inverse Metabolic Engineering: A Strategy for Directed Genetic Engineering of Useful Phenotypes. Biotechnol. Bioeng. 1996, 52, 13. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, H.; Liu, Y.; Liu, Y.; Wen, X.; Zhang, K.; Ni, X.; Gao, N.; Fan, L.; Zhang, Z.; et al. In-situ generation of large numbers of genetic combinations for metabolic reprogramming via CRISPR-guided base editing. Nat. Commun. 2021, 12, 678. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F.; Cui, L.; Siouve, E.; Becavin, C.; Depardieu, F.; Bikard, D. Genome-wide CRISPR-dCas9 screens in E. coli identify essential genes and phage host factors. PLoS Gene. 2018, 14, e1007749. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Vigouroux, A.; Rousset, F.; Varet, H.; Khanna, V.; Bikard, D. A CRISPRi screen in E. coli reveals sequence-specific toxicity of dCas9. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Emmerstorfer-Augustin, A.; Moser, S.; Pichler, H. Screening for improved isoprenoid biosynthesis in microorganisms. J. Biotechnol. 2016, 235, 112–120. [Google Scholar] [CrossRef]
- Alper, H.; Jin, Y.-S.; Moxley, J.F.; Stephanopoulos, G. Identifying gene targets for the metabolic engineering of lycopene biosynthesis in Escherichia coli. Metab. Eng. 2005, 7, 155–164. [Google Scholar] [CrossRef]
- Park, C.-S.; Lee, S.-W.; Kim, Y.-S.; Kim, E.-J.; Sin, H.-S.; Oh, D.-K.; Kim, S.-W.; Um, S.-J. Utilization of the recombinant human β-carotene-15,15′-monooxygenase gene in Escherichia coli and mammalian cells. Biotechnol. Lett. 2008, 30, 735–741. [Google Scholar] [CrossRef]
- Armstrong, G.A. Eubacteria Show Their True Colors: Genetics of Carotenoid Pigment Biosynthesis from Microbes to Plants. J. Bacteriol. 1994, 176, 4795–4802. [Google Scholar] [CrossRef]
- Belviranlı, M.; Okudan, N. Well-Known Antioxidants and Newcomers in Sport Nutrition: Coenzyme Q10, Quercetin, Resveratrol, Pterostilbene, Pycnogenol and Astaxanthin. In Antioxidants in Sport Nutrition; Lamprecht, M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 79–102. ISBN 978-1-4665-6757-3. [Google Scholar]
- Blombach, B.; Seibold, G.M. Carbohydrate metabolism in Corynebacterium glutamicum and applications for the metabolic engineering of l-lysine production strains. Appl. Microbiol. Biotechnol. 2010, 86, 1313–1322. [Google Scholar] [CrossRef]
- Wendisch, V.F.; de Graaf, A.A.; Sahm, H.; Eikmanns, B.J. Quantitative determination of metabolic fluxes during coutilization of two carbon sources: Comparative analyses with Corynebacterium glutamicum during growth on acetate and/or glucose. J. Bacteriol. 2000, 182, 3088–3096. [Google Scholar] [CrossRef]
- Becker, J.; Wittmann, C. Bio-based production of chemicals, materials and fuels—Corynebacterium glutamicum as versatile cell factory. Curr. Opin. Biotechnol. 2012, 23, 631–640. [Google Scholar] [CrossRef]
- Kogure, T.; Inui, M. Recent advances in metabolic engineering of Corynebacterium glutamicum for bioproduction of value-added aromatic chemicals and natural products. Appl. Microbiol. Biotechnol. 2018, 102, 8685–8705. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, H. Microbial Production of Amino Acids in Japan. In History of Modern Biotechnology I; Fiechter, A., Ed.; Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2000; Volume 69, pp. 71–85. ISBN 978-3-540-67793-2. [Google Scholar]
- Wendisch, V.F. Metabolic engineering advances and prospects for amino acid production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef]
