
Classification of Salmonella enterica of the (Para-)Typhoid Fever Group by Fourier-Transform Infrared (FTIR) Spectroscopy

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Collection
2.2. Sample Preparation
2.3. Spectra Acquisition and Analysis
2.4. Cluster Analysis
2.5. Development of Automated Classifiers
2.6. Ethical Clearance
3. Results
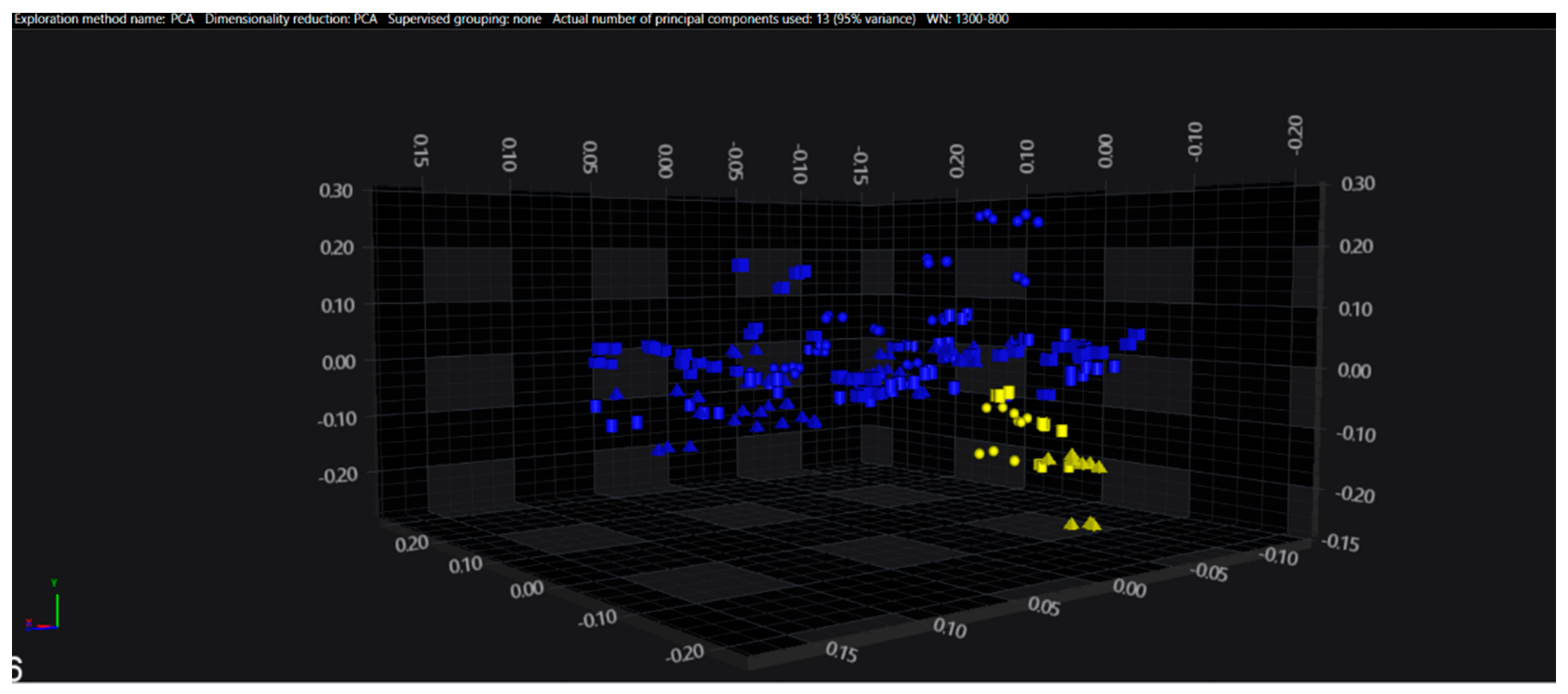
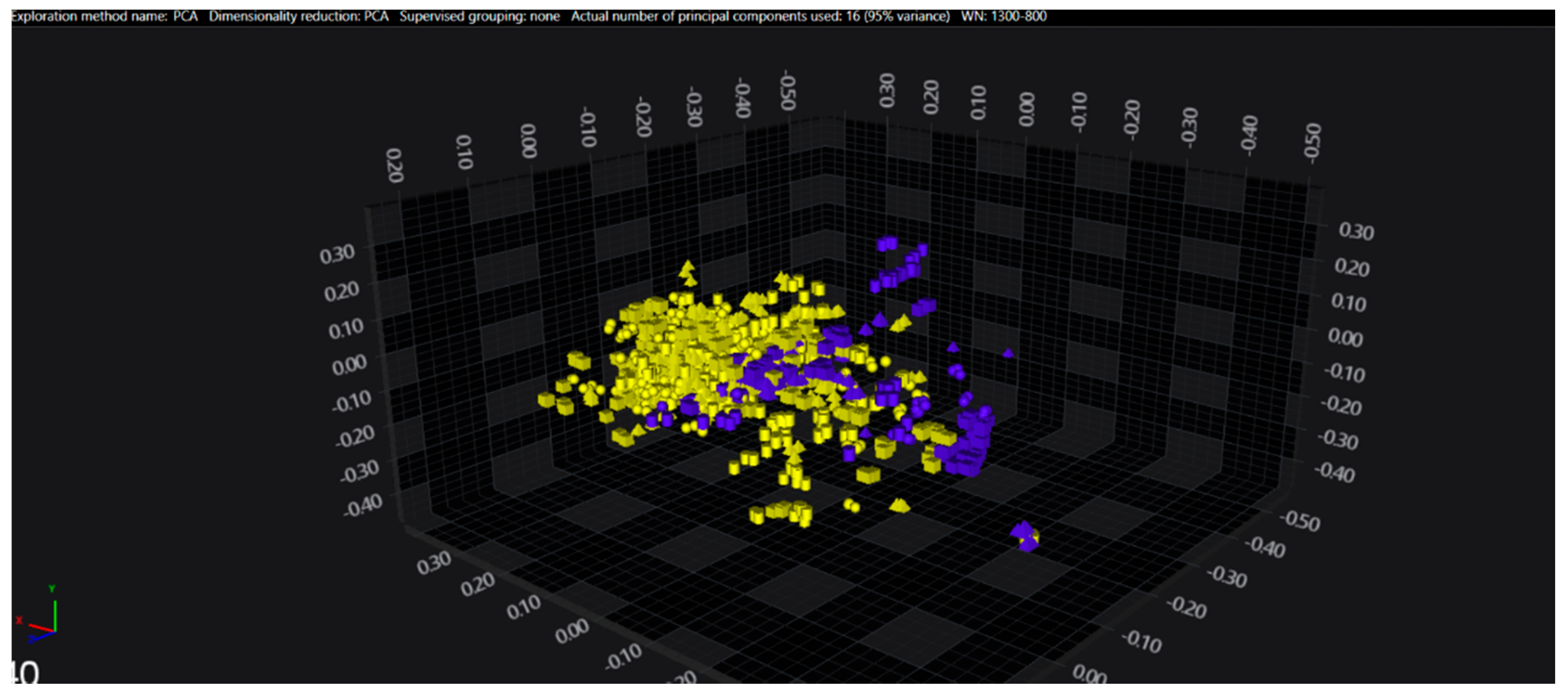
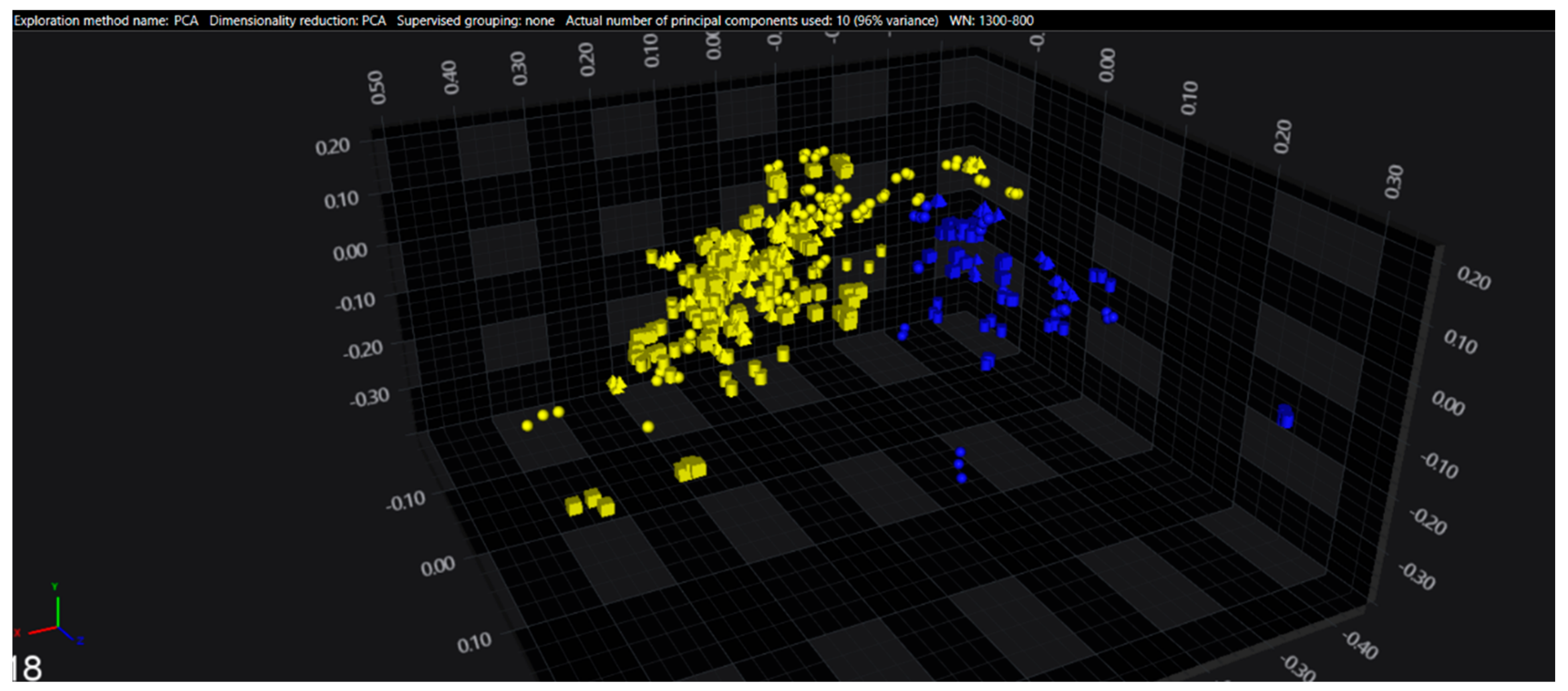
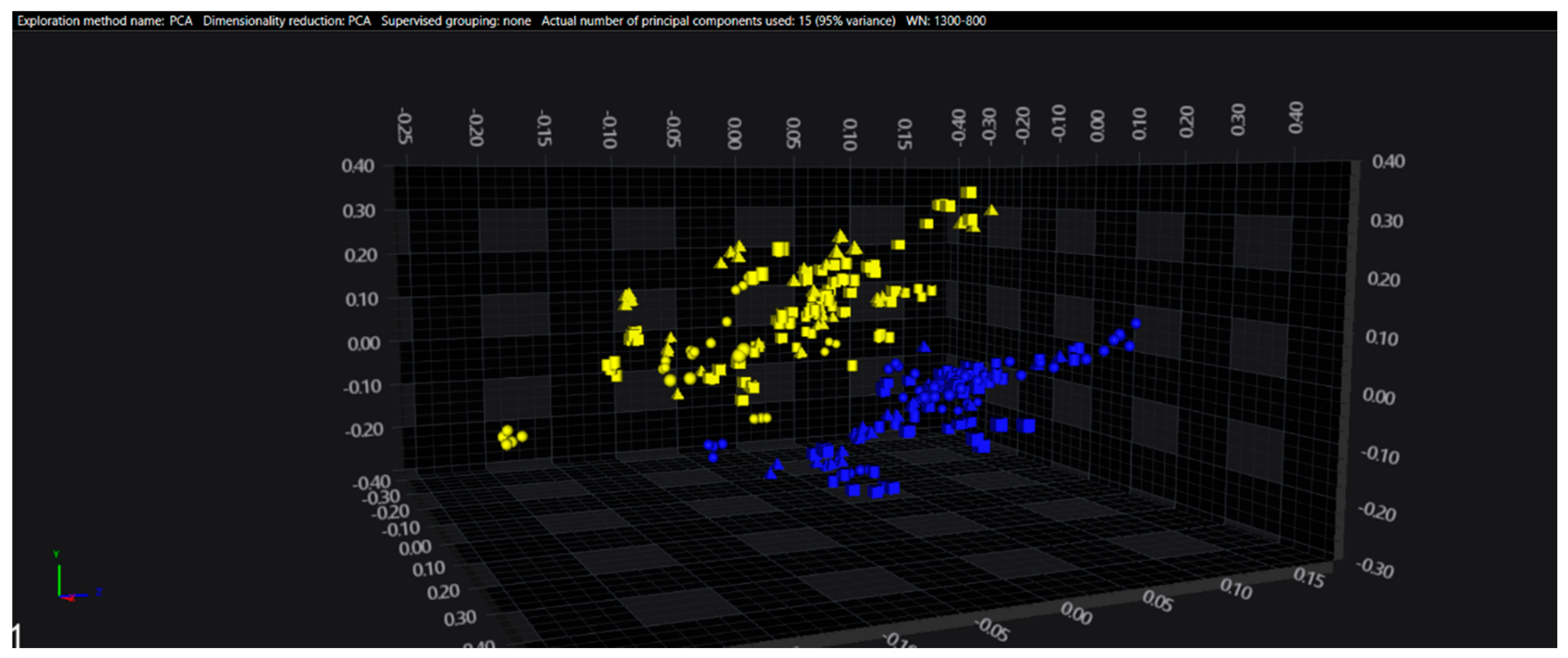
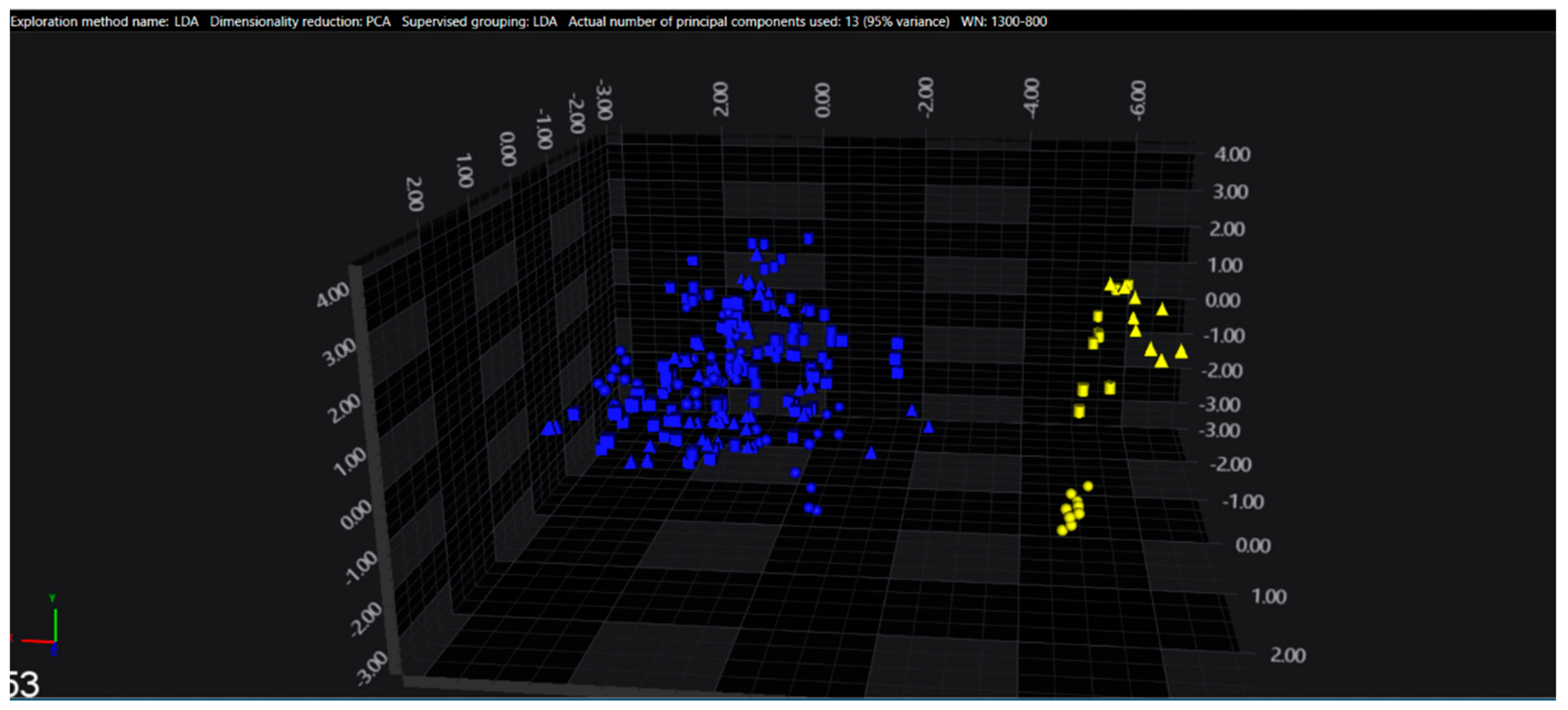
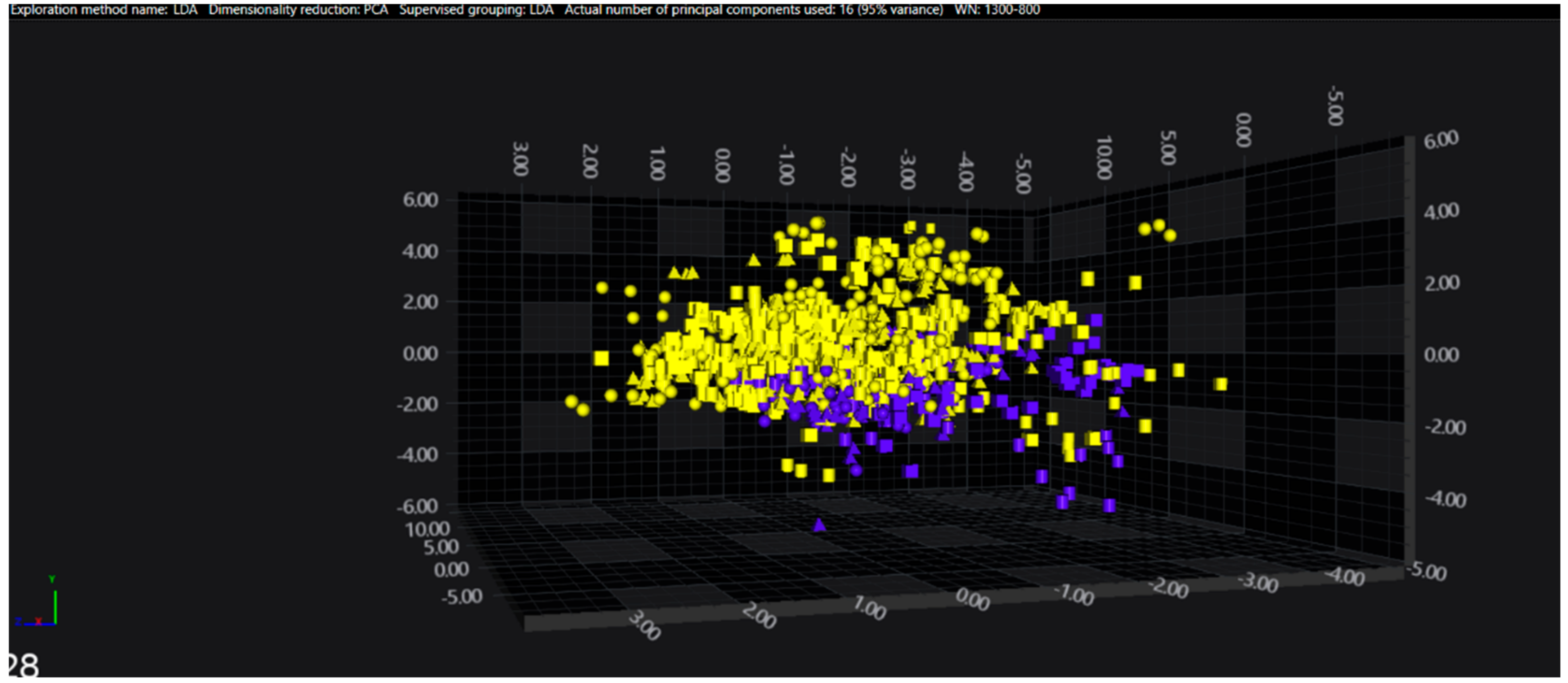
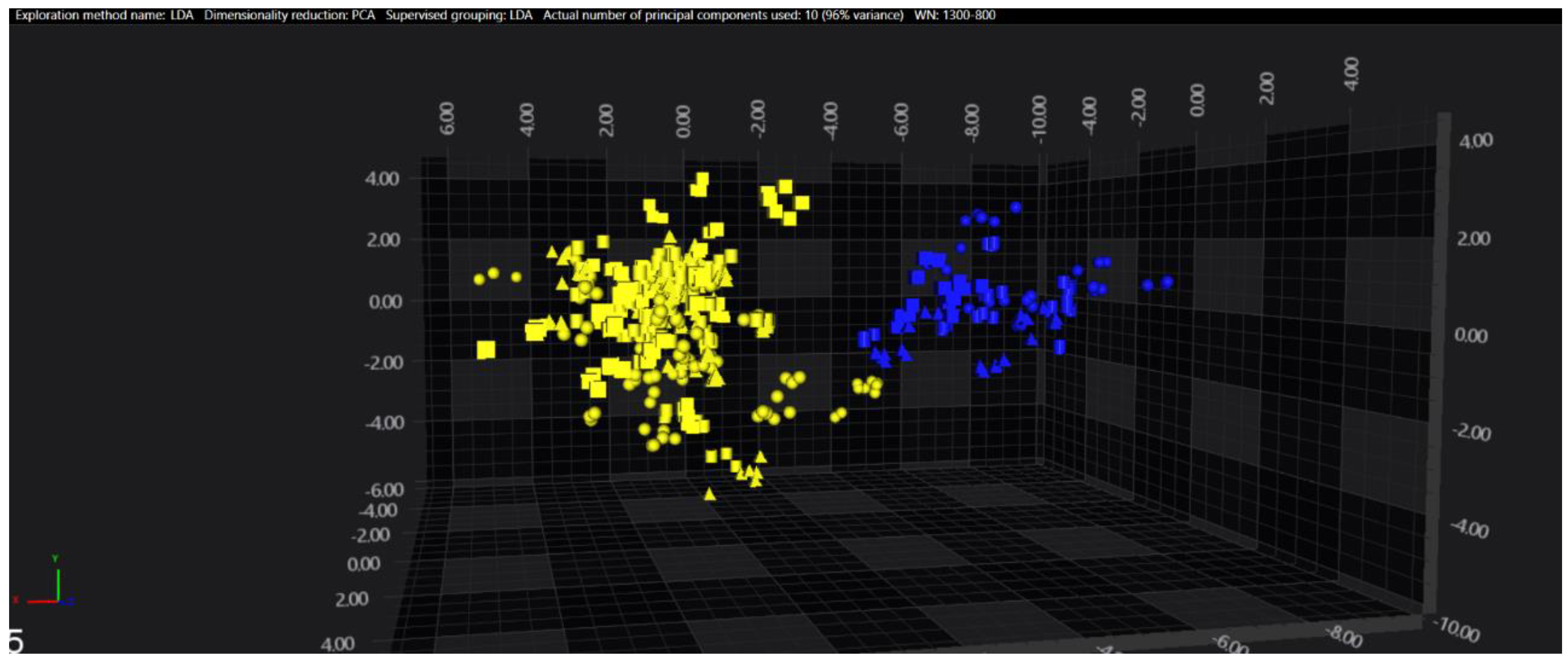
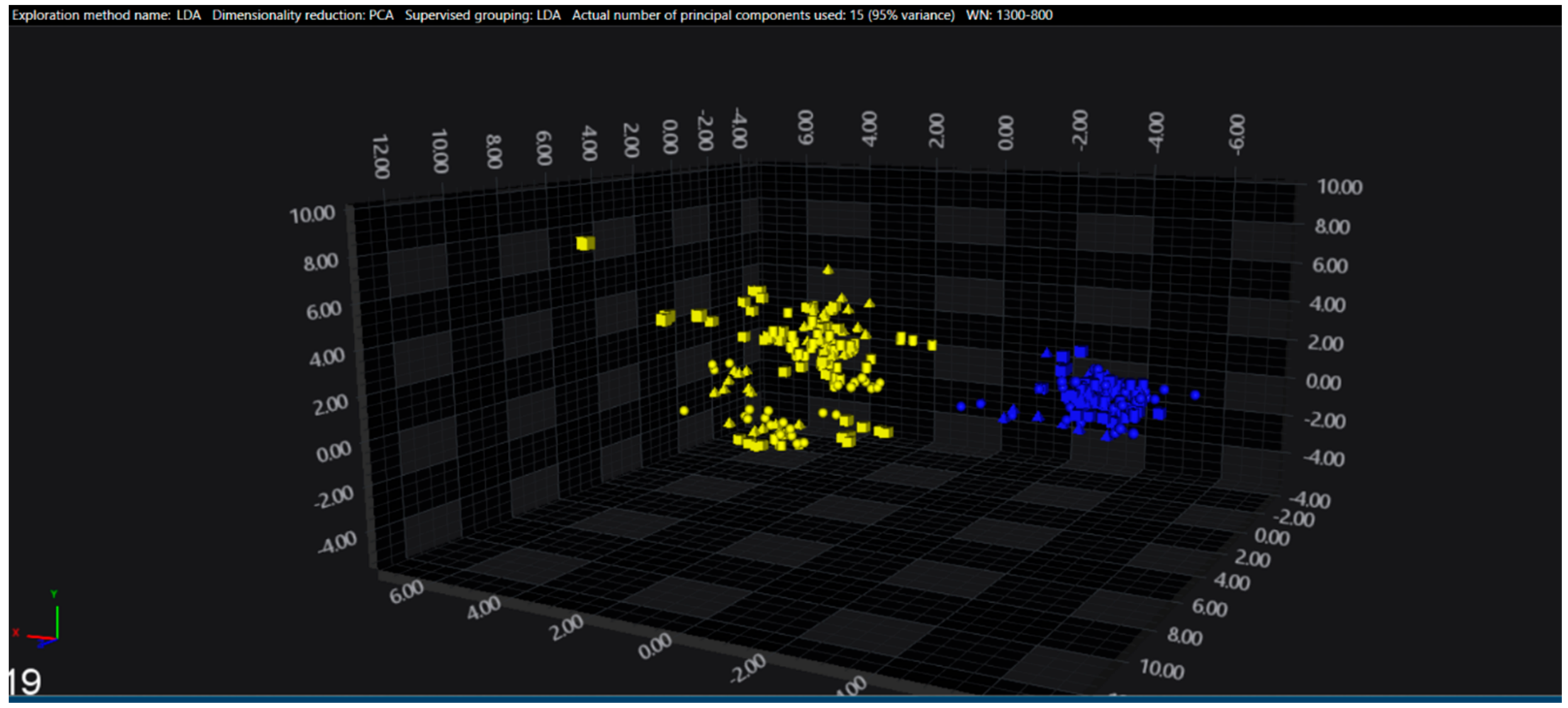
3.1. Cluster Analysis
3.2. Automated Classifiers
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diekema, D.J.; Hsueh, P.R.; Mendes, R.E.; Pfaller, M.A.; Rolston, K.V.; Sader, H.S.; Jones, R.N. The Microbiology of Bloodstream Infection: 20-Year Trends from the SENTRY Antimicrobial Surveillance Program. Antimicrob. Agents. Chemother. 2019, 63, e00355-19. [Google Scholar] [CrossRef] [Green Version]
- Morpeth, S.C.; Ramadhani, H.O.; Crump, J.A. Invasive non-Typhi Salmonella disease in Africa. Clin. Infect. Dis. 2009, 49, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Reddy, E.A.; Shaw, A.V.; Crump, J.A. Community-acquired bloodstream infections in Africa: A systematic review and meta-analysis. Lancet Infect. Dis. 2010, 10, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Gordon, M.A. Invasive nontyphoidal Salmonella disease: Epidemiology, pathogenesis and diagnosis. Curr. Opin. Infect. Dis. 2011, 24, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Deen, J.; von Seidlein, L.; Andersen, F.; Elle, N.; White, N.J.; Lubell, Y. Community-acquired bacterial bloodstream infections in developing countries in south and southeast Asia: A systematic review. Lancet Infect. Dis. 2012, 12, 480–487. [Google Scholar] [CrossRef]
- Fierer, J.; Swancutt, M. Non-typhoid Salmonella: A review. Curr. Clin. Top. Infect. Dis. 2000, 20, 134–157. [Google Scholar]
