1. Introduction
Current estimates project 40% of all plant species will become extinct within 30 years due to anthropogenic factors and climate change [
1] and these dire projections have instilled a sense of urgency amongst conservationists to safeguard the species at highest risk. Despite being incredibly diverse with upwards of 25,000 species worldwide [
2], orchids are considered highly vulnerable to extinction. According to the International Union for Conservation of Nature (IUCN), half of all orchids that have already succumbed to extinction are herbaceous terrestrials despite comprising only about one-third of all species worldwide [
3]. Depending on the species and where it grows, some terrestrial orchids may live for decades (e.g.,
Cypripedium calceolus), whereas others are ephemeral, quickly colonizing an area, setting seed, and perishing shortly thereafter within the span of one year (e.g.,
Habenaria repens [
4]). For conservationists faced with the burden of securing orchids under immediate threat, targeting the short-lived terrestrials is a logical priority. To date, those that have received the most study with respect to their biotic and abiotic requirements are rooted in temperate regions, and surprisingly little is known about their tropical counterparts. Considering that most global biodiversity "hotspots" occur in the tropics where 90% of endemic orchids are concentrated, studying these species is of utmost importance. Unfortunately, many of these hotspots are in developing countries that have few monetary resources and, consequently, they remain highly vulnerable to deforestation and other threats (e.g., agricultural practices, mining, political unrest, and poaching). Some of these areas are also remote, making their access by researchers very costly and at times hazardous.
Their vulnerability and success appear to be linked to the close associations they form with other organisms, namely, winged pollinators for sexual reproduction, and mycorrhizal fungi for seed germination and growth through to maturity. These biotic factors therefore transcend all parts of the plant throughout every phase of its life. The orchid, and the organisms they rely on, are also intimately tied to the abiotic factors present in their surroundings such as moisture [
5], soil pH [
6], sunlight [
7], and nutrients [
8], all of which may vary considerably along the terrain in each habitat [
9,
10,
11]. As a result, orchids are widespread but limited in where they grow, and they are acutely sensitive to subtle environmental changes once they become established from seed. Conserving these remarkable plants, therefore, poses many challenges that are exacerbated in this age of climate change, and fully understanding these complex interactions is a primary goal.
Despite occupying only 0.04% of the global land surface, the island nation of Madagascar is home to 4% of the world’s biological diversity, and an extraordinary level of endemism among its native flora and fauna [
12]. For example, of its estimated 10,000 vascular plant species, 90% are found nowhere else on earth [
13] and this is also true of 1000 orchid species within 57 genera [
14]. Given that 90% of Madagascar’s original vegetation has been cleared or permanently altered, many orchid populations throughout the country have been reduced in size to fragmented patches where seedling recruitment is generally low [
15], possibly due to inbreeding depression [
16]. Nevertheless, these forest remnants still support a surprisingly diverse orchid flora. A recent study revealed that Malagasy inselbergs (isolated mountains of granitic or gneissic origin) in the Central Highlands accommodated 87 orchid species despite frequent disturbances present in the region (e.g., annual burning, mining, cattle raising) [
17]. They also revealed that
Cynorkis is the most abundant genus in the region (18 species) whose members are typically short-lived, appealing terrestrial herbs that form a dense vegetative layer over shallow, nutrient-poor substrates. Collectively, these species persist in “micro-hotspots” or even smaller patches termed “nano-hotspots” where exceptional endemism may be concentrated in areas < 3 km
2 [
18]. The scale of diversity within these small hotspots offers a unique opportunity to study the fungal associates of these orchids, and by doing so these plants could potentially be propagated artificially from seed and reintroduced, augmenting their long-term conservation.
The genus
Cynorkis encompasses 170 species, with 120 found in Madagascar alone [
19], and the vast majority of which remain unstudied with respect to their mycorrhizal associations. Yokoya et al. [
20], however, revealed that one species (
Cynorkis purpurea), native to stream beds in the Central Highlands, harbored a surprisingly rich assemblage of
Rhizoctonia-like fungi that typically form mycorrhizal associations (
Ceratobasidium,
Serendipita,
Tulasnella). When seeds of
C. purpurea were later inoculated with these fungi in vitro, the mycorrhizal nature of the fungi was confirmed [
21] suggesting that this orchid may be a “fungal generalist” [
1]. The success of these two studies served as the impetus for further work involving other
Cynorkis species in the region.
The aim of this paper is to characterize the fungal diversity associated with at least 50% of Cynorkis species found in the micro-hotspot of the Central Highlands in Madagascar, including their mycorrhizal associates as well as non-mycorrhizal (saprotrophic) inhabitants. In addition, we attempt to link fungal diversity to various abiotic soil factors (e.g., pH, N, P, and K nutrient levels) at various geographic locations to garner a broader picture of how these orchids function and persist in nature. To our knowledge this is the first study of its kind for an orchid genus from Madagascar. Our goal is to provide more information for a poorly understood but well-known orchid genus in one of the most remote, highly vulnerable, and biologically significant regions on earth.
