
Identification of a Toxin–Antitoxin System That Contributes to Persister Formation by Reducing NAD in Pseudomonas aeruginosa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Construction of Plasmids and Mutation Strains
2.3. Bacteria Killing Assay
2.4. RNA Isolation and Quantitative Real-Time PCR
2.5. Transcriptional Reporter Assay
2.6. Expression and Purification of the PA14_51020 Protein
2.7. Electrophoretic Mobility Shift Assay (EMSA)
2.8. Pull Down Assay
3. Results
3.1. Identification of Novel Persister Formation Related Genes
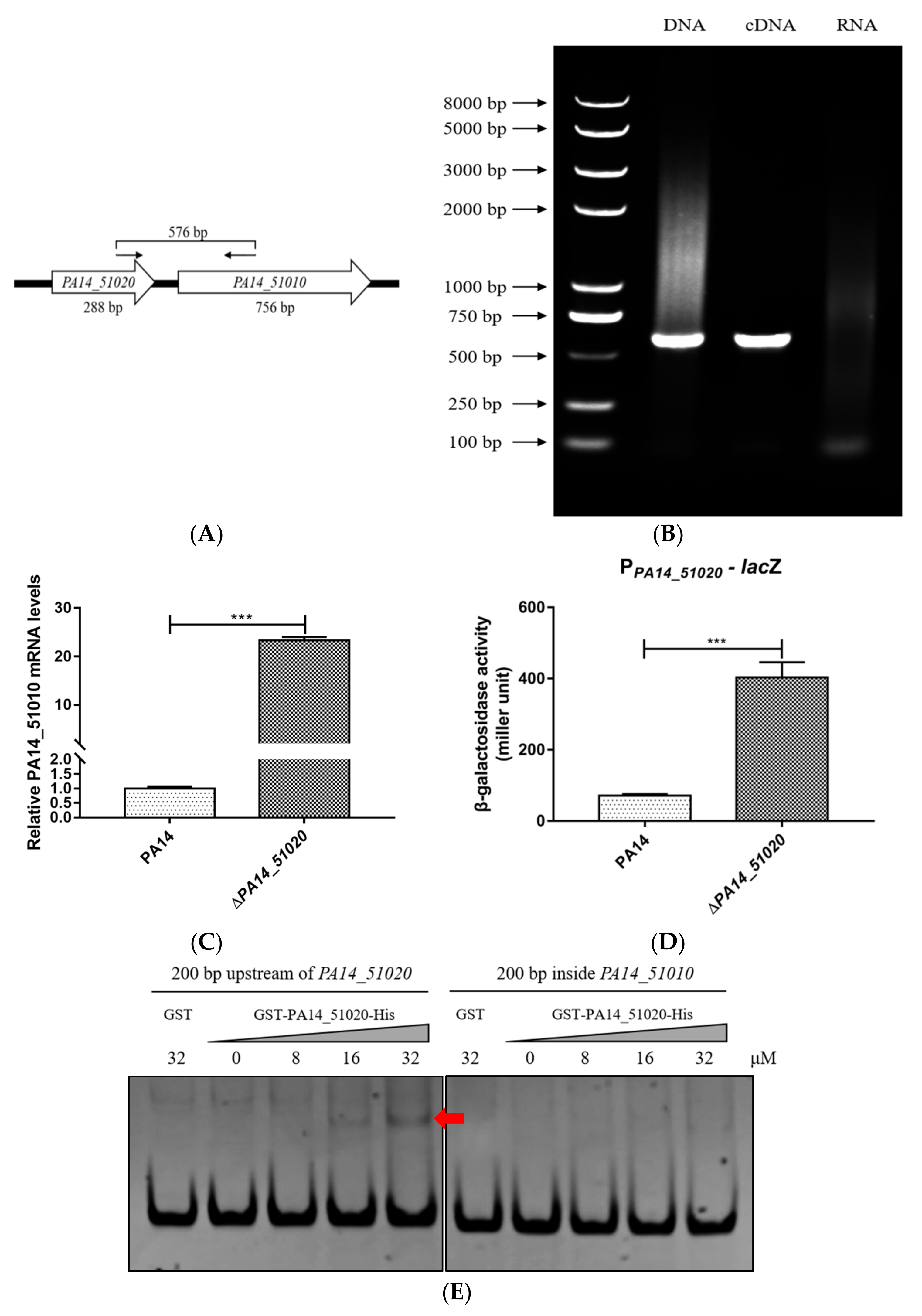
3.2. PA14_51020 Regulates the Operon of PA14_51020 and PA14_51010
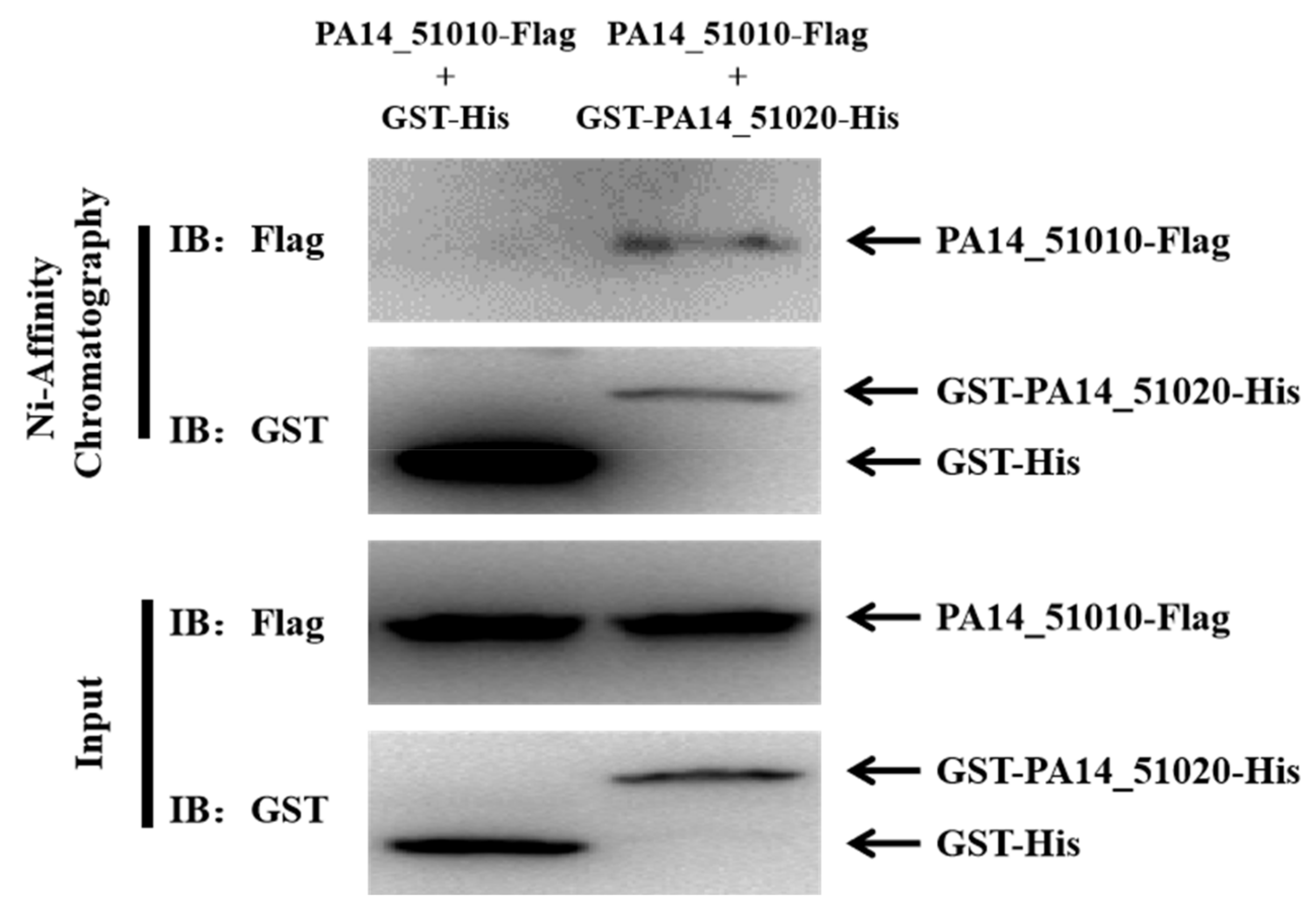
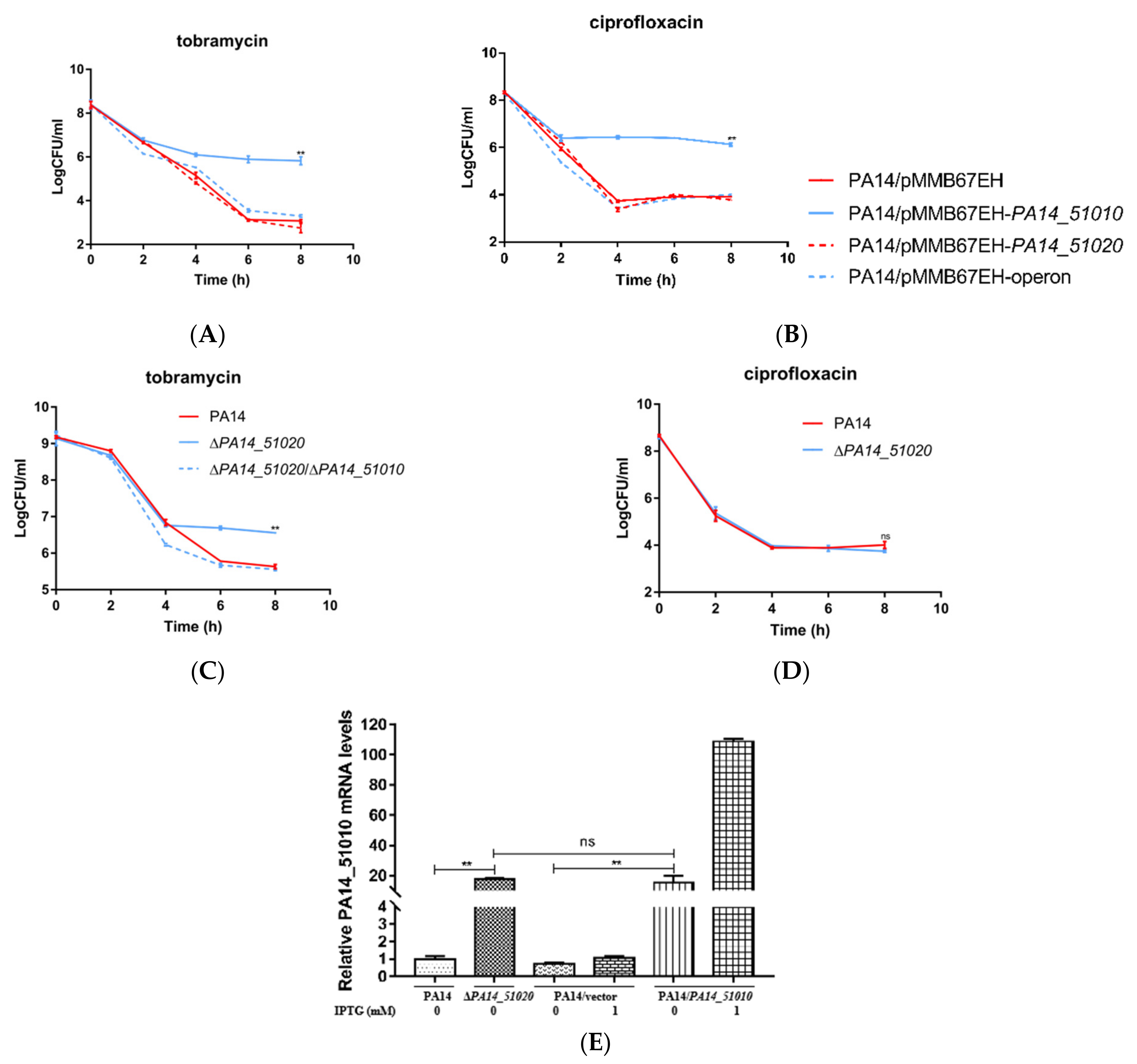
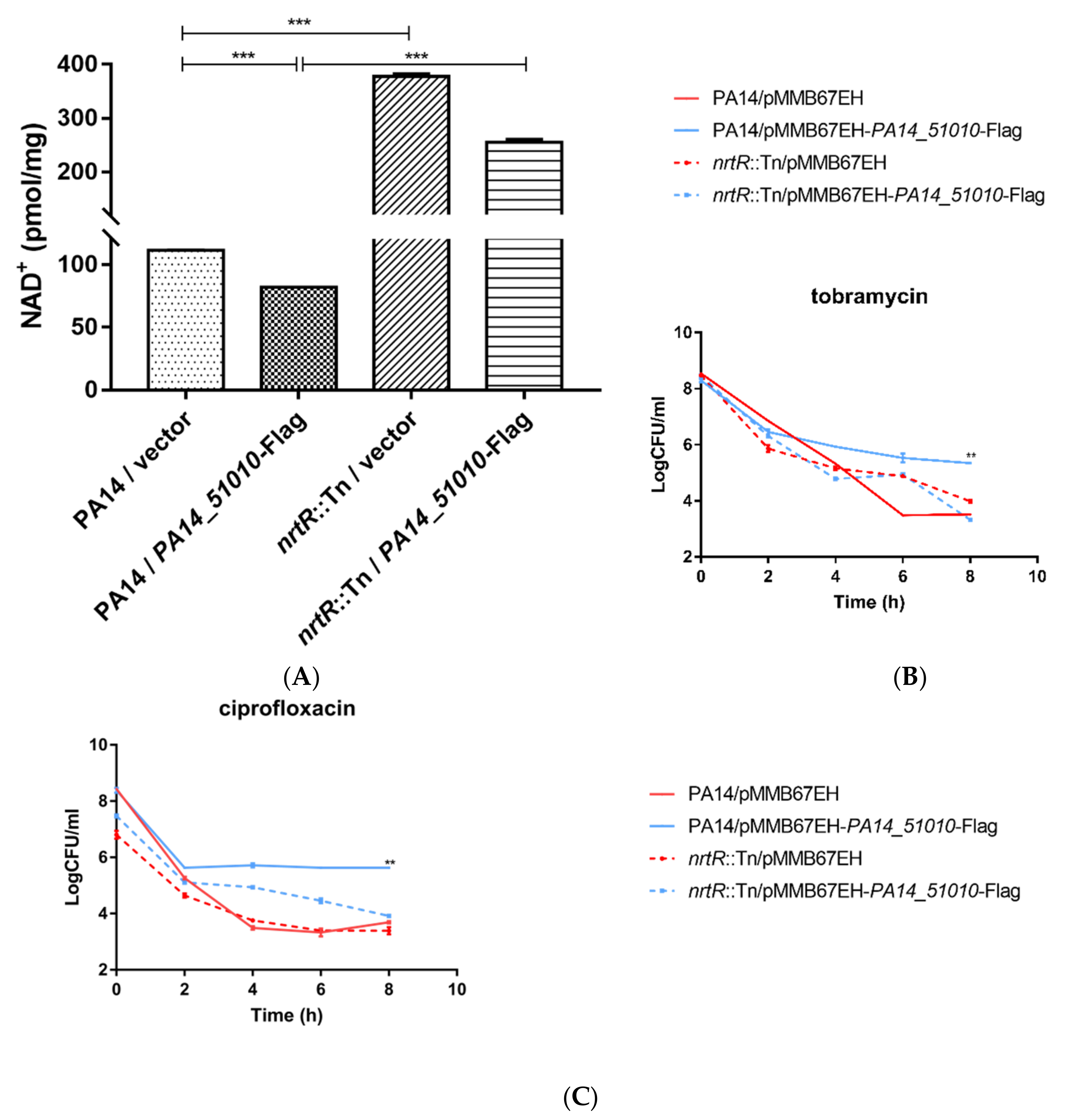
3.3. PA14_51020 and PA14_51010 Regulates Persister Formation
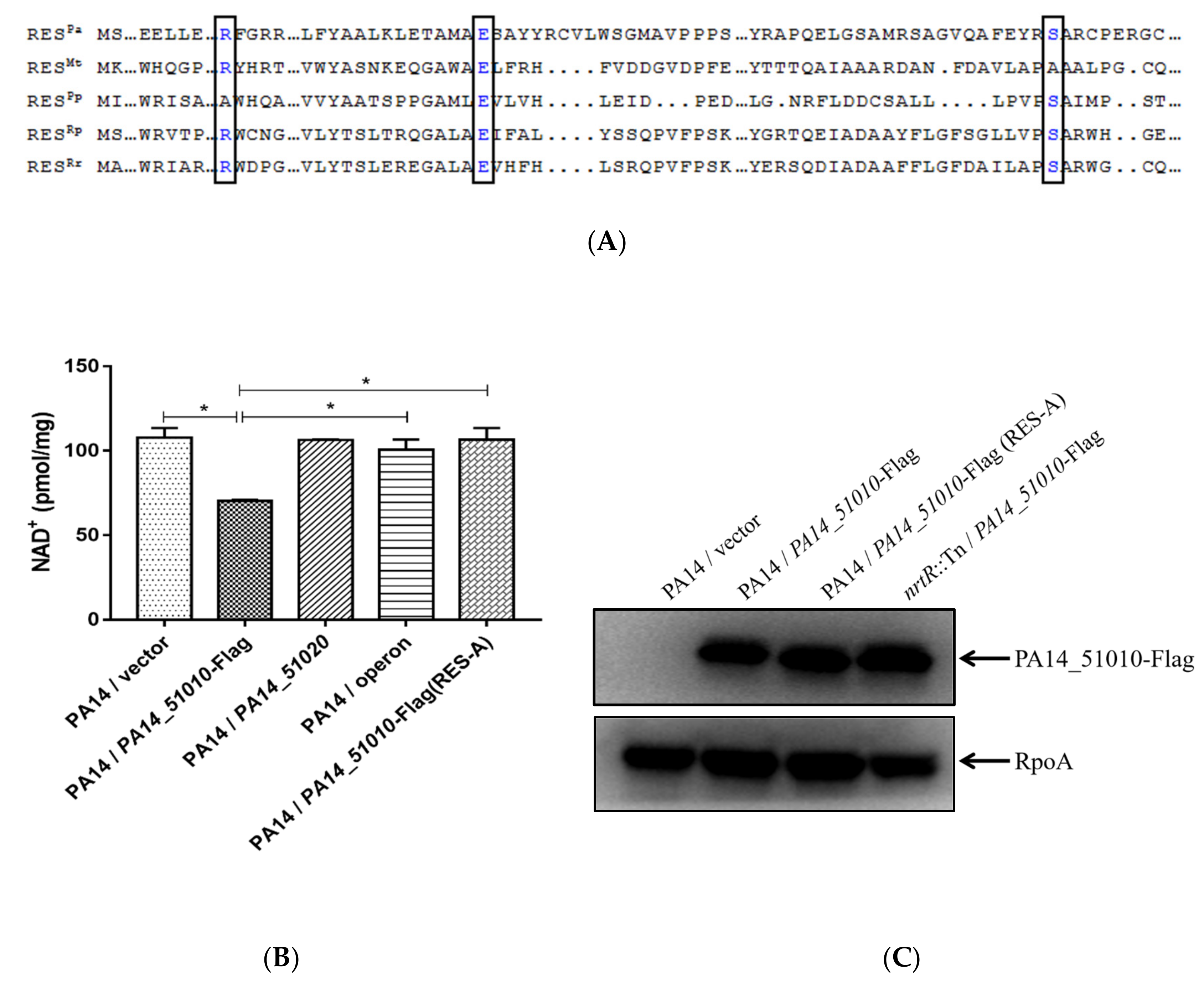
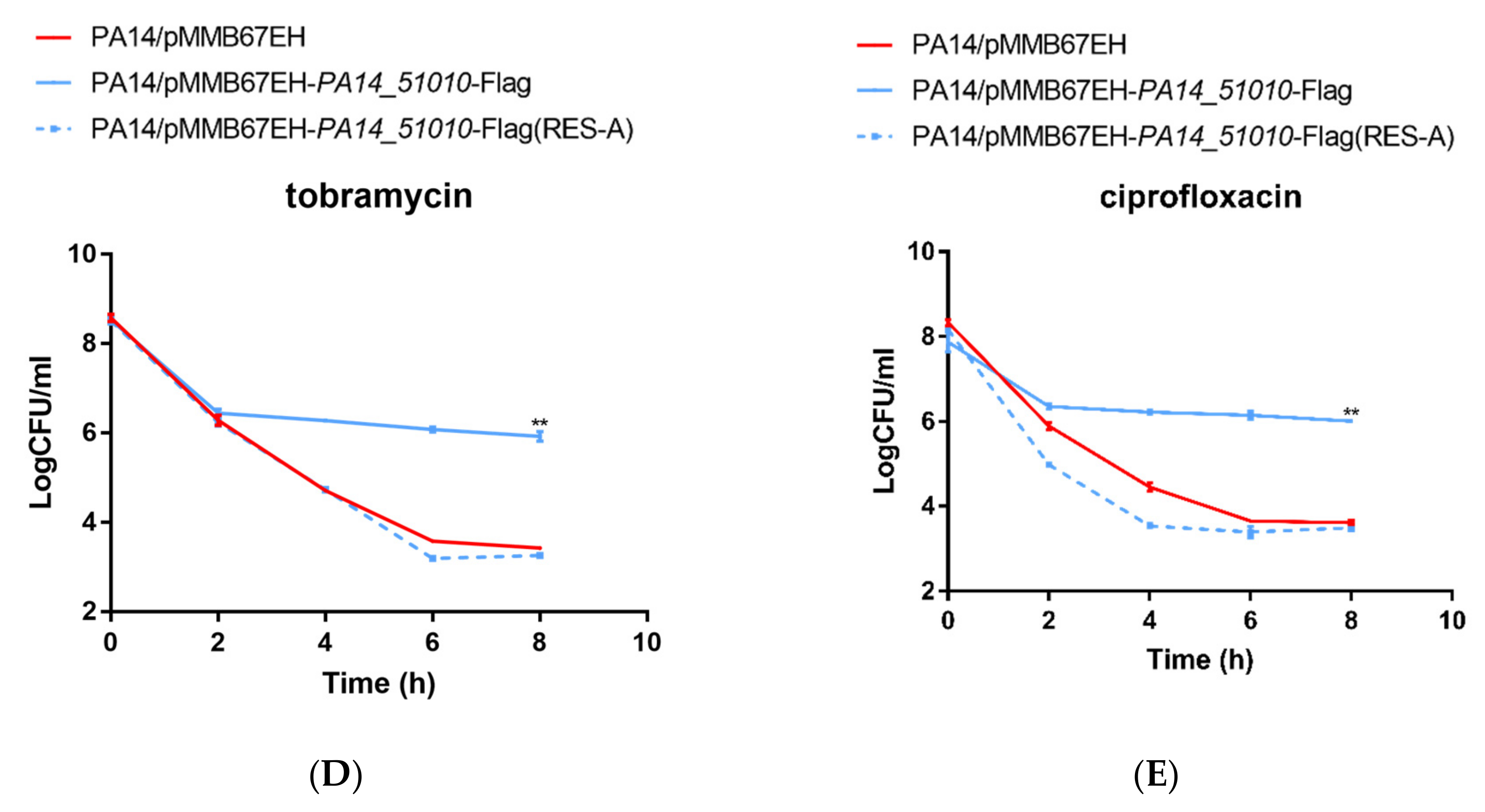
3.4. PA14_51010 Promotes Persister Formation by Reducing the Intracellular Level of NAD+
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wood, T.K. Combatting bacterial persister cells. Biotechnol. Bioeng. 2015, 113, 476–483. [Google Scholar] [CrossRef]
- Paranjape, S.S.; Shashidhar, R. Comparison of Starvation-Induced Persister Cells with Antibiotic-Induced Persister Cells. Curr. Microbiol. 2019, 76, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Genet. 2017, 15, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Vallette, F.M.; Olivier, C.; Lézot, F.; Oliver, L.; Cochonneau, D.; Lalier, L.; Cartron, P.-F.; Heymann, D. Dormant, quiescent, tolerant and persister cells: Four synonyms for the same target in cancer. Biochem. Pharmacol. 2019, 162, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial persistence as a phenotypic switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, N.R.; Lobritz, M.A.; Collins, J.J. Microbial persistence and the road to drug resistance. Cell Host Microbe 2013, 13, 632–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. Science 2017, 355, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, J.; Swaminath, S.; Nair, R.R.; Jakkala, K.; Pradhan, A.; Ajitkumar, P. De novo emergence of genetically resistant mutants of mycobacterium tuberculosis from the persistence phase cells formed against antituberculosis drugs in vitro. Antimicrob. Agents Chemother. 2017, 61, e01343-16. [Google Scholar] [CrossRef] [Green Version]
- Page, R.; Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef]
- Kim, J.-S.; Heo, P.; Yang, T.-J.; Lee, K.-S.; Cho, D.-H.; Kim, B.T.; Suh, J.-H.; Lim, H.-J.; Shin, D.; Kim, S.-K.; et al. Selective killing of bacterial persisters by a single chemical compound without affecting normal antibiotic-sensitive cells. Antimicrob. Agents Chemother. 2011, 55, 5380–5383. [Google Scholar] [CrossRef] [Green Version]