- Engels, V.; Wendisch, V.F. The DeoR-Type Regulator SugR Represses Expression of ptsG in Corynebacterium glutamicum. J. Bacteriol. 2007, 189, 2955–2966. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F. Genome-wide expression analysis in Corynebacterium glutamicum using DNA microarrays. J. Biotechnol. 2003, 104, 273–285. [Google Scholar] [CrossRef]
- Becker, J.; Klopprogge, C.; Zelder, O.; Heinzle, E.; Wittmann, C. Amplified expression of fructose 1,6-bisphosphatase in Corynebacterium glutamicum increases in vivo flux through the pentose phosphate pathway and lysine production on different carbon sources. AEM 2005, 71, 8587–8596. [Google Scholar] [CrossRef]
- Radmacher, E.; Eggeling, L. The three tricarboxylate synthase activities of Corynebacterium glutamicum and increase of l-lysine synthesis. Appl. Microbiol. Biotechnol. 2007, 76, 587–595. [Google Scholar] [CrossRef]
- van Ooyen, J.; Noack, S.; Bott, M.; Reth, A.; Eggeling, L. Improved l-lysine production with Corynebacterium glutamicum and systemic insight into citrate synthase flux and activity. Biotechnol. Bioeng. 2012, 109, 2070–2081. [Google Scholar] [CrossRef]
- Krubasik, P.; Takaichi, S.; Maoka, T.; Kobayashi, M.; Masamoto, K.; Sandmann, G. Detailed biosynthetic pathway to decaprenoxanthin diglucoside in Corynebacterium glutamicum and identification of novel intermediates. Arch. Microbiol. 2001, 176, 217–223. [Google Scholar] [CrossRef]
- Henke, N.; Heider, S.; Peters-Wendisch, P.; Wendisch, V. Production of the Marine Carotenoid Astaxanthin by Metabolically Engineered Corynebacterium glutamicum. Mar. Drugs 2016, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.E.; Peters-Wendisch, P.; Netzer, R.; Stafnes, M.; Brautaset, T.; Wendisch, V.F. Production and glucosylation of C50 and C40 carotenoids by metabolically engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2014, 98, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.E.; Peters-Wendisch, P.; Wendisch, V.F. Carotenoid biosynthesis and overproduction in Corynebacterium glutamicum. BMC Microbiol. 2012, 12, 198. [Google Scholar] [CrossRef]
- Kang, M.-K.; Eom, J.-H.; Kim, Y.; Um, Y.; Woo, H.M. Biosynthesis of pinene from glucose using metabolically-engineered Corynebacterium glutamicum. Biotechnol. Lett. 2014, 36, 2069–2077. [Google Scholar] [CrossRef]
- Ravikumar, S.; Woo, H.M.; Choi, J. Analysis of novel antioxidant sesquarterpenes (C35 terpenes) produced in recombinant Corynebacterium glutamicum. Appl. Biochem. Biotechnol. 2018, 186, 525–534. [Google Scholar] [CrossRef]
- Henke, N.; Wichmann, J.; Baier, T.; Frohwitter, J.; Lauersen, K.; Risse, J.; Peters-Wendisch, P.; Kruse, O.; Wendisch, V. Patchoulol Production with Metabolically Engineered Corynebacterium glutamicum. Genes 2018, 9, 219. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Park, J.; Woo, H.M. Overexpression of the key enzymes in the methylerythritol 4-phosphate pathway in Corynebacterium glutamicum for improving farnesyl diphosphate-derived terpene production. J. Agric. Food Chem. 2020, 68, 10780–10786. [Google Scholar] [CrossRef]
- Binder, D.; Frohwitter, J.; Mahr, R.; Bier, C.; Grünberger, A.; Loeschcke, A.; Peters-Wendisch, P.; Kohlheyer, D.; Pietruszka, J.; Frunzke, J.; et al. Light-Controlled Cell Factories: Employing Photocaged Isopropyl-β-d-Thiogalactopyranoside for Light-Mediated Optimization of Lac Promoter-Based Gene Expression and (+)-Valencene Biosynthesis in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2016, 82, 6141–6149. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, J.; Bathe, B.; Bartels, D.; Bischoff, N.; Bott, M.; Burkovski, A.; Dusch, N.; Eggeling, L.; Eikmanns, B.J.; Gaigalat, L.; et al. The complete Corynebacterium glutamicum ATCC 13032 genome sequence and its impact on the production of l-aspartate-derived amino acids and vitamins. J. Biotechnol. 2003, 104, 5–25. [Google Scholar] [CrossRef]
