
Growth of Staphylococcus epidermidis on the Surface of Teatcups from Milking Parlours

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Staphylococcal Isolates
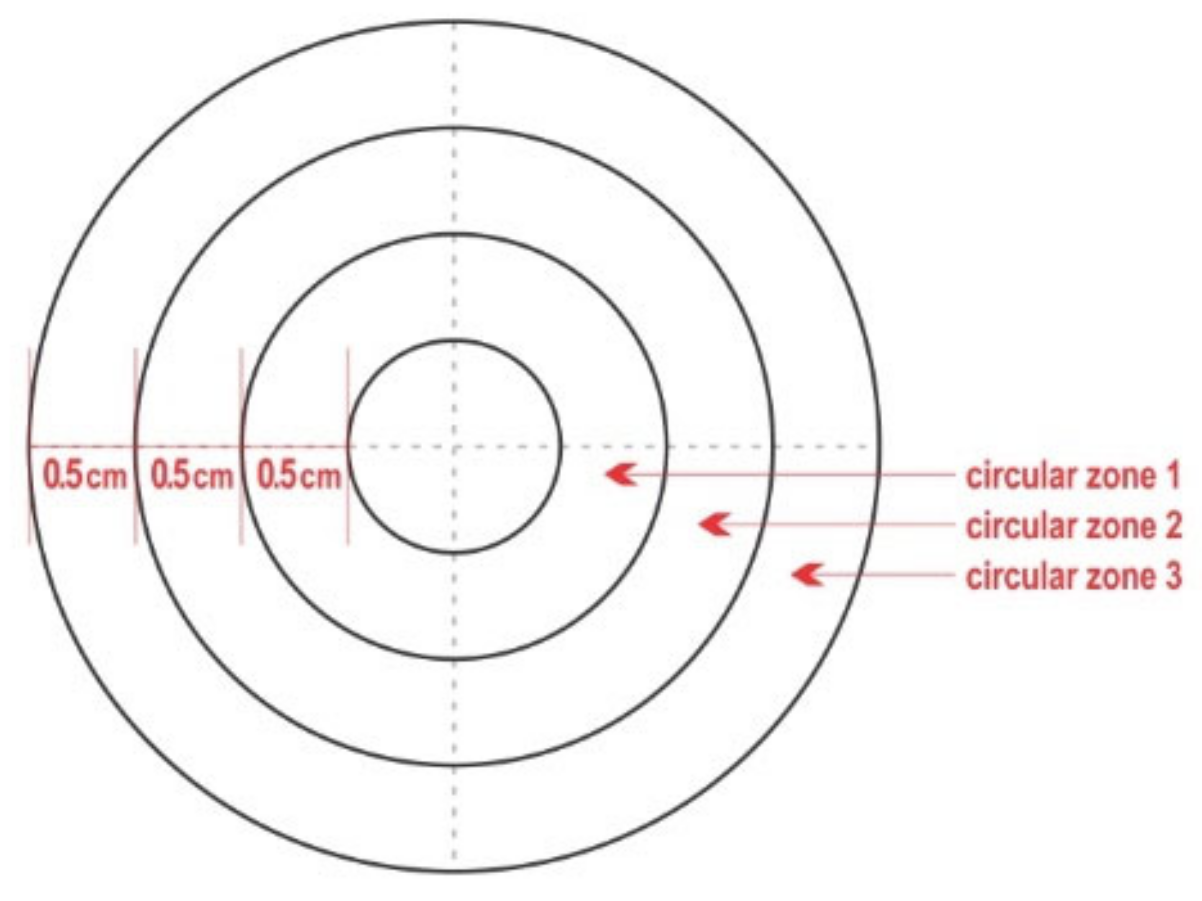
2.2. Test Material (Teatcups)
2.3. Experimental Work
2.4. Data Management and Statistical Analysis
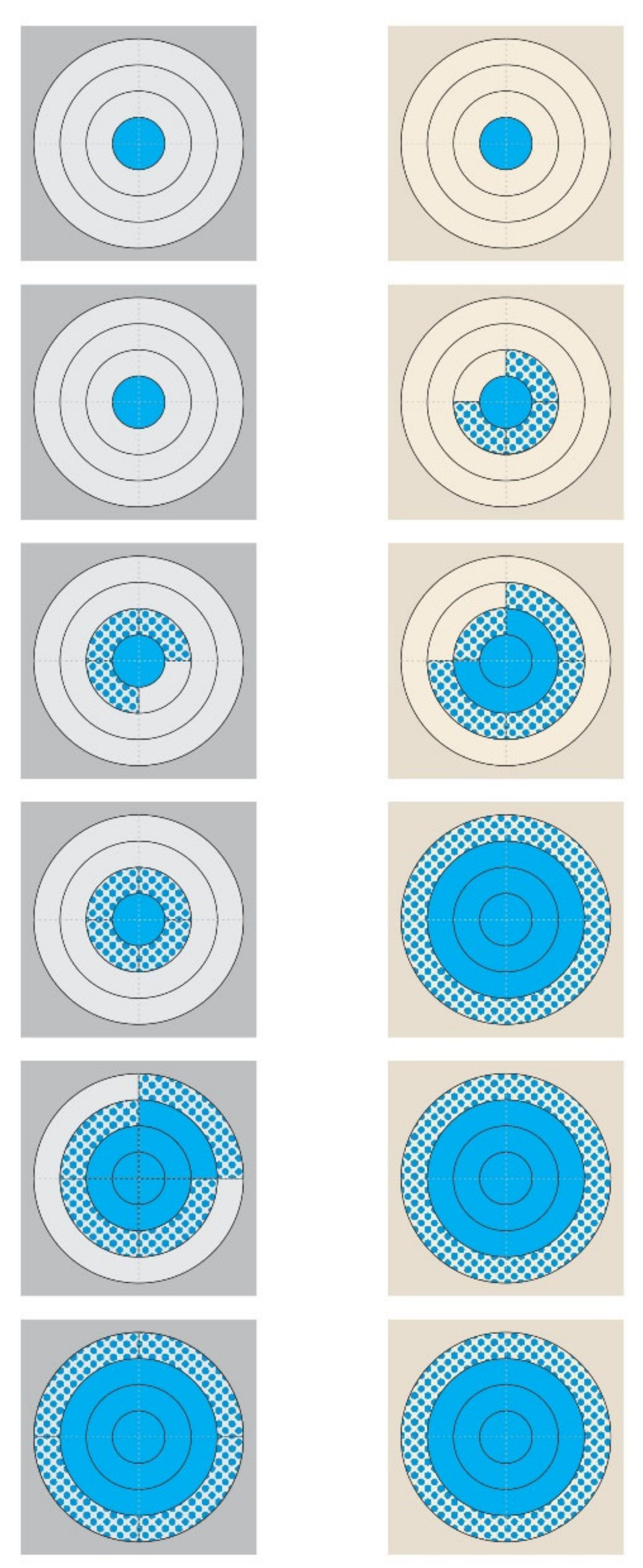
- ‘Isolation of staphylococci from the piece of teatcup material’ was equivalent to ‘presence of staphylococci on the piece of teatcup material’ on the respective quadrant that was sampled;
- A quadrant of a circular zone was considered to have been contaminated with staphylococci, if the bacteria were isolated from a swab sample;
- If no staphylococci were isolated on one sampling point from a quadrant and then isolated on the next sampling point, the contamination of the quadrant was deemed to have taken place halfway between the two sampling points;
- A quadrant of each circular zone was considered to have been fully covered with staphylococci if the bacteria were isolated from the respective concentric quadrant immediately outside it; for example, a quadrant in circular zone 1 was considered to have been fully covered if staphylococci were recovered from the respective quadrant in circular zone 2;
- In view of the design of the complex, it was possible to evaluate the full coverage of the two inner circular zones only (circular zone 1 and circular zone 2 and their combination);
- If staphylococci were not isolated on one sampling point from a quadrant and then isolated on the next sampling point, then the full coverage of the respective concentric quadrant immediately inside it was deemed to have taken place halfway between the two sampling points; for example, when staphylococci were recovered from a quadrant in circular zone 3, it was considered that the respective quadrant in circular zone 2 had been fully covered.