- Svenningsen, M.S.; Veress, A.; Harms, A.; Mitarai, N.; Semsey, S. Birth and Resuscitation of (p)ppGpp Induced Antibiotic Tolerant Persister Cells. Science Rep. 2019, 9, 6056. [Google Scholar] [CrossRef]
- Wen, Y.; Behiels, E.; Devreese, B. Toxin-Antitoxin systems: Their role in persistence, biofilm formation, and pathogenicity. Pathog. Dis. 2014, 70, 240–249. [Google Scholar] [CrossRef]
- Schuster, C.F.; Bertram, R. Toxin-antitoxin systems are ubiquitous and versatile modulators of prokaryotic cell fate. FEMS Microbiol. Lett. 2013, 340, 73–85. [Google Scholar] [CrossRef]
- García-Contreras, R.; Zhang, X.-S.; Kim, Y.; Wood, T.K. Protein translation and cell death: The role of rare tRNAs in biofilm formation and in activating dormant phage killer genes. PLoS ONE 2008, 3, e2394. [Google Scholar] [CrossRef]
- Marimon, O.; Teixeira, J.M.C.; Cordeiro, T.N.; Soo, V.W.C.; Wood, T.L.; Mayzel, M.; Amata, I.; García, J.; Morera, A.; Gay, M.; et al. An oxygen-sensitive toxin–antitoxin system. Nat. Commun. 2016, 7, 13634. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Gao, X.; Zhu, K.; Yin, H.; Mao, X.; Wojdyla, J.A.; Qin, B.; Huang, H.; Wang, M.; Sun, Y.-C.; et al. Characterization of a toxin-antitoxin system in Mycobacterium tuberculosis suggests neutralization by phosphorylation as the antitoxicity mechanism. Commun. Biol. 2020, 3, 216. [Google Scholar] [CrossRef]
- Yao, J.; Zhen, X.; Tang, K.; Liu, T.; Xu, X.; Chen, Z.; Guo, Y.; Liu, X.; Wood, T.K.; Ouyang, S.; et al. Novel polyadenylylation-dependent neutralization mechanism of the HEPN/MNT toxin/antitoxin system. Nucleic Acids Res. 2020, 48, 11054–11067. [Google Scholar] [CrossRef]
- Wang, X.; Yao, J.; Sun, Y.-C.; Wood, T.K. Type VII toxin/antitoxin classification system for antitoxins that enzymatically neutralize toxins. Trends Microbiol. 2020. [Google Scholar] [CrossRef]
- Syed, M.A.; Lévesque, C.M. Chromosomal bacterial type II toxin–antitoxin systems. Can. J. Microbiol. 2012, 58, 553–562. [Google Scholar] [CrossRef]
- Sala, A.; Bordes, P.; Genevaux, P. Multiple toxin-antitoxin systems in Mycobacterium tuberculosis. Toxins 2014, 6, 1002–1020. [Google Scholar] [CrossRef] [Green Version]
- Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Type II Toxin-Antitoxin Systems: Evolution and revolutions. J. Bacteriol. 2020, 202, e00763-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A paradigm for adaptation, survival, and persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Viducic, D.; Ono, T.; Murakami, K.; Susilowati, H.; Kayama, S.; Hirota, K.; Miyake, Y. Functional Analysis ofspoT, relA and dksA Genes on quinolone tolerance in Pseudomonas aeruginosa under nongrowing condition. Microbiol. Immunol. 2006, 50, 349–357. [Google Scholar] [CrossRef]