- Wain, J.; Hendriksen, R.S.; Mikoleit, M.L.; Keddy, K.H.; Ochiai, R.L. Typhoid fever. Lancet. 2015, 385, 1136–1145. [Google Scholar] [CrossRef]
- Dougan, G.; Baker, S. Salmonella enterica serovar Typhi and the pathogenesis of typhoid fever. Annu. Rev. Microbiol. 2014, 68, 317–336. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Im, J.; Parajulee, P.; Holm, M.; Cruz Espinoza, L.M.; Poudyal, N.; Mogeni, O.D.; Marks, F. A Systematic Review of Typhoid Fever Occurrence in Africa. Clin. Infect. Dis. 2019, 69 (Suppl. 6), S492–S498. [Google Scholar] [CrossRef]
- Mutua, J.M.; Wang, F.B.; Vaidya, N.K. Modeling malaria and typhoid fever co-infection dynamics. Math. Biosci. 2015, 264, 128–144. [Google Scholar] [CrossRef]
- Gibani, M.M.; Britto, C.; Pollard, A.J. Typhoid and paratyphoid fever: A call to action. Curr. Opin. Infect. Dis. 2018, 31, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Fangtham, M.; Wilde, H. Emergence of Salmonella paratyphi A as a major cause of enteric fever: Need for early detection, preventive measures, and effective vaccines. J. Travel Med. 2008, 15, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Sahastrabuddhe, S.; Carbis, R.; Wierzba, T.F.; Ochiai, R.L. Increasing rates of Salmonella Paratyphi A and the current status of its vaccine development. Expert. Rev. Vaccines 2013, 12, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Arndt, M.B.; Mosites, E.M.; Tian, M.; Forouzanfar, M.H.; Mokhdad, A.H.; Meller, M.; Ochiai, R.L.; Walson, J.L. Estimating the burden of paratyphoid a in Asia and Africa. PLoS Negl. Trop. Dis. 2014, 8, e2925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, M.R.; Koornhof, H.J.; Crisp, S.I.; Palmhert, H.L.; Fitzstephens, A. Enteric fever caused by Salmonella paratyphi C in South and South West Africa. S. Afr. Med. J. 1978, 54, 434–438. [Google Scholar]
- Fuche, F.J.; Sen, S.; Jones, J.A.; Nkeze, J.; Permala-Booth, J.; Tapia, M.D.; Sow, S.O.; Tamboura, B.; Touré, A.; Onwuchekwa, U.; et al. Characterization of Invasive Salmonella Serogroup C1 Infections in Mali. Am. J. Trop. Med. Hyg. 2018, 98, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Luby, S.P.; Saha, S.; Andrews, J.R. Towards sustainable public health surveillance for enteric fever. Vaccine. 2015, 33 (Suppl. 3), C3–C7. [Google Scholar] [CrossRef]