- Krubasik, P.; Kobayashi, M.; Sandmann, G. Expression and functional analysis of a gene cluster involved in the synthesis of decaprenoxanthin reveals the mechanisms for C50 carotenoid formation: Decaprenoxanthin formation. Eur. J. Biochem. 2001, 268, 3702–3708. [Google Scholar] [CrossRef]
- Krubasik, P.; Sandmann, G. A carotenogenic gene cluster from Brevibacterium linens with novel lycopene cyclase genes involved in the synthesis of aromatic carotenoids. Mol. Gen. Genet. 2000, 263, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Lim, J.H.; Kim, S.Y.; Im, D.-K.; Seok, J.Y.; Lee, S.-J.V.; Oh, M.-K.; Jung, G.Y. Precise precursor rebalancing for isoprenoids production by fine control of gapA expression in Escherichia coli. Metab. Eng. 2016, 38, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Farmer, W.R.; Liao, J.C. Precursor balancing for metabolic engineering of lycopene production in Escherichia coli. Biotechnol. Prog. 2001, 17, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, Q.; Sun, T.; Zhu, X.; Xu, H.; Tang, J.; Zhang, X.; Ma, Y. Engineering central metabolic modules of Escherichia coli for improving β-carotene production. Metab. Eng. 2013, 17, 42–50. [Google Scholar] [CrossRef]
- Zhao, X.; Shi, F.; Zhan, W. Overexpression of ZWF1 and POS5 improves carotenoid biosynthesis in recombinant Saccharomyces cerevisiae. Lett. Appl. Microbiol. 2015, 61, 354–360. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, F.; Yang, Y.; Jiang, Y.; Huo, Y.-X. Optimizing a CRISPR-Cpf1-based genome engineering system for Corynebacterium glutamicum. Microb. Cell Fact. 2019, 18, 60. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qian, F.; Yang, J.; Liu, Y.; Dong, F.; Xu, C.; Sun, B.; Chen, B.; Xu, X.; Li, Y.; et al. CRISPR-Cpf1 assisted genome editing of Corynebacterium glutamicum. Nat. Commun. 2017, 8, 15179. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Lu, Y.; Zheng, P.; Sun, J.; Ma, Y. Development of a CRISPR/Cas9 genome editing toolbox for Corynebacterium glutamicum. Microb. Cell Fact. 2017, 16, 205. [Google Scholar] [CrossRef]
- Cho, J.S.; Choi, K.R.; Prabowo, C.P.S.; Shin, J.H.; Yang, D.; Jang, J.; Lee, S.Y. CRISPR/Cas9-coupled recombineering for metabolic engineering of Corynebacterium glutamicum. Metab. Eng. 2017, 42, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Cleto, S.; Jensen, J.V.; Wendisch, V.F.; Lu, T.K. Corynebacterium glutamicum Metabolic Engineering with CRISPR interference (CRISPRi). ACS Synth. Biol. 2016, 5, 375–385. [Google Scholar] [CrossRef]
- Lee, S.S.; Shin, H.; Jo, S.; Lee, S.-M.; Um, Y.; Woo, H.M. Rapid identification of unknown carboxyl esterase activity in Corynebacterium glutamicum using RNA-guided CRISPR interference. Enzym. Microb. Technol. 2018, 114, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Lv, L.; Chen, J.-C.; Chen, G.-Q. Controlling microbial PHB synthesis via CRISPRi. Appl. Microbiol. Biotechnol. 2017, 101, 5861–5867. [Google Scholar] [CrossRef]
- Park, J.; Shin, H.; Lee, S.-M.; Um, Y.; Woo, H.M. RNA-guided single/double gene repressions in Corynebacterium glutamicum using an efficient CRISPR interference and its application to industrial strain. Microb. Cell Fact. 2018, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Woo, H.M. CRISPR interference-mediated metabolic engineering of Corynebacterium glutamicum for homo-butyrate production. Biotechnol. Bioeng. 2018, 115, 2067–2074. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, Z.-Q.; Liu, C.; Zheng, Y.-G. Application of CRISPRi in Corynebacterium glutamicum for shikimic acid production. Biotechnol. Lett. 2016, 38, 2153–2161. [Google Scholar] [CrossRef]