- Soares, A.; Alexandre, K.; Etienne, M. Tolerance and persistence of Pseudomonas aeruginosa in biofilms exposed to antibiotics: Molecular mechanisms, antibiotic strategies and therapeutic perspectives. Front. Microbiol. 2020, 11, 2057. [Google Scholar] [CrossRef]
- Muthuramalingam, M.; White, J.C.; Murphy, T.; Ames, J.R.; Bourne, C.R. The toxin from a ParDE toxin-antitoxin system found in Pseudomonas aeruginosa offers protection to cells challenged with anti-gyrase antibiotics. Mol. Microbiol. 2018, 111, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Shen, M.; Lu, S.; Le, S.; Tan, Y.; Wang, J.; Zhao, X.; Shen, W.; Guo, K.; Yang, Y.; et al. Identification and characterization of the HicAB Toxin-Antitoxin System in the opportunistic pathogen Pseudomonas aeruginosa. Toxins 2016, 8, 113. [Google Scholar] [CrossRef] [Green Version]
- Coskun, U.S.S. Effect of mazEF, higBA and relBE Toxin-Antitoxin Systems on antibiotic resistance in Pseudomonas aeruginosa and Staphylococcus isolates. Malawi Med. J. 2018, 30, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.L.; Wood, T.K. The HigB/HigA toxin/antitoxin system of Pseudomonas aeruginosa influences the virulence factors pyochelin, pyocyanin, and biofilm formation. Microbiologyopen 2016, 5, 499–511. [Google Scholar] [CrossRef]
- Li, M.; Long, Y.; Liu, Y.; Liu, Y.; Chen, R.; Shi, J.; Zhang, L.; Jin, Y.; Yang, L.; Bai, F.; et al. HigB of Pseudomonas aeruginosa enhances killing of phagocytes by up-regulating the Type III secretion system in ciprofloxacin induced persister cells. Front. Cell. Infect. Microbiol. 2016, 6, 125. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xia, B.; Li, M.; Shi, J.; Long, Y.; Jin, Y.; Bai, F.; Cheng, Z.; Jin, S.; Wu, W. HigB reciprocally controls biofilm formation and the expression of Type III Secretion System Genes through Influencing the Intracellular c-di-GMP level in Pseudomonas aeruginosa. Toxins 2018, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Sun, C.; Li, Y.; Tang, K.; Ni, S.; Wang, X. Antitoxin HigA inhibits virulence gene mvfR expression in Pseudomonas aeruginosa. Environ. Microbiol. 2019, 21, 2707–2723. [Google Scholar] [CrossRef] [PubMed]