3. Results
Collecting orchid and soil samples from the Central Highlands of Madagascar for the current study was challenging due to problems with access, number of days required to screen and culture in vitro over a three-week period. The number of roots that yielded pelotons was relatively low (
Supplementary Materials File S1). A smart kit is the answer to collect and culture pelotons within couple of days of collecting which will help recover all peloton-forming culturable fungi collected from the wild. Although, in many species, more than two plants were sampled recovery of
Rhizoctonia OTUs was significantly low. Except in
Cynorkis fastigiata, which yielded tul5, all
Rhizoctonia OTUs were identified from roots of mature plants.
3.1. Distribution of Mycorrhizal and Non-Mycorrhizal Fungi
Culturable mycorrhizal fungi were obtained from nine of the 11 Cynorkis species:
Cynorkis flexuosa,
Cynorkis fastigiata,
Cynorkis gibbosa,
Cynorkis graminea,
C. purpurea,
C. ×
ranaivosonii,
Cynorkis ridleyi,
Cynorkis speciosa. and
Cynorkis sp.) while two species (
Cynorkis papillosa and
Cynorkis uniflora) failed to yield fungal endophytes. Cantharellales (
Ceratobasidium and
Tulasnella) fungi were successfully isolated from eight out of 11 species while members of Serendipita (
Sebacinales) were recovered in four species (
Figure 4). Except
C. papillosa and
C. uniflora, all other species had mycorrhizal OTUs in their root cortical cells.
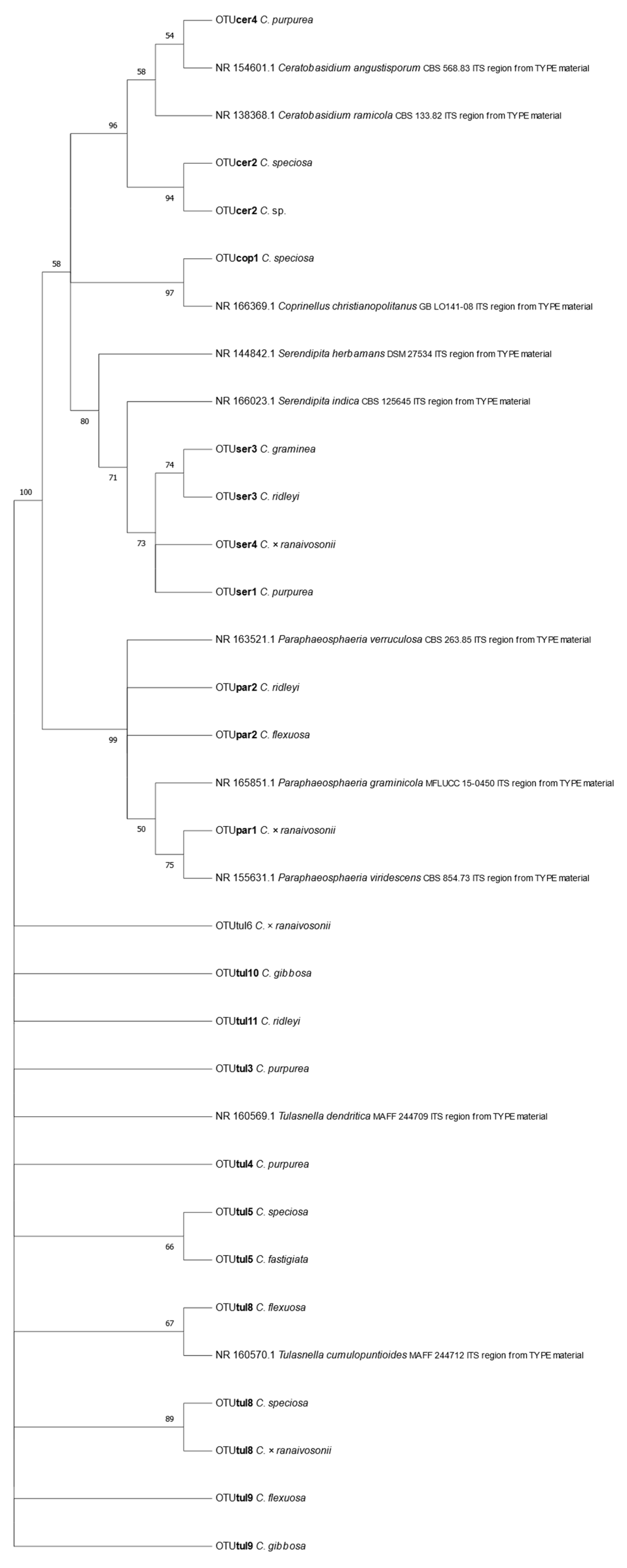
Of the fungi obtained, mycorrhizal fungal OTUs from the genera Ceratobasidium, Serendipita, and Tulasnella were identified in addition to orchid mycorrhizae such as Coprinellus and Paraphaeosphaeria. In total two Ceratobasidium, three Serendipita and eight Tulasnella OTUs were associated with nine of the Cynorkis species. OTU tul8 was found in three Cynorkis species, OTUs tul5 and tul9 in two species while the remainder were recorded only once in the genus. Culturable mycorrhizal OTUs were distributed in varied numbers with four in C. purpurea while C. × ranaivosonii and C. speciosa had three OTUs. Only a single OTU each were recovered from C. graminea and Cynorkis sp.