- Selander, R.K.; Beltran, P.; Smith, N.H.; Barker, R.M.; Crichton, P.B.; Old, D.C.; Musser, J.M.; Whittam, T.S. Genetic population structure, clonal phylogeny, and pathogenicity of Salmonella paratyphi B. Infect. Immun. 1990, 58, 1891–1901. [Google Scholar] [CrossRef] [Green Version]
- Huehn, S.; Helmuth, R.; Bunge, C.; Guerra, B.; Junker, E.; Davies, R.H.; Wattiau, P.; van Pelt, W.; Malorny, B. Characterization of pathogenic and resistant genome repertoire reveals two clonal lines in Salmonella enterica subsp. enterica serovar Paratyphi B (+)-tartrate positive. Foodborne Pathog. Dis. 2009, 6, 431–443. [Google Scholar] [CrossRef]
- Barker, R.M.; Kearney, G.M.; Nicholson, P.; Blair, A.L.; Porter, R.C.; Crichton, P.B. Types of Salmonella paratyphi B and their phylogenetic significance. J. Med. Microbiol. 1988, 26, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doublet, B.; Praud, K.; Nguyen-Ho-Bao, T.; Argudín, M.A.; Bertrand, S.; Butaye, P.; Cloeckaert, A. Extended-spectrum β-lactamase- and AmpC β-lactamase-producing D-tartrate-positive Salmonella enterica serovar Paratyphi B from broilers and human patients in Belgium, 2008–2010. J. Antimicrob. Chemother. 2014, 69, 1257–1264. [Google Scholar] [CrossRef] [Green Version]
- Toboldt, A.; Tietze, E.; Helmuth, R.; Fruth, A.; Junker, E.; Malorny, B. Human infections attributable to the D-tartrate-fermenting variant of Salmonella enterica serovar Paratyphi B in Germany originate in reptiles and, on rare occasions, poultry. Appl. Environ. Microbiol. 2012, 78, 7347–7357. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.R.; Ryan, E.T. Diagnostics for invasive Salmonella infections: Current challenges and future directions. Vaccine 2015, 33 (Suppl. 3), C8–C15. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Ray, U.; Akhter, I.; Chattopadhyay, A.; Paul, D.K.; Dutta, S. Evaluation of fliC-d based direct blood PCR assays for typhoid diagnosis. BMC Microbiol. 2016, 16, 108. [Google Scholar] [CrossRef] [Green Version]