- Liberati, N.T.; Urbach, J.M.; Miyata, S.; Lee, D.G.; Drenkard, E.; Wu, G.; Villanueva, J.; Wei, T.; Ausubel, F.M. An ordered, nonredundant library of Pseudomonas aeruginosa strain PA14 transposon insertion mutants. Proc. Natl. Acad. Sci. USA 2006, 103, 2833–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.-H.; Schweizer, H.P. mini-Tn7 insertion in bacteria with single attTn7 sites: Example Pseudomonas aeruginosa. Nat. Protoc. 2006, 1, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Fürste, J.P.; Pansegrau, W.; Frank, R.; Blöcker, H.; Scholz, P.; Bagdasarian, M.; Lanka, E. Molecular cloning of the plasmid RP4 primase region in a multi-host-range tacP expression vector. Gene 1986, 48, 119–131. [Google Scholar] [CrossRef]
- Chen, R.; Wei, X.; Li, Z.; Weng, Y.; Xia, Y.; Ren, W.; Wang, X.; Jin, Y.; Bai, F.; Cheng, Z.; et al. Identification of a small RNA that directly controls the translation of the quorum sensing signal synthase gene rhlI in Pseudomonas aeruginosa. Environ. Microbiol. 2019, 21, 2933–2947. [Google Scholar] [CrossRef]
- Xia, B.; Li, M.; Tian, Z.; Chen, G.; Liu, C.; Xia, Y.; Jin, Y.; Bai, F.; Cheng, Z.; Jin, S.; et al. Oligoribonuclease contributes to tolerance to aminoglycoside and β-Lactam antibiotics by regulating KatA in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2019, 63, e00212-19. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Weng, Y.; Chen, F.; Liu, Y.; Zhao, Q.; Chen, R.; Pan, X.; Liu, C.; Cheng, Z.; Jin, S.; Jin, Y.; et al. Pseudomonas aeruginosa enolase influences bacterial tolerance to oxidative stresses and virulence. Front. Microbiol. 2016, 7, 1999. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Shi, J.; Liu, C.; Jin, Y.; Li, K.; Chen, R.; Jin, S.; Wu, W. PrtR Homeostasis Contributes to Pseudomonas aeruginosa pathogenesis and resistance against ciprofloxacin. Infect. Immun. 2014, 82, 1638–1647. [Google Scholar] [CrossRef] [Green Version]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S.L. Enhanced annotations and features for comparing thousands of Pseudomonas genomes in the Pseudomonas genome database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [Green Version]
- Skjerning, R.B.; Senissar, M.; Winther, K.S.; Gerdes, K.; Brodersen, D.E. The RES domain toxins of RES-Xre toxin-antitoxin modules induce cell stasis by degrading NAD+. Mol. Microbiol. 2018, 111, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Freire, D.M.; Gutierrez, C.; Garza-Garcia, A.; Grabowska, A.D.; Sala, A.J.; Ariyachaokun, K.; Panikova, T.; Beckham, K.S.; Colom, A.; Pogenberg, V.; et al. An NAD+ phosphorylase toxin triggers Mycobacterium tuberculosis cell death. Mol. Cell 2019, 73, 1282–1291. [Google Scholar] [CrossRef] [Green Version]
- Gazzaniga, F.; Stebbins, R.; Chang, S.Z.; McPeek, M.A.; Brenner, C. Microbial NAD metabolism: Lessons from comparative genomics. Microbiol. Mol. Biol. Rev. 2009, 73, 529–541. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Wei, W.; Hassan, B.H.; Li, J.; Deng, J.-Y.; Feng, Y. A single regulator NrtR controls bacterial NAD+ homeostasis via its acetylation. eLife 2019, 8, e51603. [Google Scholar] [CrossRef]