In total there were13
Rhizoctonia OTUs identified from nine of the 11
Cynorkis species of which eight were
Tulasnella, three were
Serendipita and two were
Ceratobasidium (
Figure 4).
Cynorkis purpurea and
C. ×
ranaivosonii hosted four OTUs followed by,
C. flexuosa,
C. ridleyi and
C. speciosa (three each). OTUs tul8, tul9, cer2 seb3 were the only OTUs found in two
Cynorkis spp.
C. fastigiata and
C. graminea had a single OTU each. Non-Rhizoctonial
Paraphaeosphaeria was identified from three taxa (
C. flexuosa,
C. ×
ranaivosonii and
C. ridleyi).
3.2. Phylogenetic Characterization of Cynorkis spp.
The primary aim of genotyping the host plants was to confirm the taxonomic status of protocorms, seedlings and plants without flowers from which root materials were collected. All sequenced juvenile samples matched those of the expected Cynorkis species following sequencing of the chloroplast DNA region trnL-F. Phylogenetic analysis showed that C. graminea and C. ridleyi were of the same clade and they were also closely related to C. papillosa and Cynorkis sp., all of which consist of relatively small plants with small flowers.
3.3. Fungal Diversity Shannon’s Index and (H) and Simpson’s Index (D) of Cynorkis spp.
Figure 5A–C shows H and D for mycorrhizal (A), non-mycorrhizal (B) and combined (C) groups). For mycorrhizal group (
Figure 5A), H and D were higher for four species namely
C. speciosa, C. purpurea,
C. ×
ranaivosonii and C. ridleyi in comparison to others whereas the latter two were amongst the top in H and D for the non-mycorrhizal group (
Figure 5B). When mycorrhizal and non-mycorrhizal groups were combined, H and D showed slightly different trend as
C. ridleyi >
C. graminea>
C. ×
ranaivosonii. Our analyses revealed the diversity values for different
Cynrokis spp. For example,
C. ×
ranaivosonii followed by
C. ridleyi from wet habitats (arb. value 3), had the highest diversity index values (