- Gauttam, R.; Seibold, G.M.; Mueller, P.; Weil, T.; Weiß, T.; Handrick, R.; Eikmanns, B.J. A simple dual-inducible CRISPR interference system for multiple gene targeting in Corynebacterium glutamicum. Plasmid 2019, 103, 25–35. [Google Scholar] [CrossRef]
- Baumgart, M.; Unthan, S.; Rückert, C.; Sivalingam, J.; Grünberger, A.; Kalinowski, J.; Bott, M.; Noack, S.; Frunzke, J. Construction of a prophage-free variant of Corynebacterium glutamicum ATCC 13032 for use as a platform strain for basic research and industrial biotechnology. Appl. Environ. Microbiol. 2013, 79, 6006–6015. [Google Scholar] [CrossRef]
- Eggeling, L.; Reyes, O. Experiments. In Handbook of Corynebacterium glutamicum; CRC Press: Boca Raton, FL, USA, 2005; pp. 3535–3566. [Google Scholar]
- Abe, S.; Takayama, K.-I.; Kinoshita, S. Taxonomical studies on glutamic acid-producing bacteria. J. Gen. Appl. Microbiol. 1967, 13, 279–301. [Google Scholar] [CrossRef]
- Schreiner, M.E.; Fiur, D.; Holátko, J.; Pátek, M.; Eikmanns, B.J.; Bacteriol, J. E1 Enzyme of the Pyruvate Dehydrogenase Complex in Corynebacterium glutamicum: Molecular Analysis of the Gene and Phylogenetic Aspects. J. Bacteriol. 2005, 187, 6005–6018. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.K.; Lindner, S.N.; Wendisch, V.F. Metabolic engineering of an ATP-neutral Embden-Meyerhof-Parnas pathway in Corynebacterium glutamicum: Growth restoration by an adaptive point mutation in NADH dehydrogenase. Appl. Environ. Microbiol. 2015, 81, 1996–2005. [Google Scholar] [CrossRef] [PubMed]
- Lindner, S.N.; Petrov, D.P.; Hagmann, C.T.; Henrich, A.; Krämer, R.; Eikmanns, B.J.; Wendisch, V.F.; Seibold, G.M. Phosphotransferase system-mediated glucose uptake is repressed in phosphoglucoisomerase-deficient Corynebacterium glutamicum strains. Appl. Environ. Microbiol. 2013, 79, 2588–2595. [Google Scholar] [CrossRef]
- Henke, N.A.; Heider, S.A.E.; Hannibal, S.; Wendisch, V.F.; Peters-Wendisch, P. Isoprenoid pyrophosphate-dependent transcriptional regulation of carotenogenesis in Corynebacterium glutamicum. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbachb, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Schultenkämper, K.; Brito, L.F.; López, M.G.; Brautaset, T.; Wendisch, V.F. Establishment and application of CRISPR interference to affect sporulation, hydrogen peroxide detoxification, and mannitol catabolism in the methylotrophic thermophile Bacillus methanolicus. Appl. Microbiol. Biotechnol. 2019, 103, 5879–5889. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Blin, K.; Pedersen, L.E.; Weber, T.; Lee, S.Y. CRISPy-Web: An online resource to design sgRNAs for CRISPR applications. Synth. Syst. Biotechnol. 2016, 1, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Eggeling, L.; Bott, M. Handbook of Corynebacterium glutamicum, 1st ed.; CRC Press: Bota Raton, FL, USA, 2004. [Google Scholar]