- Tennant, S.M.; Toema, D.; Qamar, F.; Iqbal, N.; Boyd, M.A.; Marshall, J.M.; Blackwelder, W.C.; Wu, Y.; Quadri, F.; Khan, A.; et al. Detection of Typhoidal and Paratyphoidal Salmonella in Blood by Real-time Polymerase Chain Reaction. Clin. Infect. Dis. 2015, 61 (Suppl. 4), S241–S250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, K.; Itoh, K.; Nakajima, H.; Kurazono, T.; Yamaguchi, M.; Moriya, K.; Ezaki, T.; Kawamura, Y.; Tamura, K.; Watanabe, H. Selective amplification of tyv (rfbE), prt (rfbS), viaB, and fliC genes by multiplex PCR for identification of Salmonella enterica serovars Typhi and Paratyphi, A.J. Clin. Microbiol. 2002, 40, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.F.; Reen, F.J.; Gilroy, D.; Buckley, J.; Frye, J.G.; Boyd, E.F. Rapid multiplex PCR and real-time TaqMan PCR assays for detection of Salmonella enterica and the highly virulent serovars Choleraesuis and Paratyphi, C.J. Clin. Microbiol. 2008, 46, 4018–4022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngan, G.J.; Ng, L.M.; Lin, R.T.; Teo, J.W. Development of a novel multiplex PCR for the detection and differentiation of Salmonella enterica serovars Typhi and Paratyphi, A. Res. Microbiol. 2010, 161, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Nga, T.V.; Karkey, A.; Dongol, S.; Thuy, H.N.; Dunstan, S.; Holt, K.; Tu le, T.P.; Campbell, J.I.; Chau, T.T.; Chau, N.V.; et al. The sensitivity of real-time PCR amplification targeting invasive Salmonella serovars in biological specimens. BMC Infect. Dis. 2010, 10, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Shi, X.; Zhu, Y.; Qiu, Y.; Li, Y.; Lv, D.; Hu, Q. Development of modified molecular beacon based real-time PCR assay for the rapid detection of Salmonella choleraesuis and Salmonella paratyphi C. Wei Sheng Yan Jiu 2011, 40, 514–516. [Google Scholar]
- Xu, L.; Yuan, M.; Sun, W.; Ke, B.; Hong, Y. Taq Man probe-based quadruple real-time PCR for detection of Salmonella paratyphi A/B/C and Salmonella typhi. Wei Sheng Yan Jiu 2017, 46, 298–302. [Google Scholar]