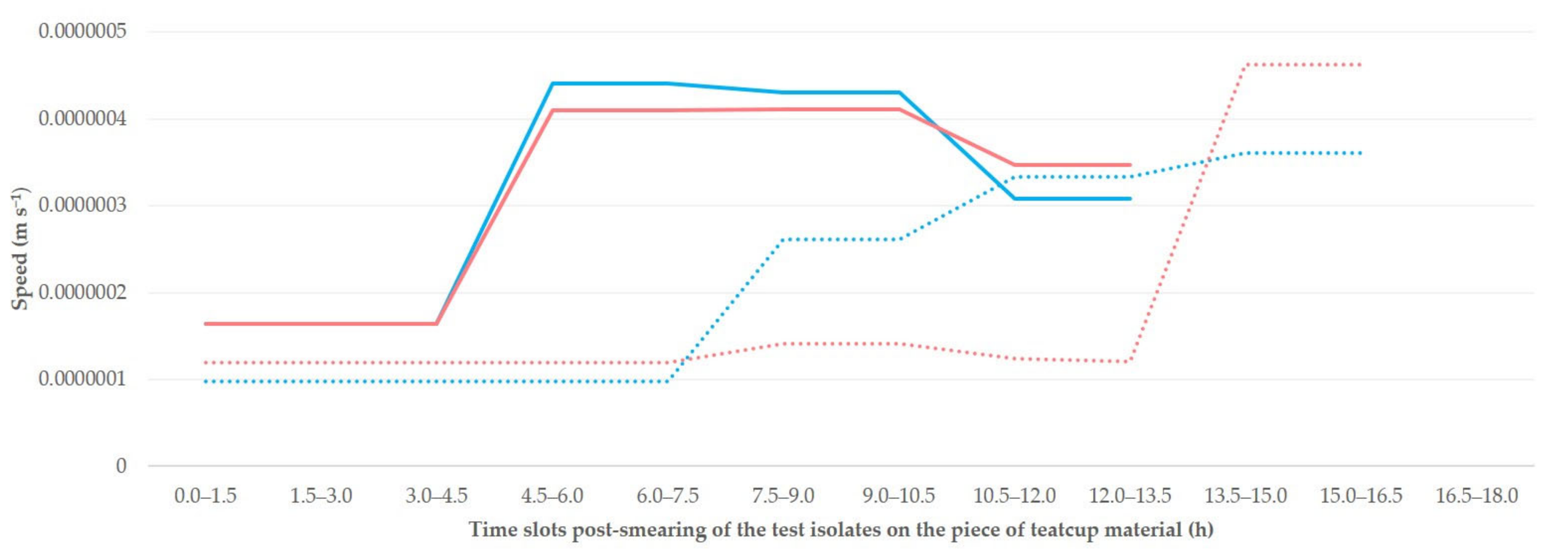
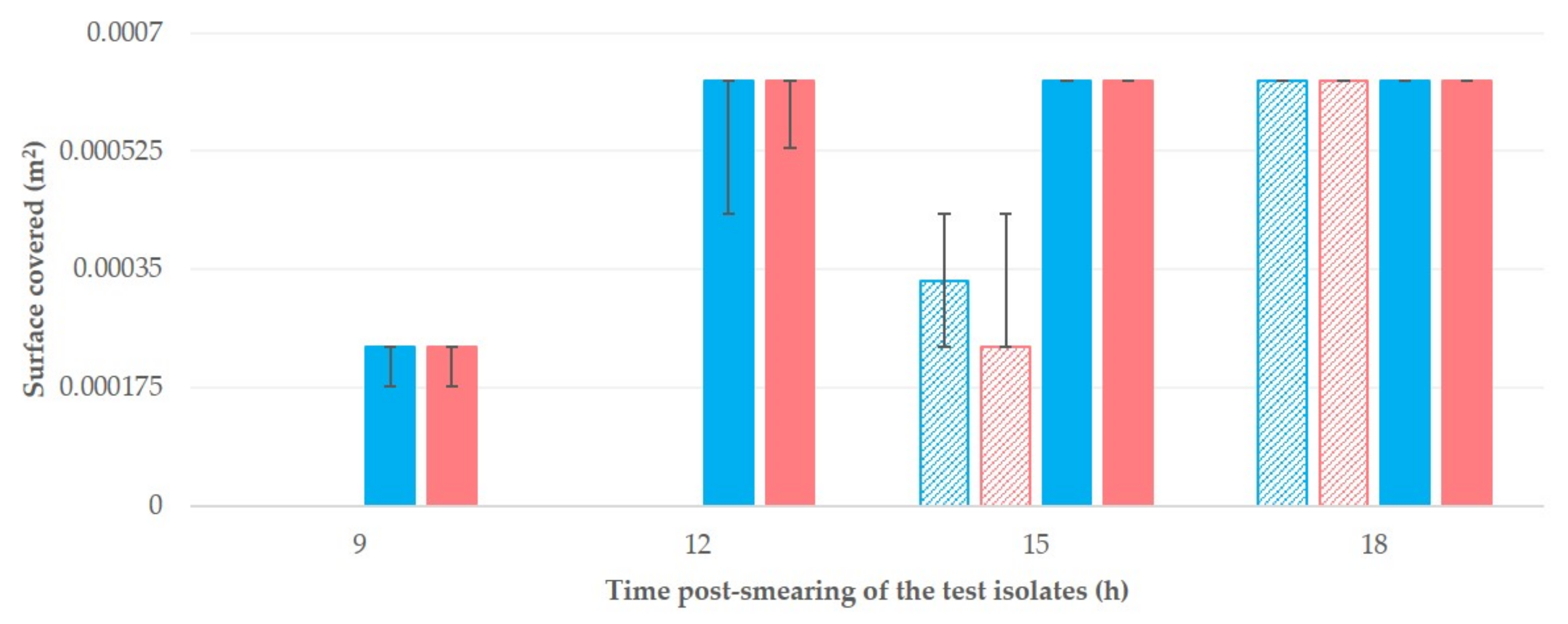
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Emori, T.G.; Gaynes, R.P. An overview of nosocomial infections, including the role of the microbiology laboratory. Clin. Microbiol. Rev. 1993, 6, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Legakis, I.; Barbouni, A.; Legakis, N. The impact of nosocomial (healthcare-acquired) infections on public health. Sci. Chron. 2020, 25, 643–654. [Google Scholar]
- Gkana, E.; Lianou, A.; Nychas, G-J.E. Transfer of Salmonella enterica ser. Typhimurium from beef to tomato through kitchen equipment and the efficacy of intermediate cleaning procedures. J. Food Prot. 2016, 79, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Michael, C.K.; Lianou, D.T.; Vasileiou, N.G.C.; Tsilipounidaki, K.; Katsafadou, A.I.; Politis, A.I.; Kordalis, N.G.; Ioannidi, K.S.; Gougoulis, D.A.; Trikalinou, C.; et al. Association of staphylococcal populations on teatcups of milking rarlours with vaccination against staphylococcal mastitis in sheep and goat farms. Pathogens 2021, 10, 385. [Google Scholar] [CrossRef] [PubMed]
- Bremer, P.J.; Fillery, S.; McQuillan, A.J. Laboratory scale Clean-In-Place (CIP) studies on the effectiveness of different caustic and acid wash steps on the removal of dairy biofilms. Int. J. Food Microbiol. 2006, 106, 254–262. [Google Scholar] [CrossRef]
- Fragkou, I.A.; Papaioannou, N.; Cripps, P.J.; Boscos, C.M.; Fthenakis, G.C. Teat lesions predispose to invasion of the ovine mammary gland by Mannheimia haemolytica. J. Comp. Pathol. 2007, 139, 239–244. [Google Scholar] [CrossRef]
- Romero, G.; Peris, C.; Fthenakis, G.C.; Diaz, J.R. Effects of machine milking on udder health in dairy ewes. Small Rumin. Res. 2020, 188, 106096. [Google Scholar] [CrossRef]
- Vasileiou, N.G.C.; Cripps, P.J.; Ioannidi, K.S.; Chatzopoulos, D.C.; Gougoulis, D.A.; Sarrou, S.; Orfanou, D.C.; Politis, A.; Calvo Gonzalez-Valerio, T.; Argyros, S.; et al. Extensive countrywide field investigation of subclinical mastitis in sheep in Greece. J. Dairy Sci. 2018, 101, 7297–7310. [Google Scholar] [CrossRef] [Green Version]
- Fragkou, I.A.; Boscos, C.M.; Fthenakis, G.C. Diagnosis of clinical or subclinical mastitis in ewes. Small Rumin. Res. 2014, 118, 86–92. [Google Scholar] [CrossRef]
- Katsarou, E.I.; Chatzopoulos, D.C.; Giannoulis, T.; Ioannidi, K.S.; Katsafadou, A.I.; Kontou, P.I.; Lianou, D.T.; Mamuris, Z.; Mavrogianni, V.S.; Michael, C.K.; et al. MLST-based analysis and antimicrobial resistance of Staphylococcus epidermidis from cases of sheep mastitis in Greece. Biology 2021, 10, 170. [Google Scholar] [CrossRef]