- van der Rest, M.E.; Lange, C.; Molenaar, D. A heat shock following electroporation induces highly efficient transformation of Corynebacterium glutamicum with xenogeneic plasmid DNA. Appl. Microbiol. Biotechnol. 1999, 52, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Dollinger, G.; Walsh, P.S.; Griffith, R. Simultaneous amplification and detection of specific DNA sequences. Nat. Biotechnol. 1992, 10, 413–417. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Ajikumar, P.K.; Xiao, W.-H.; Tyo, K.E.J.; Wang, Y.; Simeon, F.; Leonard, E.; Mucha, O.; Phon, T.H.; Pfeifer, B.; Stephanopoulos, G. Isoprenoid pathway optimization for taxol precursor overproduction in Escherichia coli. Science 2010, 330, 70–74. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Peters-Wendisch, P.; Beekwilder, J.; Wendisch, V.F. IdsA is the major geranylgeranyl pyrophosphate synthase involved in carotenogenesis in Corynebacterium glutamicum. FEBS J. 2014, 281, 4906–4920. [Google Scholar] [CrossRef]
- Moise, A.R.; Al-Babili, S.; Wurtzel, E.T. Mechanistic aspects of carotenoid biosynthesis. Chem. Rev. 2014, 114, 164–193. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, H.; Henke, N.A.; Heider, S.A.E.; Wendisch, V.F. Overexpression of the primary sigma factor gene SigA improved carotenoid production by Corynebacterium glutamicum: Application to production of β-carotene and the non-native linear C50 carotenoid bisanhydrobacterioruberin. Metab. Eng. Comm. 2017, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Engels, V.; Lindner, S.N.; Wendisch, V.F. The global repressor SugR controls expression of genes of glycolysis and of the l-lactate dehydrogenase LdhA in Corynebacterium glutamicum. J. Bacteriol. 2008, 190, 8033–8044. [Google Scholar] [CrossRef]
- Mo, X.-H.; Zhang, H.; Wang, T.-M.; Zhang, C.; Zhang, C.; Xing, X.-H.; Yang, S. Establishment of CRISPR interference in Methylorubrum extorquens and application of rapidly mining a new phytoene desaturase involved in carotenoid biosynthesis. Appl. Microbiol. Biotechnol. 2020, 104, 4515–4532. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Liu, J.-Z. Enhanced astaxanthin production in Escherichia coli via morphology and oxidative stress engineering. J. Agric. Food Chem. 2019, 67, 11703–11709. [Google Scholar] [CrossRef]
- Wendisch, V.F.; Bott, M.; Kalinowski, J.; Oldiges, M.; Wiechert, W. Emerging Corynebacterium glutamicum Systems Biology. J. Biotechnol. 2006, 124, 74–92. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, K.; Teramoto, H.; Inui, M.; Yukawa, H. Molecular mechanism of SugR-mediated sugar-dependent expression of the ldhA gene encoding l-lactate dehydrogenase in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2009, 83, 315–327. [Google Scholar] [CrossRef]
- Toyoda, K.; Teramoto, H.; Inui, M.; Yukawa, H. Expression of the gapA gene encoding glyceraldehyde-3-phosphate dehydrogenase of Corynebacterium glutamicum is regulated by the global regulator SugR. Appl. Microbiol. Biotechnol. 2008, 81, 291–301. [Google Scholar] [CrossRef]
- Tanaka, Y.; Teramoto, H.; Inui, M.; Yukawa, H. Regulation of expression of general components of the phosphoenolpyruvate: Carbohydrate phosphotransferase system (PTS) by the global regulator SugR in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2008, 78, 309–318. [Google Scholar] [CrossRef]
- Gaigalat, L.; Schlüter, J.-P.; Hartmann, M.; Mormann, S.; Tauch, A.; Pühler, A.; Kalinowski, J. The DeoR-type transcriptional regulator SugR acts as a repressor for genes encoding the phosphoenolpyruvate:sugar phosphotransferase system (PTS) in Corynebacterium glutamicum. BMC Mol. Biol. 2007, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- Donati, S.; Kuntz, M.; Pahl, V.; Farke, N.; Beuter, D.; Glatter, T.; Gomes-Filho, J.V.; Randau, L.; Wang, C.-Y.; Link, H. Multi-omics analysis of CRISPRi-knockdowns identifies mechanisms that buffer decreases of enzymes in E. coli metabolism. Cell Syst. 2021, 12, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Alper, H.S.; Avalos, J.L. Metabolic Pathway Engineering. Synth. Syst. Biotechnol. 2018, 3, 1–2. [Google Scholar] [CrossRef]