- Zhai, L.; Yu, Q.; Bie, X.; Lu, Z.; Lv, F.; Zhang, C.; Kong, X.; Zhao, H. Development of a PCR test system for specific detection of Salmonella Paratyphi B in foods. FEMS Microbiol. Lett. 2014, 355, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, J.; Saffie, N.; Sjasri, F.A.; Husin, A.; Abdul-Rahman, Z.; Ismail, A.; Aziah, I.; Mohamed, M. Rapid detection of Salmonella Typhi by loop-mediated isothermal amplification (LAMP) method. Braz. J. Microbiol. 2015, 45, 1385–1391. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.; Du, P.; Kan, B.; Yan, M. The Development and Evaluation of a Loop-Mediated Isothermal Amplification Method for the Rapid Detection of Salmonella enterica serovar Typhi. PLoS ONE 2015, 10, e0124507. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.; Yan, M.; Du, P.; Chen, C.; Kan, B. Rapid and Sensitive Salmonella Typhi Detection in Blood and Fecal Samples Using Reverse Transcription Loop-Mediated Isothermal Amplification. Foodborne Pathog. Dis. 2015, 12, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Das, R.; Nigam, M.R.; Elangovan, R.; Pandya, D.; Jha, S.; Kalyanasundaram, D. Rapid Detection Device for Salmonella typhi in Milk, Juice, Water and Calf Serum. Indian. J. Microbiol. 2018, 58, 381–392. [Google Scholar]
- Kaur, A.; Kapil, A.; Elangovan, R.; Jha, S.; Kalyanasundaram, D. Highly-sensitive detection of Salmonella typhi in clinical blood samples by magnetic nanoparticle-based enrichment and in-situ measurement of isothermal amplification of nucleic acids. PLoS ONE 2018, 13, e0194817. [Google Scholar]
- Rojak, S.; Wiemer, D.F.; Wille, A.; Loderstädt, U.; Wassill, L.; Hinz, R.; Hagen, R.M.; Frickmann, H. Loop-mediated isothermal amplification for paratyphoid fever—A proof-of-principle analysis. Lett. Appl. Microbiol. 2019, 68, 509–513. [Google Scholar] [CrossRef]