- Okon, E.; Dethlefsen, S.; Pelnikevich, A.; Van Barneveld, A.; Munder, A.; Tümmler, B. Key role of an ADP-ribose—Dependent transcriptional regulator of NAD metabolism for fitness and virulence of Pseudomonas aeruginosa. Int. J. Med Microbiol. 2017, 307, 83–94. [Google Scholar] [CrossRef]
- Gad, G.F.; El-Domany, R.A.; Ashour, H.M. Antimicrobial Susceptibility Profile of Pseudomonas aeruginosa Isolates in Egypt. J. Urol. 2008, 180, 176–181. [Google Scholar] [CrossRef]
- Elkhatib, W.F.; Khalil, M.A.; Ashour, H.M. Integrons and antiseptic resistance genes mediate resistance of Acinetobacter baumannii and Pseudomonas aeruginosa Isolates from intensive care unit patients with wound infections. Curr. Mol. Med. 2019, 19, 286–293. [Google Scholar] [CrossRef]
- Mann, D.L.; Foale, R.A.; Gillam, L.D.; Schoenfeld, D.; Newell, J.; Weyman, A.E. Early natural history of regional left ventricular dysfunction after experimental myocardial infarction. Am. Heart J. 1988, 115, 538–546. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, C.; Li, Y.; Li, J.; Wan, Q.; Chen, J.; Tay, F.R.; Niu, L. Considerations and caveats in combating ESKAPE pathogens against nosocomial infections. Adv. Sci. 2019, 7, 1901872. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Wei, Y.; Shen, Y.; Li, X.; Zhou, H.; Tai, C.; Deng, Z.; Ou, H.-Y. TADB 2.0: An updated database of bacterial Type II Toxin–Antitoxin loci. Nucleic Acids Res. 2018, 46, D749–D753. [Google Scholar] [CrossRef]
- Ariyachaokun, K.; Grabowska, A.D.; Gutierrez, C.; Neyrolles, O. Multi-Stress induction of the Mycobacterium tuberculosis MbcTA bactericidal toxin-antitoxin system. Toxins 2020, 12, 329. [Google Scholar] [CrossRef]
- Long, Y.; Fu, W.; Li, S.; Ren, H.; Li, M.; Liu, C.; Zhang, B.; Xia, Y.; Fan, Z.; Xu, C.; et al. Identification of novel genes that promote persister formation by repressing transcription and cell division in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2019, 74, 2575–2587. [Google Scholar] [CrossRef]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354, aaf4268. [Google Scholar] [CrossRef]
- Lewis, K. Persister Cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef]
- Shan, Y.; Lazinski, D.; Rowe, S.; Camilli, A.; Lewis, K. Genetic Basis of Persister Tolerance to Aminoglycosides in Escherichia coli. mBio 2015, 6, e00078-15. [Google Scholar] [CrossRef] [Green Version]
- Goormaghtigh, F.; Fraikin, N.; Putrinš, M.; Hallaert, T.; Hauryliuk, V.; Garcia-Pino, A.; Sjödin, A.; Kasvandik, S.; Udekwu, K.; Tenson, T.; et al. Reassessing the role of type II toxin-antitoxin systems in formation of Escherichia coli type II persister cells. mBio 2018, 9, e00640-18. [Google Scholar] [CrossRef] [Green Version]