- Blazeck, J.; Alper, H.S. Promoter engineering: Recent advances in controlling transcription at the most fundamental level. Biotechnol. J. 2013, 8, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, S.Y.; Kim, T.Y.; Kim, H.U. Application of systems biology for bioprocess development. Trends Biotechnol. 2008, 26, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Sander, T.; Wang, C.Y.; Glatter, T.; Link, H. CRISPRi-based downregulation of transcriptional feedback improves growth and metabolism of arginine overproducing E. coli. ACS Synth. Biol. 2019, 8, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hou, Z.; Ma, Q.; Mo, X.; Sun, Q.; Tan, M.; Xia, L.; Lin, G.; Yang, M.; Zhang, Y.; et al. CRISPRi-based dynamic control of carbon flow for efficient N -acetyl glucosamine production and its metabolomic effects in Escherichia coli. J. Agric. Food Chem. 2020, 68, 3203–3213. [Google Scholar] [CrossRef]
- Marx, A.; de Graaf, A.A.; Wiechert, W.; Eggeling, L.; Sahm, H. Determination of the fluxes in the central metabolism of Corynebacterium glutamicum by nuclear magnetic resonance spectroscopy combined with metabolite balancing. Biotechnol. Bioeng. 1996, 49, 111–129. [Google Scholar] [CrossRef]
- Marx, A.; Striegel, K.; de Graaf, A.A.; Sahm, H.; Eggeling, L. Response of the central metabolism of Corynebacterium glutamicum to different flux burdens. Biotechnol. Bioeng. 1997, 56, 168–180. [Google Scholar] [CrossRef]
- Kiefer, P.; Heinzle, E.; Zelder, O.; Wittmann, C. Comparative metabolic flux analysis of lysine-producing Corynebacterium glutamicum cultured on glucose or fructose. AEM 2004, 70, 229–239. [Google Scholar] [CrossRef]
- Georgi, T.; Rittmann, D.; Wendisch, V.F. Lysine and glutamate production by Corynebacterium glutamicum on glucose, fructose and sucrose: Roles of malic enzyme and fructose-1,6-bisphosphatase. Metab. Eng. 2005, 7, 291–301. [Google Scholar] [CrossRef]
- Marx, A.; Hans, S.; Möckel, B.; Bathe, B.; de Graaf, A.A. Metabolic phenotype of phosphoglucose isomerase mutants of Corynebacterium glutamicum. J. Biotechnol. 2003, 104, 185–197. [Google Scholar] [CrossRef]
- Goyal, A.; Myacheva, K.; Groß, M.; Klingenberg, M.; Duran Arqué, B.; Diederichs, S. Challenges of CRISPR/Cas9 applications for long non-coding RNA genes. Nucleic Acids Res. 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Characteristics | Reference |
|---|---|---|
| Corynebacterium glutamicum strains | ||
| Wild type (WT) | Wild-type ATCC 13032 | [59] |
| WT ΔaceE | aceE (cg2466) deletion mutant of WT | [60] |
| WT ΔgapA | gapA (cg1791) deletion mutant of WT | [61] |
| WT Δpgi | pgi (cg0973) deletion mutant of WT | [62] |
| WT ΔsdhCAB | sdhCAB (cg0445/0447/0448) deletion mutant of WT | This work |
| WT ΔsugR | sugR (cg2115) deletion mutant of WT | [25] |
| MB001 | Prophage-cured, genome-reduced WT | [57] |
| MB001 ΔcrtR | crtR (cg0725) deletion mutant of MB001 | [63] |
| E. coli strains | ||
| E. coli DH5α | F-thi−1 endA1 hsdr17(r-, m-) supE44 ΔlacU169 (Φ80lacZΔM15) recA1 gyrA96 | [64] |
| Plasmids | ||
| pK19mobsacB | Mobilizable E. coli vector used for the construction of insertion and deletion mutants of C. glutamicum (oriV, sacB, lacZα); KanR | [65] |
| pK19mobsacB-ΔsdhCAB | pK19mobsacB for the deletion of sdhCAB (cg0445/0447/0448); KanR | This work |
| pRG_dCas9 | E. coli/C. glutamicum shuttle clustered regularly interspaced short palindromic repeats interference (CRISPRi) vector, anhydrotetracycline (aTc)- and IPTG-inducible; CmR | [56] |