- Frickmann, H.; Wiemer, D.F.; Wassill, L.; Hinz, R.; Rojak, S.; Wille, A.; Loderstädt, U.; Schwarz, N.G.; von Kalckreuth, V.; Im, J.; et al. Loop-mediated isothermal amplification-based detection of typhoid fever on an automated Genie II Mk2 system—A case-control-based approach. Acta Trop. 2019, 190, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Q.; Feng, Y.; Wang, Y.; Zou, Q.H.; Chen, F.; Guo, J.T.; Peng, Y.H.; Jin, Y.; Li, Y.G.; Hu, S.N.; et al. Salmonella paratyphi C: Genetic divergence from Salmonella choleraesuis and pathogenic convergence with Salmonella typhi. PLoS ONE 2009, 4, e4510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhns, M.; Zautner, A.E.; Rabsch, W.; Zimmermann, O.; Weig, M.; Bader, O.; Groß, U. Rapid discrimination of Salmonella enterica serovar Typhi from other serovars by MALDI-TOF mass spectrometry. PLoS ONE 2012, 7, e40004. [Google Scholar] [CrossRef]
- Baker, M.J.; Trevisan, J.; Bassan, P.; Bhargava, R.; Butler, H.J.; Dorling, K.M.; Fielden, P.R.; Fogarty, S.W.; Fullwood, N.J.; Heys, K.A.; et al. Using Fourier transform IR spectroscopy to analyze biological materials. Nat. Protoc. 2014, 1771–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumann, D.; Helm, D.; Labischinski, H. Microbiological characterization by FT-IR spectroscopy. Nature 1991, 351, 81–82. [Google Scholar] [CrossRef] [PubMed]
- Helm, D.; Labischinski, H.; Schallehn, G.; Naumann, D. Classification and identification of bacteria by Fourier-transform infrared spectroscopy. J. Gen. Microbiol. 1991, 137, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Lasch, P.; Naumann, D. Infrared spectroscopy in microbiology. In Encyclopedia of Analytical Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–32. [Google Scholar]
- Vogt, S.; Löffler, K.; Dinkelacker, A.G.; Bader, B.; Autenrieth, I.B.; Peter, S.; Liese, J. Fourier-Transform Infrared (FTIR) Spectroscopy for Typing of Clinical Enterobacter cloacae Complex Isolates. Front. Microbiol. 2019, 10, 2582. [Google Scholar] [CrossRef] [Green Version]