- Helaine, S.; Cheverton, A.M.; Watson, K.G.; Faure, L.M.; Matthews, S.A.; Holden, D.W. Internalization of Salmonella by macrophages induces formation of nonreplicating persisters. Science 2014, 343, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Claudi, B.; Spröte, P.; Chirkova, A.; Personnic, N.; Zankl, J.; Schürmann, N.; Schmidt, A.; Bumann, D. Phenotypic variation of Salmonella in host tissues delays eradication by antimicrobial chemotherapy. Cell 2014, 158, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Pacios, O.; Blasco, L.; Bleriot, I.; Fernandez-Garcia, L.; Ambroa, A.; López, M.; Bou, G.; Cantón, R.; Garcia-Contreras, R.; Wood, T.K.; et al. (p)ppGpp and its role in bacterial persistence: New challenges. Antimicrob. Agents Chemother. 2020, 64, e01283-20. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J.; Williamson, K.S.; Folsom, J.P.; Boegli, L.; James, G.A. Contribution of stress responses to antibiotic tolerance in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2015, 59, 3838–3847. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Yu, H.; Zhang, D.; Xiong, J.; Qiu, J.; Xin, R.; He, X.; Sheng, H.; Cai, W.; Jiang, L.; et al. Role of ppGpp in Pseudomonas aeruginosa acute pulmonary infection and virulence regulation. Microbiol. Res. 2016, 192, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Vogt, S.L.; Green, C.; Stevens, K.M.; Day, B.; Erickson, D.L.; Woods, D.E.; Storey, D.G. The stringent response is essential for Pseudomonas aeruginosa virulence in the rat lung agar bead and drosophila melanogaster feeding models of infection. Infect. Immun. 2011, 79, 4094–4104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pletzer, D.; Blimkie, T.M.; Wolfmeier, H.; Li, Y.; Baghela, A.; Lee, A.H.Y.; Falsafi, R.; Hancock, R.E.W. The stringent stress response controls proteases and global regulators under optimal growth conditions in Pseudomonas aeruginosa. mSystems 2020, 5, e00495-20. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Source (Reference) |
|---|---|---|
| P. aeruginosa | ||
| PA14 | Wild-type strain of Pseudomonas aeruginosa | [32] |
| ΔPA14_51020 | PA14 deleted of PA14_51020 | This study |
| ΔPA14_51010 | PA14 deleted of PA14_51010 | This study |
| ΔPA14_51020ΔPA14_51010 | PA14 deleted of PA14_51020 and PA14_51010 | This study |
| Plasmid | ||
| pEX18Tc | Gene replacement vector; Tc r | [33] |
| pMMB67EH | Expression vector with tac promoter; Amp r | [34] |
| pUCP20-lacZ | Promoterless lacZ fusion vector; Amp r | [35] |