Supplementary Materials File S2). However, the combined diversity index of 10 species (excluding
C. purpurea) shows
C. ridleyi harbored the most fungal diversity (only two mycorrhizal fungi identified-
Tulasnella and
Serendipita) followed by
C. graminea which had
Serendipita as the lone mycorrhizal fungus. Combined analysis for diversity using Shannon and Simpson’s shows
C. ridleyi harboring the most diverse assemblage of fungi identified from its roots.
C. purpurea followed by
C. ×
ranaivosonii from wet habitats (arb. value 3) had the highest diversity for mycorrhizal fungi (as confirmed by Shannon diversity analysis in
Figure 5) while
C. ridleyi came second in diversity index according to Simpson’s index.
3.4. Relationships between Abiotic Factors and the Fungal Diversity
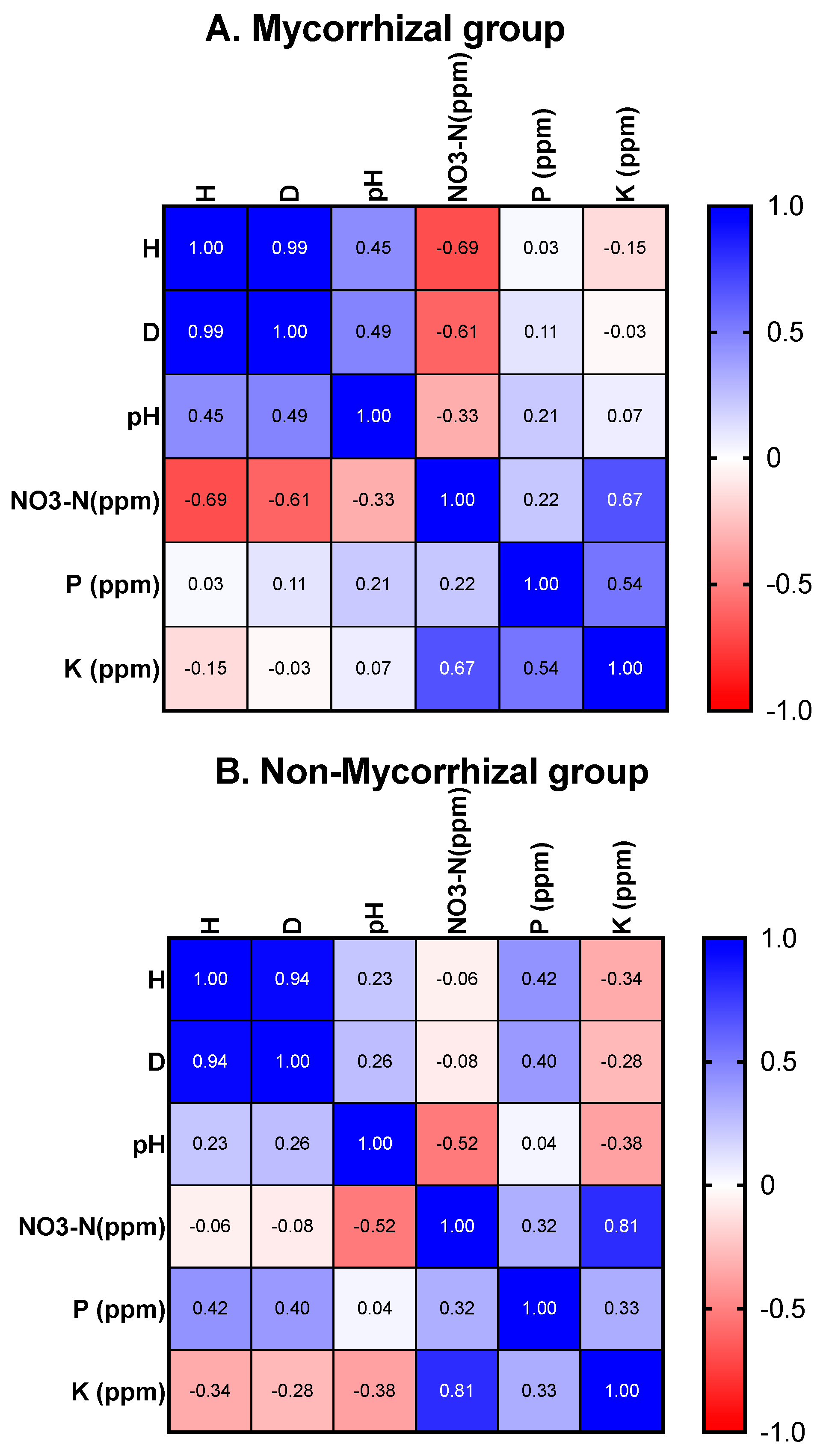
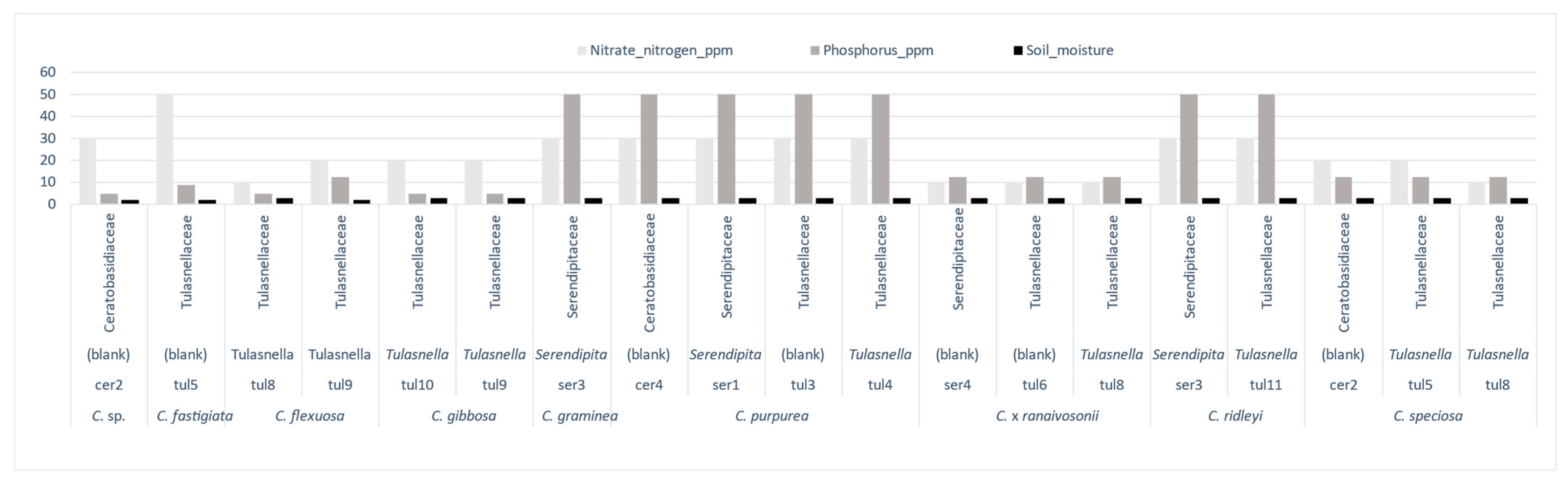
Except for C. purpurea, the link between these Cynorkis species, fungal diversity (H) and (D) (mycorrhizal and non-mycorrhizal), and soil characteristics is a mixed one. For example, the Cynorkis species documented in this study were found growing in seasonally wet, free-draining soil, somewhat peaty and high in organic matter at varying stages of decomposition. Very little loam, sand, silt, or other sediments were observed. Soil sample analysis revealed that all Cynorkis species inhabited soils that were nutritionally poor and slightly acidic. Average values (for 102 mycorrhizal and non-mycorrhizal OTUs) were in the lower scale for respective minerals. NO3-N (24 ppm, 10 ppm lowest value), P (18.4 ppm, 5 ppm lowest value), K (110 ppm, 50 ppm lowest value), and average pH was 5.2. Our observations showed that out of the 11 species studied, three (Cynorkis sp., C. fastigiata, C. flexuosa) were growing in a moist soil environment (arbitrary value of 2) while the others were associated with a wetter (value 3) soil.