- Martak, D.; Valot, B.; Sauget, M.; Cholley, P.; Thouverez, M.; Bertrand, X.; Hocquet, D. Fourier-Transform InfraRed Spectroscopy Can Quickly Type Gram-Negative Bacilli Responsible for Hospital Outbreaks. Front. Microbiol. 2019, 10, 1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinkelacker, A.G.; Vogt, S.; Oberhettinger, P.; Mauder, N.; Rau, J.; Kostrzewa, M.; Rossen, J.W.A.; Autenrieth, I.B.; Peter, S.; Liese, J. Typing and Species Identification of Clinical Klebsiella Isolates by Fourier Transform Infrared Spectroscopy and Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2018, 56, e00843-18. [Google Scholar] [CrossRef] [Green Version]
- Burckhardt, I.; Sebastian, K.; Mauder, N.; Kostrzewa, M.; Burckhardt, F.; Zimmermann, S. Analysis of Streptococcus pneumoniae using Fourier-transformed infrared spectroscopy allows prediction of capsular serotype. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1883–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Jongmans-Hochschulz, E.; Mauder, N.; Imirzalioglu, C.; Wichels, A.; Gerdts, G. The Travelling Particles: Investigating microplastics as possible transport vectors for multidrug resistant E. coli in the Weser estuary (Germany). Sci. Total. Environ. 2020, 720, 137603. [Google Scholar] [CrossRef]
- Rodrigues, C.; Sousa, C.; Lopes, J.A.; Novais, Â.; Peixe, L. A Front Line on Klebsiella pneumoniae Capsular Polysaccharide Knowledge: Fourier Transform Infrared Spectroscopy as an Accurate and Fast Typing Tool. mSystems 2020, 5, e00386-19. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhou, H.; Lu, J.; Sun, Q.; Liu, C.; Zeng, Y.; Zhang, R. Evaluation of the IR Biotyper for Klebsiella pneumoniae typing and its potentials in hospital hygiene management. Microb. Biotechnol. 2020, 18. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.; Reuhs, B.L.; Mauer, L.J. Differentiation of Outer Membrane Proteins from Salmonella enterica Serotypes Using Fourier Transform Infrared Spectroscopy and Chemometrics. Lett. Appl. Microbiol. 2006, 42, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Männing, A.; Baldauf, N.A.; Rodriguez-Romo, L.A.; Yousef, A.E.; Rodríguez-Saona, L.E. Differentiation of Salmonella Enterica Serovars and Strains in Cultures and Food Using Infrared Spectroscopic and Microspectroscopic Techniques Combined With Soft Independent Modeling of Class Analogy Pattern Recognition Analysis. J. Food Prot. 2008, 71, 2249–2256. [Google Scholar] [CrossRef] [PubMed]