| piCas | E. coli/B. methanolicus shuttle CRISPRi vector, mannitol-inducible; CmR | [66] |
| pS_dCas9 | pRG_dCas9 carrying the dCas9 handle followed by the terminator from S. pyogenes; CmR | This work |
| pS_dCas9_aceA | pS_dCas9 plasmid carrying the aceA (cg2560) sgRNA | This work |
| pS_dCas9_aceB | pS_dCas9 plasmid carrying the aceB (cg2559) sgRNA | This work |
| pS_dCas9_aceE | pS_dCas9 plasmid carrying the aceE (cg2466) sgRNA | This work |
| pS_dCas9_ackA | pS_dCas9 plasmid carrying the ackA (cg3047) sgRNA | This work |
| pS_dCas9_acn | pS_dCas9 plasmid carrying the acn (cg1737) sgRNA | This work |
| pS_dCas9_crtB | pS_dCas9 plasmid carrying the crtB (cg0721) sgRNA | This work |
| pS_dCas9_crtE | pS_dCas9 plasmid carrying the crtE (cg0723) sgRNA | This work |
| pS_dCas9_crtEb | pS_dCas9 plasmid carrying the crtEb (cg0717) sgRNA | This work |
| pS_dCas9_crtI | pS_dCas9 plasmid carrying the crtI (cg0720) sgRNA | This work |
| pS_dCas9_crtR | pS_dCas9 plasmid carrying the crtR (cg0725) sgRNA | This work |
| pS_dCas9_crtX | pS_dCas9 plasmid carrying the crtX (cg0730) sgRNA | This work |
| pS_dCas9_crtYe | pS_dCas9 plasmid carrying the crtYe (cg0719) sgRNA | This work |
| pS_dCas9_deoC | pS_dCas9 plasmid carrying the deoC (cg0458) sgRNA | This work |
| pS_dCas9_dxr | pS_dCas9 plasmid carrying the dxr (cg2208) sgRNA | This work |
| pS_dCas9_dxs | pS_dCas9 plasmid carrying the dxs (cg2083) sgRNA | This work |
| pS_dCas9_eno | pS_dCas9 plasmid carrying the eno (cg1111) sgRNA | This work |
| pS_dCas9_fba | pS_dCas9 plasmid carrying the fba (cg3068) sgRNA | This work |
| pS_dCas9_fbp | pS_dCas9 plasmid carrying the fbp (cg1157) sgRNA | This work |
| pS_dCas9_fixB | pS_dCas9 plasmid carrying the fixB (cg1386) sgRNA | This work |
| pS_dCas9_fum | pS_dCas9 plasmid carrying the fum (cg1145) sgRNA | This work |
| pS_dCas9_gapA | pS_dCas9 plasmid carrying the gapA (cg1791) sgRNA | This work |
| pS_dCas9_gltA | pS_dCas9 plasmid carrying the gltA (cg0949) sgRNA | This work |
| pS_dCas9_glxR | pS_dCas9 plasmid carrying the glxR (cg0350) sgRNA | This work |
| pS_dCas9_gnd | pS_dCas9 plasmid carrying the gnd (cg1643) sgRNA | This work |
| pS_dCas9_icd | pS_dCas9 plasmid carrying the icd (cg0766) sgRNA | This work |
| pS_dCas9_idsA | pS_dCas9 plasmid carrying the idsA (cg2384) sgRNA | This work |
| pS_dCas9_IIdD | pS_dCas9 plasmid carrying the IIdD (cg3227) sgRNA | This work |
| pS_dCas9_ispE | pS_dCas9 plasmid carrying the ispE (cg1039) sgRNA | This work |
| pS_dCas9_ispF | pS_dCas9 plasmid carrying the ispF (cg2944) sgRNA | This work |
| pS_dCas9_ispG | pS_dCas9 plasmid carrying the ispG (cg2206) sgRNA | This work |
| pS_dCas9_ispH | pS_dCas9 plasmid carrying the ispH (cg1164) sgRNA | This work |
| pS_dCas9_ldh | pS_dCas9 plasmid carrying the ldh (cg3219) sgRNA | This work |
| pS_dCas9_malE | pS_dCas9 plasmid carrying the malE (cg3335) sgRNA | This work |
| pS_dCas9_mdh | pS_dCas9 plasmid carrying the mdh (cg2613) sgRNA | This work |
| pS_dCas9_mmpl | pS_dCas9 plasmid carrying the mmpl (cg0722) sgRNA | This work |
| pS_dCas9_odhA | pS_dCas9 plasmid carrying the odhA (cg1280) sgRNA | This work |
| pS_dCas9_odx | pS_dCas9 plasmid carrying the odx (cg1458) sgRNA | This work |
| pS_dCas9_opcA | pS_dCas9 plasmid carrying the opcA (cg1779) sgRNA | This work |
| pS_dCas9_pck | pS_dCas9 plasmid carrying the pck (cg3169) sgRNA | This work |
| pS_dCas9_pfkA | pS_dCas9 plasmid carrying the pfkA (cg1409) sgRNA | This work |
| pS_dCas9_pgi | pS_dCas9 plasmid carrying the pgi (cg0973) sgRNA | This work |
| pS_dCas9_pgk | pS_dCas9 plasmid carrying the pgk (cg1790) sgRNA | This work |