Based on the measured soil properties (pH, NO
3-N, P, and K) and calculated diversity indices (H and D), Pearson correlation heatmaps are depicted in
Figure 6 for mycorrhizal (
Figure 6A), non-mycorrhizal (
Figure 6B) and combined (
Figure 6C) for all taxa of except
C. purpurea. Note that
p values are provided in the
Supplementary Materials File S2.
Tulasnella OTUs were found in all soil types while
Serendipita OTUs were found in species inhabiting in granite/rock areas close to river/seepage. Diversity of mycorrhizal OTUs was comparably higher in wet habitat as shown in
Figure 7 (arbitrary score of 3).
C. flexuosa (tul8 and tul9)
C. ×
ranaivosonii (ser4, tul6, and tul8),
C. speciosa (cer2, tul5, and tul8) and
C. gibbosa (tul9, tul10) were from soil with low NO
3-N and P content. These orchids recruited either tul8 or tul9, two closely related OTUs (
Figure 4). Of these four species tul8 was present in all but
C. gibbosa which recruited tul9. This means low P and N favored orchids to recruit a selection of
Tulasnella OTUs and small number of
Serendipita and
Ceratobasidium OTUs.
On the contrary,
C. graminea,
C. purpurea and
C. ridleyi were the only three species found in N and P rich soils, P (50 ppm) and N (30 ppm). Under same soil conditions
C. graminea and
C. ridleyi recruited a single
Serandipita OTU and an additional tul11 by
C. ridleyi. Both were from similar moisture conditions. In
C. ridleyi a diminutive species putative mycorrhizal OTUs ser3 and tul11 were identified from plants collected from soil with 50 ppm of
p while plants from soil with 9 ppm of P had Hypocreales and Helotiales fungi in addition to other non-mycorrhizal fungi (
Figure 7).
When all
Cynorkis spp. were analyzed (except
C. purpurea) NO
3-N, pH, P, and K are found to be correlated with diversity of fungi (H and D). Specifically, mycorrhizal diversity is inversely correlated to NO
3-N in
Cynorkis spp. (
Figure 6A). However, pH is positively correlated with both diversity indices (the difference between pH values were miniscule though). This revealed a positive correlation between higher pH values and higher orchid diversity and abundance. When different taxa were analyzed separately most of the orchids studied favored lower nitrogen and P levels (
Figure 7).
P is positively correlated with diversity (H and D) of non-mycorrhizal for all
Cynorkis taxa (except
C. purpurea). This indicates that more non-mycorrhizal fungi were recruited when P was comparatively high. K content was negatively correlated with diversity (H) of non-mycorrhizal fungi (
Figure 6B). Likewise, P is positively and significantly (
p = 0.009) correlated with D when mycorrhizal and non-mycorrhizal fungi are combined (
Figure 6C).