| pMMB67EH-PA14_51010-Flag | PA14_51010 gene with Flag-tag driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-PA14_40220-Flag | PA14_40220 gene with Flag-tag driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-PA14_21710-Flag | PA14_21710 gene with Flag-tag driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-PA14_28120-Flag | PA14_28120 gene with Flag-tag driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-PA14_28790-Flag | PA14_28790 gene with Flag-tag driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-PA14_60050-Flag | PA14_60050 gene with Flag-tag driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-PA14_71340-Flag | PA14_71340 gene with Flag-tag driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-GST-His | GST with His-tag driven by tac promoter on pMMB67EH; Amp r | [36] |
| pMMB67EH-GST-PA14_51020-His | PA14_51020 gene with His-tag fused to GST driven by tac promoter on pMMB67EH; Amp r | This study |
| pEX18Tc-ΔPA14_51020 | PA14_51020 gene of PA14 deletion on pEX18Tc; Tc r | This study |
| pEX18Tc-ΔPA14_51010 | PA14_51010 gene of PA14 deletion on pEX18Tc; Tc r | This study |
| pEX18Tc-ΔPA14_51020ΔPA14_51010 | PA14_51020 and PA14_51010 gene of PA14 deletion on pEX18Tc; Tc r | This study |
| pUCP20-PPA14_51020-lacZ | PA14_51020 promoter of PA14 on a promoterless lacZ fusion vector; Amp r | This study |
| pMMB67EH-PA14_51020 | PA14_51020 gene driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-operon (PA14_51020-PA14_51010) | The operon (PA14_51020-PA14_51010) driven by tac promoter on pMMB67EH; Amp r | This study |
| pMMB67EH-PA14_51010 (RES-A) | PA14_51010 gene with simultaneous replacement of the R, E, S residues with A residues driven by tac promoter on pMMB67EH; Amp r | This study |
| Primer | Sequence 5′–3′ | Purpose |
| EcoRI-PA14_51020-up-F | CCGGAATTCGCTGGAGTTGCTGACCG | PA14_51020, PA14_51010 deletion |
| KpnI-PA14_51020-up-R | CGGGGTACCGCCCCAATTGCTCGC | PA14_51020, PA14_51010 deletion |
| KpnI-PA14_51020-down-F | CGGGGTACCACGCCTGTCACGGAAAAG | PA14_51020 deletion |
| HindIII-PA14_51020-down-F | CCCAAGCTTTCGCCGAAGCCTCTTGC | PA14_51020 deletion |
| EcoRI-PA14_51010-up-F | CCGGAATTCTCAGCAGCATCCGTCGCGAT | PA14_51010 deletion |
| KpnI-PA14_51010-up-R | CGGGGTACCCTTCCGCCCCTCGCTTCCTG | PA14_51010 deletion |
| KpnI-PA14_51010-down-F | CGGGGTACCGAGCTGTTCCTGGTGG | PA14_51020 and PA14_51010 deletion |
| HindIII-PA14_51010-down-R | CCCAAGCTTGACGCACTTCCTCTTCC | PA14_51020 and PA14_51010 deletion |
| SmaI-PPA14_51020-F | TCCCCCGGGGGCAATGGGCCGATCGAATC | PA14_51020 promoter cloning |
| BamHI-PPA14_51020-R | CGCGGATCCCCCCAATTGCTCGCGCGCGG | PA14_51020 promoter cloning |
| EcoRI-PA14_51010-F | CCGGAATTCTCCGACACCACAGGAAGCGA | PA14_51010 cloning |
| HindIII-PA14_51010-R | CCCAAGCTTTCATTTATCATCATCATCTTTGTAATCCGCCGGATGCGGCA | PA14_51010 cloning |