- Vasileiou, N.G.C.; Chatzopoulos, D.C.; Gougoulis, D.A.; Sarrou, S.; Katsafadou, A.I.; Spyrou, V.; Mavrogianni, V.S.; Petinaki, E.; Fthenakis, G.C. Slime-producing staphylococci as causal agents of subclinical mastitis in sheep. Vet. Microbiol. 2018, 224, 93–99. [Google Scholar] [CrossRef]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method of detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. J. Hyg. Camb. 1938, 38, 732–749. [Google Scholar] [CrossRef] [Green Version]
- Barrow, G.I.; Feltham, R.K.A. Manual for the Identification of Medical Bacteria, 3rd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Euzeby, J.P. List of bacterial names with standing in nomenclature: A folder available on the Internet. Int. J. Syst. Bacteriol. 1997, 47, 590–592. [Google Scholar] [CrossRef] [Green Version]
- Henrichsen, J. Bacterial surface translocation: A survey and a classification. Bacteriol. Rev. 1972, 36, 478–503. [Google Scholar] [CrossRef]
- Pollitt, E.J.G.; Diggle, S.P. Defining motility in the Staphylococci. Cell. Mol. Life Sci. 2017, 74, 2943–2958. [Google Scholar] [CrossRef] [Green Version]
- Dordet-Frisoni, E.; Gaillard-Martinie, B.; Talon, R.; Leroy, S. Surface migration of Staphylococcus xylosus on low-agar media. Res. Microbiol. 2008, 159, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.; Azeredo, J.; Teixeira, P. The importance of physicochemical properties in biofilm formation and activity. In Biofilms in Wastewater Treatment: An Interdisciplinary Approach; Wuertz, S., Bishop, P.L., Wilderer, P.A., Eds.; International Water Association Publications: London, UK, 2003; pp. 211–231. [Google Scholar]
- Busscher, H.J.; Sjollema, J.; Van der Mei, H. Relative importance of surface free energy as a measure of hydrophobicity in bacterial adhesion to solid surfaces. In Microbial Cell Surface Hydrophobicity; Doyle, R.J., Rosenberg, M., Eds.; American Society for Microbiology: Washington, DC, USA, 1990; pp. 333–359. [Google Scholar]
- Oliveira, R.; Azeredo, J.; Teixeira, P.; Fonseca, A.P. The role of hydrophobicity in bacterial adhesion. In Biofilm Community Interactions: Chance or Necessity; Gilbert, P., Allison, D., Brading, M., Verran, J., Walker, J., Eds.; Bioline: Cardiff, UK, 2001; pp. 11–22. [Google Scholar]
- Van Loosdrecht, M.C.M.; Norde, W.; Lyklema, J.; Zehnder, A.J.B. Hydrophobic and electrostatic parameters in bacterial adhesion. Aquatic Sci. 1990, 52, 103–114. [Google Scholar] [CrossRef]