Paraphaeosphaeria OTUs, a group of putative mycorrhizal fungi, were detected in
C. flexuosa,
C. ×
ranaivosonii, and
C. ridleyi while
Coprinellus was found in
C. speciosa.
Cynorkis ×
ranaivosonii followed by
C. ridleyi, then
C. graminea had the highest non-mycorrhizal fungal diversity (as shown by Shannon and Simpson’s diversity analyses (
Figure 6B and
Figure 8). Data on non-mycorrhizal fungi of
C. purpurea were missing because this species was not collected and included for further analysis. Hence, results on
C. purpurea are incomplete.
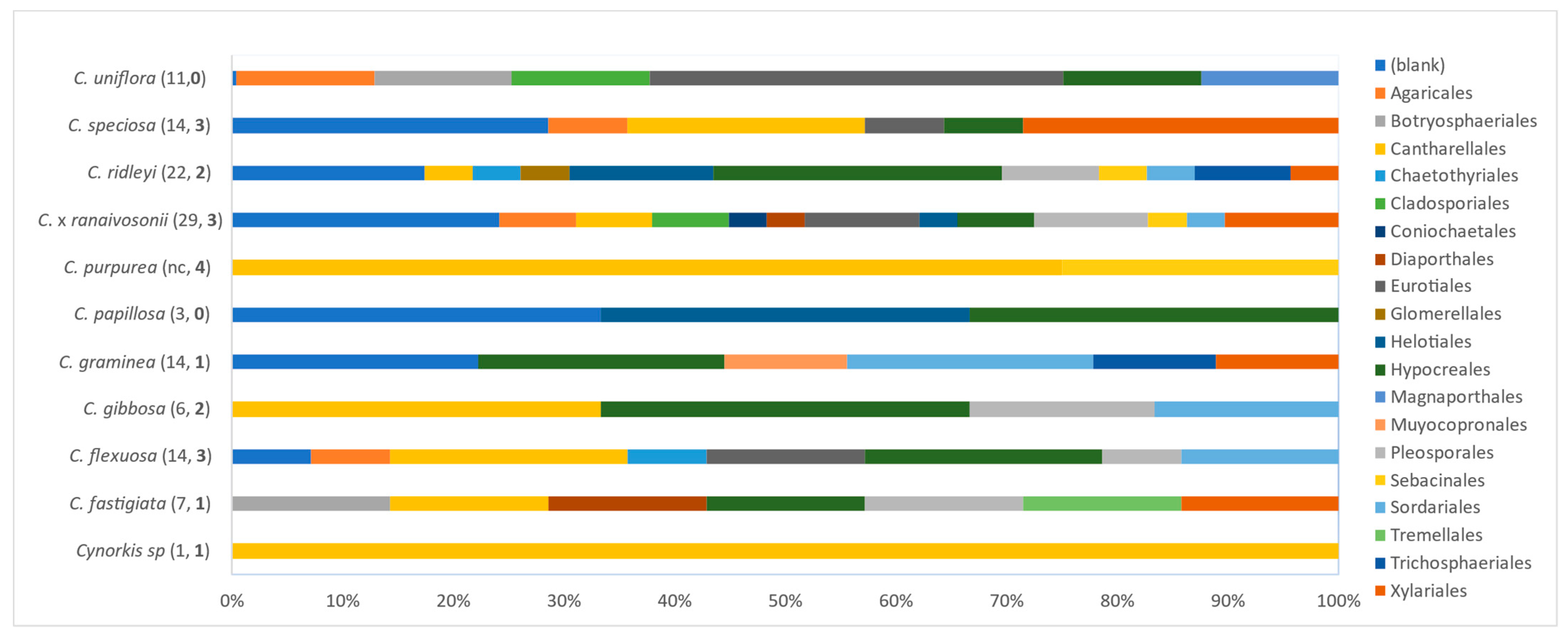
The fungal diversity in 11 species of Cynorkis was statistically significant based on P content in the soil. However, data were not significant when calculated separately for mycorrhizla and non-mycorrhizal fungi. Hypocreales, an order characterized by having dark-septate hyphae, were recruited invariably by these orchids when Rhizoctonia lineages were absent.
Two smallest herbaceous orchids,
C. graminea and
C. ridleyi, growing in arb wet = 3, harbored the most non-mycorrhizal fungi, second only to
C. ×
ranaivosonii (
Figure 8). Members of the Hypocreales colonized nine out of 10 species assessed (
C. purpurea excluded). Fungi assignable to the Eurotiales and Xylariales were secondary inhabitants, found in five species each. Members of the Eurotiales, Pleosporales, Sordariales, and Xylariales were present in nearly 50% of
Cynorkis species (
Figure 8) suggesting that a variety of dark-septate endophytes colonize the roots of this genus. Interestingly, fungi assignable to Pleosporales and Xylariales were found only in roots that were also colonized with mycorrhizal fungi. Further, worth noting were the presence of Psathyrellaceae OTUs (Agaricales comprises ectomycorrhizal fungi such as
Coprinellus) detected in four
Cynorkis species. Only a few OTUs were recovered from
Cynorkis sp. and
C. papillosa due to deterioration of root samples over the three week period.
4. Discussion
The Central Highlands of Madagascar is characterized by tough terrain that severely restricts access to orchid populations found clustered along the region’s inselbergs in pockets where moisture collects and only accessible by foot. Restrictions by the Madagascan authorities on sample collecting also limited the number of root/protocorms that we were able to collect. Despite these hardships, we were able to obtain enough samples to provide a snapshot of Cynorkis diversity in the Central Highlands linked to soil, moisture, nutrients, and fungal diversity for a respectable number of species.
About 10% of all angiosperm species are orchids which are associated with roughly 25,000 Basidiomycota and some Ascomycota as fungal partners [
36,
37]. These associations are guided by soil characteristics, seasonality, and life forms of the orchids [
9,
10,
11].
To our knowledge, this is the first study that correlated soil nutrients, moisture, and fungal diversity in a well-known group of terrestrial orchids (Cynorkis spp.), the majority of which are endemic to Madagascar. These results could have a long-term impact on the conservation of species native to central Madagascar especially for reintroduction efforts being planned that will require navigating hardships imposed by climate change.
Most of the 11
Cynorkis species studied herein occupied two primary habitats—gallery forests and grasslands. Historically these species would have been abundantly distributed in wet soil (arb value 3) but may have adapted to less wet (arb value 2) soil due to deforestation and ongoing climate change. In a climate change scenario of erratic rainfall and drought, it is plausible that these
Cynorkis species will face increasing challenges in survival and natural regeneration. Accordingly, mycorrhizal diversity needed by these orchids for seed germination and survival must be maintained to reduce population decline in these unique habitats that support the majority of
Cynorkis spp. Even though some
Cynorkis spp. appear to be resilient in moist (arb 2) areas, further environmental changes that permanently alter these habitats by stripping critically needed moisture may impede the long-term survival of these species. The results of a previous in vitro symbiotic seed germination study by Rafter et al. [
21] demonstrated that
C. purpurea is a generalist. Most OTUs were identified from
C. purpurea in this study which may also suggest the generalist nature. Nurfadilah et al. [
38] demonstrated that different soil N forms (ammonium and nitrate) can be exploited by generalists possibly because the co-occurrence of mycorrhizal fungi enables the plants to utilize a wider range of nutrient sources.