| BamHI-PA14_51020-F | CGCGGATCCACGCAGCTCGAACTGGCCGG | PA14_51020 cloning |
| HindIII-PA14_51020-R | CCCAAGCTTTCAGTGGTGGTGGTGGTGGTGGACCTTGCCGCGGATCGCAT | PA14_51020 cloning |
| EcoRI-GST-F | CCGGAATTCTTTAAGAAGGAGATATAATGTCCCCTATACTAGGTTA | PA14_51020 cloning |
| BamHI-GST-R | CGCGGATCCACCAGAACCACTAGTTGAAC | PA14_51020 cloning |
| BamHI-PA14_40220-F | CGCGGATCCGCTCGTTTCACCGGTAGCGG | PA14_40220 cloning |
| HindIII-PA14_40220-R | CCCAAGCTTCTATTTATCATCATCATCTTTGTAATCGGCGTCGCGCCGA | PA14_40220 cloning |
| BamHI-PA14_21710-F | CGCGGATCCACGCTCTGATGGGAGCGGAG | PA14_21710 cloning |
| HindIII-PA14_21710-R | CCCAAGCTTTCATTTATCATCATCATCTTTGTAATCGCCGGTGAAGCTGGCT | PA14_21710 cloning |
| EcoRI-PA14_28120-F | CCGGAATTCCCGCCAGCCTGTACGCACAA | PA14_28120 cloning |
| BamHI-PA14_28120-R | CGCGGATCCTCATTTATCATCATCATCTTTGTAATCGCCTCGCGCCAGT | PA14_28120 cloning |
| EcoRI-PA14_28790-F | CCGGAATTCCAGCATATGCGGGAGCTGTT | PA14_28790 cloning |
| HindIII-PA14_28790-R | CCCAAGCTTTCATTTATCATCATCATCTTTGTAATCGTGAGTACCAGCCC | PA14_28790 cloning |
| EcoRI-PA14_60050-F | CCGGAATTCGAGCTCGGCAACCAGGCGAG | PA14_60050 cloning |
| HindIII-PA14_60050-R | CCCAAGCTTTCATTTATCATCATCATCTTTGTAATCTCGTTGGGGCAGGT | PA14_60050 cloning |
| EcoRI-PA14_71340-F | CCGGAATTCCCCCGCTCCACCCTTTCCCA | PA14_71340 cloning |
| HindIII-PA14_71340-R | CCCAAGCTTTCATTTATCATCATCATCTTTGTAATCTTGAGGTTGCT | PA14_71340 cloning |
| EMSA-upstream-F | GTTTTTCTCTCTATCACGCC | EMSA |
| EMSA-upstream-R | CCCCAATTGCTCGCGCGCGG | EMSA |
| EMSA-inside-F | CGGGCTGGAAGGTGGAGCGG | EMSA |
| EMSA-inside-R | CGCGGGCGTGAACAGGGCGA | EMSA |
| PA14_51010-F | GAGCCAAGCCTGTTCTAC | qRT-PCR |
| PA14_51010-R | CAGGACACAACGGTAATACG | qRT-PCR |
| PA14_51020-F | CACTCCCAACCATCAC | qRT-PCR |
| PA14_51020-R | AGGTATTCCAGCACAT | qRT-PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Li, S.; Li, H.; Jin, Y.; Bai, F.; Cheng, Z.; Wu, W. Identification of a Toxin–Antitoxin System That Contributes to Persister Formation by Reducing NAD in Pseudomonas aeruginosa. Microorganisms 2021, 9, 753. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040753
Zhou J, Li S, Li H, Jin Y, Bai F, Cheng Z, Wu W. Identification of a Toxin–Antitoxin System That Contributes to Persister Formation by Reducing NAD in Pseudomonas aeruginosa. Microorganisms. 2021; 9(4):753. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040753
Chicago/Turabian StyleZhou, Jingyi, Shouyi Li, Haozhou Li, Yongxin Jin, Fang Bai, Zhihui Cheng, and Weihui Wu. 2021. "Identification of a Toxin–Antitoxin System That Contributes to Persister Formation by Reducing NAD in Pseudomonas aeruginosa" Microorganisms 9, no. 4: 753. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040753