- Lin, M.H.; Ke, W.J.; Liu, C.C.; Yang, M.W. Modulation of Staphylococcus aureus spreading by water. Sci. Rep. 2016, 6, 25233. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M. Biofilm formation: A clinically relevant microbiological process. Clin. Infect. Dis. 2001, 33, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Rose, K.; Steinbüchel, A. Biodegradation of natural rubber and related compounds: Recent insights into a hardly understood catabolic capability of microorganisms. Appl. Environ. Microbiol. 2005, 71, 2803–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salomez, M.; Subileau, M.; Intapun, J.; Bonfils, F.; Sainte-Beuve, J.; Vaysse, L.; Dubreucq, E. Micro-organisms in latex and natural rubber coagula of Hevea brasiliensis and their impact on rubber composition, structure and properties. J. Appl. Microbiol. 2014, 117, 921–929. [Google Scholar] [CrossRef]
- Salomez, M.; Subileau, M.; Vallaeys, T.; Santoni, S.; Bonfils, F.; Sainte-Beuve, J.; Intapun, J.; Granet, F.; Vaysse, L.; Dubreucq, E. Microbial communities in natural rubber coagula during maturation: Impacts on technological properties of dry natural rubber. J. Appl. Microbiol. 2018, 124, 444–456. [Google Scholar] [CrossRef]
- Sousa, C.; Teixeira, P.; Oliveira, R. Influence of surface properties on the adhesion of Staphylococcus epidermidis to acrylic and silicone. Int. J. Biomater. 2009, 2009, 718017. [Google Scholar] [CrossRef] [Green Version]
- Kodjikian, L.; Burillon, C.; Chanloy, C.; Bostvironnois, C.; Pellon, G.; Mari, E.; Freney, J.; Roger, T. In vivo study of bacterial adhesion to five types of intraocular lenses. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3717–3721. [Google Scholar]
- Kim, S.E.; Lee, S.J.; Lee, S.Y.; Yoon, J.S. Clinical significance of microbial growth on the surfaces of silicone tubes removed fromdacryocystorhinostomy patients. Am. J. Ophthalmol. 2012, 153, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Gelasakis, A.I.; Mavrogianni, V.S.; Petridis, I.G.; Vasileiou, N.G.C.; Fthenakis, G.C. Mastitis in sheep—The last 10 years and the future of research. Vet. Microbiol. 2015, 185, 136–146. [Google Scholar] [CrossRef]
- Contreras, G.A.; Rodríguez, J.M. Mastitis: Comparative etiology and epidemiology. J. Mammary Gland Biol. Neoplasia 2011, 16, 339–356. [Google Scholar] [CrossRef]
- Erskine, R.J. Mastitis in cattle. In Merck Veterinary Manual; Winter, A.L., Ed.; MSD Publishing Group, Merck & Co. Inc.: Kenilworth, NJ, USA, 2020. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after Smearing | Total Recoveries | |