Ephemeral flush vegetation as reported by Rabarimanarivo et al. [
17] has a meadow-like physiognomy and display seasonality between rainy and dry season known to support genera such as
Cynorkis. Most members of this genus occupy gently inclined rocky slopes where water constantly seeps during the rainy season and in some cases even during the dry season. Although
Cynorkis is distributed in many other parts of Madagascar, the majority are found in wetter habitats which make them vulnerable to population decline in the absence of optimal moisture levels. Annual wet season rains may represent a key factor in generating spontaneous seedlings, and that if this cycle is broken in successive years, some
Cynorkis taxa will likely suffer from population decline. As this biodiversity hotspot has experienced extensive deforestation and fragmentation, climate change is expected to exacerbate species loss in vulnerable habitats in the decades to come [
39].
Out of 11 species, C. speciosa, C. purpurea, C. × ranaivosonii, and C. ridleyi had highest biodiversity indices for mycorrhizal fungi. When mycorrhizal and non-mycorrhizal groups were combined, H and D showed slightly different trend as C. ridleyi > C. graminea with the highest indices. Combined analysis for diversity using Shannon and Simpson’s shows C. ridleyi harboring the most diverse assemblage of fungi identified from its roots. The small-flowered C. graminea and C. ridleyi, inhabiting close to fast flowing river from lithophytic habitat hosted the greatest number of fungi. This may be because these species undergo more environmental fluxes than other orchids which would drive them to recruit a diverse fungal assemblage for their resilience in the habitat.
Diversity (H and D) of
Cynorkis spp. were correlated with NO
3-N, pH, P, and K. The general picture is showing less nitrogen favored greater mycorrhizal diversity. However, when each species from unique habitats and soil types were assessed separately, they showed low P in addition to NO
3-N favored distribution of
Cynorkis spp. and the assemblage of mycorrhizal fungi. Therefore, for the purpose of understanding specific requirements for reintroduction, targeted environmental analysis is required. Previous studies indicate that low P content may be linked to higher mycorrhizal diversity in
Cephalanthera rubra [
40] and several orchids from Madagascar [
11]. Likewise, Van der Ent et al. [
41] found that the rhizosphere soil concentrations of P and K was low in soil in the study site where
Paphiopedilum rothschildianum was growing. Figura et al. [
42], based on in vitro seed germination studies, found that nitrates negatively affect symbiotic seed germination. The current study showed less NO
3-N and higher P favored greater mycorrhizal diversity.
Cynorkis spp. had high diversity (H and D indices) for non-mycorrhizal fungi when P content was high and K content was low. As Bell et al. [
40] reported with
Cephalanthera rubra, members of
Cynorkis also recruited more non-mycorrhizal fungi when soil P values were comparatively high. The Central Highlands landscape is prone to eutrophication due to burning and grazing. Nitrate leeching into soil and finding their way into streams where
Cynorkis are rooted is a real possibility which may adversely affect seedling recruitment. A more detailed analysis, with soil samples from wider population ranges, will be required to demonstrate the likelihood that this may occur.
In
C. ridleyi, plants collected from soil with 50 ppm of P had two putative mycorrhizal OTUs, ser3 and tul11, while plants from soil with 9 ppm of P had non-mycorrhizal fungi like Hyopcreales and Helotiales. The distribution of soil fungi is related to spatial variation and soil chemistry [
43,
44,
45], the suite of fungi the orchid hosts is correlated to the soil environment, indicating how soil chemistry determines orchid fungal associations across landscapes. However, Waud et al. [
46] noted that while mycorrhizal specificity is not critical for the distribution of an endangered mycorrhizal generalist orchid (
Liparis loeselii), germination was affected by soil moisture content. Thus, the distribution of rare orchid species may depend on a complex interaction between soil parameters and the soil mycobiota. Further studies to understand orchid recruitment are needed to reach conclusions on the causes of rarity of
Cynorkis orchids.
We studied 11
Cynorkis species of which nine yielded 13 mycorrhizal fungi. While eight had
Tulasnella OTUs,
Serendipita was only observed in three species suggesting that
Serendipita is more limited in distribution in granite/rocky habitats. These fungi when available, it may have considerable potential at generating seedlings. Rafter et al. [
21] revealed that in vitro seedling development in
C. purpurea was supported by a wide suite of fungi but was most pronounced with
Serendipita OTUs. The phylogenetic tree of mycorrhizal fungi shows the
Serendipita fungal OTUs are associated with phylogenetically close
C. graminea, and
C. ridleyi. This observation is important, and requires further study linking phylogeny of orchid species, mycorrhizal fungi, and their habitats, in this case granite outcrops which has a thin layer of topsoil.