| pS_dCas9_pgl | pS_dCas9 plasmid carrying the pgl (cg1780) sgRNA | This work |
| pS_dCas9_pgm | pS_dCas9 plasmid carrying the pgm (cg2800) sgRNA | This work |
| pS_dCas9_ppc | pS_dCas9 plasmid carrying the ppc (cg1787) sgRNA | This work |
| pS_dCas9_ppsA | pS_dCas9 plasmid carrying the ppsA (cg0644) sgRNA | This work |
| pS_dCas9_pqo | pS_dCas9 plasmid carrying the pqo (cg2891) sgRNA | This work |
| pS_dCas9_pta | pS_dCas9 plasmid carrying the pta (cg3048) sgRNA | This work |
| pS_dCas9_ptsG | pS_dCas9 plasmid carrying the ptsG (cg1537) sgRNA | This work |
| pS_dCas9_pyc | pS_dCas9 plasmid carrying the pyc (cg0791) sgRNA | This work |
| pS_dCas9_pyk | pS_dCas9 plasmid carrying the pyk (cg2291) sgRNA | This work |
| pS_dCas9_ramB | pS_dCas9 plasmid carrying the ramB (cg0444) sgRNA | This work |
| pS_dCas9_rpe | pS_dCas9 plasmid carrying the rpe (cg1801) sgRNA | This work |
| pS_dCas9_rpi | pS_dCas9 plasmid carrying the rpi (cg2658) sgRNA | This work |
| pS_dCas9_rsdA | pS_dCas9 plasmid carrying the rsdA (cg0697) sgRNA | This work |
| pS_dCas9_rshA | pS_dCas9 plasmid carrying the rshA (cg0877) sgRNA | This work |
| pS_dCas9_sdhA | pS_dCas9 plasmid carrying the sdhA (cg0446) sgRNA | This work |
| pS_dCas9_sdhB | pS_dCas9 plasmid carrying the sdhB (cg0447) sgRNA | This work |
| pS_dCas9_sdhCD | pS_dCas9 plasmid carrying the sdhCD (cg0445) sgRNA | This work |
| pS_dCas9_sigA | pS_dCas9 plasmid carrying the sigA (cg2092) sgRNA | This work |
| pS_dCas9_sigB | pS_dCas9 plasmid carrying the sigB (cg2102) sgRNA | This work |
| pS_dCas9_sigC | pS_dCas9 plasmid carrying the sigC (cg0309) sgRNA | This work |
| pS_dCas9_sigD | pS_dCas9 plasmid carrying the sigD (cg0696) sgRNA | This work |
| pS_dCas9_sigE | pS_dCas9 plasmid carrying the sigE (cg1271) sgRNA | This work |
| pS_dCas9_sigH | pS_dCas9 plasmid carrying the sigH (cg0876) sgRNA | This work |
| pS_dCas9_sigM | pS_dCas9 plasmid carrying the sigM (cg3420) sgRNA | This work |
| pS_dCas9_sucC | pS_dCas9 plasmid carrying the sucC (cg2837) sgRNA | This work |
| pS_dCas9_sucD | pS_dCas9 plasmid carrying the sucD (cg2836) sgRNA | This work |
| pS_dCas9_sugR | pS_dCas9 plasmid carrying the sugR (cg2115) sgRNA | This work |
| pS_dCas9_tal | pS_dCas9 plasmid carrying the tal (cg1776) sgRNA | This work |
| pS_dCas9_thiE | pS_dCas9 plasmid carrying the thiE (cg2236) sgRNA | This work |
| pS_dCas9_tkt | pS_dCas9 plasmid carrying the tkt (cg1774) sgRNA | This work |
| pS_dCas9_tpi | pS_dCas9 plasmid carrying the tpi (cg1789) sgRNA | This work |
| pS_dCas9_zwf | pS_dCas9 plasmid carrying the zwf (cg1778) sgRNA | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Göttl, V.L.; Schmitt, I.; Braun, K.; Peters-Wendisch, P.; Wendisch, V.F.; Henke, N.A. CRISPRi-Library-Guided Target Identification for Engineering Carotenoid Production by Corynebacterium glutamicum. Microorganisms 2021, 9, 670. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040670
Göttl VL, Schmitt I, Braun K, Peters-Wendisch P, Wendisch VF, Henke NA. CRISPRi-Library-Guided Target Identification for Engineering Carotenoid Production by Corynebacterium glutamicum. Microorganisms. 2021; 9(4):670. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040670
Chicago/Turabian StyleGöttl, Vanessa L., Ina Schmitt, Kristina Braun, Petra Peters-Wendisch, Volker F. Wendisch, and Nadja A. Henke. 2021. "CRISPRi-Library-Guided Target Identification for Engineering Carotenoid Production by Corynebacterium glutamicum" Microorganisms 9, no. 4: 670. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040670