|---|---|---|
| Isolate A | Isolate B | |
| Teatcups for cattle | ||
| 3 h | 0/108 | 0/108 |
| 6 h | 0/108 | 0/108 |
| 9 h | 32/108 | 33/108 |
| 12 h | 36/108 | 36/108 |
| 15 h | 80/108 | 77/108 |
| 18 h | 108/108 | 108/108 |
| 21 h | 108/108 | 108/108 |
| 24 h | 108/108 | 108/108 |
| Total | 472/864 | 470/864 |
| Teatcups for sheep | ||
| 3 h | 0/108 | 0/108 |
| 6 h | 33/108 | 33/108 |
| 9 h | 71/108 | 71/108 |
| 12 h | 102/108 | 106/108 |
| 15 h | 108/108 | 108/108 |
| 18 h | 108/108 | 108/108 |
| 21 h | 108/108 | 108/108 |
| 24 h | 108/108 | 108/108 |
| Total | 638/864 | 642/864 |
| Grand total | 1110/1728 | 1112/1728 |
| Time after Smearing | Isolate A | Isolate B |
|---|---|---|
| Teatcups for cattle | ||
| 9 h | 0.000000 a (0.000000–0.000000) | 0.00000 a (0.00000–0.00000) |
| 12 h | 0.000000 b (0.00000–0.000000) | 0.00000 b (0.00000–0.00000) |
| 15 h | 0.000334 c (0.000236–0.000432) | 0.000236 c (0.000236–0.000432) |
| 18 h | 0.000628 (0.000628–0.000628) | 0.000628 (0.000628–0.000628) |
| Teatcups for sheep | ||
| 9 h | 0.000236 a (0.000177–0.000236) | 0.000236 a (0.000177–0.000432) |
| 12 h | 0.000628 b (0.000628–0.000628) | 0.000628 b (0.000530–0.000628) |
| 15 h | 0.000628 c (0.000628–0.000628) | 0.000628 c (0.000628–0.000628) |
| 18 h | 0.000628 (0.000628–0.000628) | 0.000628 (0.000628–0.000628) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsarou, E.I.; Katsafadou, A.I.; Karakasidis, T.; Chatzopoulos, D.C.; Vasileiou, N.G.C.; Lianou, D.T.; Mavrogianni, V.S.; Petinaki, E.; Fthenakis, G.C. Growth of Staphylococcus epidermidis on the Surface of Teatcups from Milking Parlours. Microorganisms 2021, 9, 852. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040852
Katsarou EI, Katsafadou AI, Karakasidis T, Chatzopoulos DC, Vasileiou NGC, Lianou DT, Mavrogianni VS, Petinaki E, Fthenakis GC. Growth of Staphylococcus epidermidis on the Surface of Teatcups from Milking Parlours. Microorganisms. 2021; 9(4):852. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040852
Chicago/Turabian StyleKatsarou, Eleni I., Angeliki I. Katsafadou, Theodoros Karakasidis, Dimitris C. Chatzopoulos, Natalia G. C. Vasileiou, Daphne T. Lianou, Vasia S. Mavrogianni, Efthymia Petinaki, and George C. Fthenakis. 2021. "Growth of Staphylococcus epidermidis on the Surface of Teatcups from Milking Parlours" Microorganisms 9, no. 4: 852. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040852