Ascomycetous
Paraphaeosphaeria was also present in
C. flexuosa,
C. ×
ranaivosonii, and
C. ridleyi while
Coprinellus was found in
C. speciosa. These species may be of generalist nature or we may incidentally have collected the healthiest samples. Molecular data in a forage grass revealed that the root mycobiota was dominated by
Paraphaeosphaeria spp. which was clearly tied to water amendments and the addition of similar volumes of water [
47]. They also suggested that
Paraphaeosphaeria was involved with a common but complicated strategy of water translocation in a variety of soil types, like that shown in the arbuscular mycorrhizal fungi. Whether or not this strategy is also employed in
Cynorkis is not known but warrants further study.
Yagame et al. [
48] revealed for the first time that members of the basidiomycete genus
Coprinellus (Psathyrellaceae) have the potential to serve as orchid symbionts capable of facilitating seed germination in a photosynthetic orchid. In a subsequent study by Shao et al. [
49], OTUs assignable to
Coprinellus were present in protocorms of several epiphytic
Dendrobium species, although in their germination trials, these
Coprinellus isolates failed to stimulate symbiotic germination in vitro. Yokoya et al. [
11] reported
Coprinellus sp. was identified in
Liparis ochracea collected from Madagascar.
Four orchid taxa recruited Helotiales in this study, and these fungi were included as putative mycorrhizal associates in studies pertaining to fungal diversity in
Epipactis spp. and
Dactylorhiza spp. [
37,
50]. They found that, in
Epipactis spp., Helotiales was the most abundant alleged mycorrhizal fungal group. For
Paphiopedilum spicerianum inhabiting natural habitats, the most significant factors for fungal communities were hydrolysableN, totalP, and water content [
51], where it was found that fungal community change could be related to nutrient requirements and plant phenology.
Serendipita isolates from
C. graminea and
C. ridleyi were of the same OTU (OTUser3) that was previously found to support seedling growth of
C. purpurea [
21]. These two species are phylogenetically closely related and, like
C. purpurea, are relatively diminutive among the
Cynorkis species that were seen growing at the margins of small rivers. These species may have retained juvenile characteristics that predispose them to continue their association with fungal symbionts from seedling stage. OTU ser3 may be one of the favored OMFs of
Cynorkis species, especially for juvenile stages supporting results of the in vitro symbiosis seen in
C. purpurea [
21]. Studies by Djordjević et al. [
9] showed that among other variables soil moisture, altitude, and soil pH were the main factors affecting the distribution and abundance of orchid species in grasslands and herbaceous wetlands in western Serbia. As herbaceous terrestrial orchids are more vulnerable to extinction compared to other orchids, species like
C. graminea and
C. ridleyi found in riverbanks may be vulnerable to population decline due to climate change related moisture imbalances.
5. Conclusions
All species studied had either mycorrhizal fungi or Hypocreales fungi. Only one juvenile sample yielded a Rhizoctonia OTU (from a protocorm of C. fastigiata) which is a striking finding. Tulasnella fungi were identified from different soil types while Serendipita were only from Cynorkis spp. from rocky/granite close to rivers or have water seepages. Phylogenetically closely related, two small-flowered species, C. graminea and C. ridleyi, hosted the same Serendipita OTU. They were from the banks of a river in a lithophytic habitat which calls for further studies to understand these associations and its influence on species resilience.
Diversity of mycorrhizal fungi were negatively correlated with NO3-N while higher P content favored greater non-mycorrhizal diversity (H and D indices). However, when different species from unique habitats were assessed separately there were clear differences. The majority of the Cynorkis spp. studied were found in soils with low NO3-N and P contents. Specifically, most Tulasnella OTUs, the dominant mycorrhizal lineage, were found in soil with low P content. Although this study was hindered by the small sample sizes imposed by permit restrictions, our findings have indicated avenues of future investigations. Detailed studies on other Cynorkis species from different regions of Madagascar with varied soil types and moisture levels will shed more light on the association between different fungal groups and members of this important genus.
The majority of
Cynorkis species the most dominant herbaceous orchid genus in the Central Highland inselbergs of Madagascar [
17]—are found in soil which is either moist or wet for at least part of the year [
14]. Climate change, illegal collecting, and other anthropogenic pressures will continue to undermine orchid population resilience in the coming decades in the island nation. Detailed studies are required to understand the similarities and differences between these orchids and how they adapt to different ecological conditions. This will help develop pragmatic conservation policies underpinned by symbiotic technology and soil data for effective reintroduction/reinforcement to augment populations vulnerable to decline.
Our results advocate the need to introduce Cynorkis seedlings to primarily wet habitats, beginning with an explorative study to assess seedling establishment and survival. Considering that members of this genus appear to be short-lived (ephemeral) compared to other terrestrial orchids, establishing self-sustaining populations capable of anthesis and seedling recruitment from these plants is another conservation priority.



 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}