- De Lamo-Castellví, S.; Männing, A.; Rodríguez-Saona, L.E. Fourier-transform Infrared Spectroscopy Combined With Immunomagnetic Separation as a Tool to Discriminate Salmonella Serovars. Analyst 2010, 135, 2987–2992. [Google Scholar] [CrossRef]
- Preisner, O.; Guiomar, R.; Machado, J.; Menezes, J.C.; Lopes, J.A. Application of Fourier transform infrared spectroscopy and chemometrics for differentiation of Salmonella enterica serovar Enteritidis phage types. Appl. Environm. Microbiol. 2010, 76, 3538–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaram, J.; Park, B.; Hinton, A., Jr.; Yoon, S.C.; Windham, W.R.; Lawrence, K.C. Classification and structural analysis of live and dead Salmonella cells using Fourier transform infrared spectroscopy and principal component analysis. J. Agric. Food Chem. 2012, 60, 991–1004. [Google Scholar] [CrossRef]
- Campos, J.; Sousa, C.; Mourão, J.; Lopez, J.; Antunes, P.; Peixe, L. Discrimination of non-typhoid Salmonella serogroups and serotypes by Fourier Transform Infrared Spectroscopy: A comprehensive analysis. Int. J. Food Microbiol. 2018, 285, 34–41. [Google Scholar] [CrossRef]
- Grimont, P.A.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars; WHO Collaborating Center for Reference and Research on Salmonella: Paris, France, 2007. [Google Scholar]
- Ashton, P.M.; Nair, S.; Peters, T.M.; Bale, J.A.; Powell, D.G.; Painset, A.; Tewolde, R.; Schaefer, U.; Jenkins, C.; Dallman, T.J.; et al. Salmonella Whole Genome Sequencing Implementation Group. Identification of Salmonella for public health surveillance using whole genome sequencing. PeerJ 2016, 4, e1752. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, G.M.; Morin, P.M. Salmonella Serotyping Using Whole Genome Sequencing. Front. Microbiol. 2018, 9, 2993. [Google Scholar] [CrossRef] [Green Version]
- Yachison, C.A.; Yoshida, C.; Robertson, J.; Nash, J.H.E.; Kruczkiewicz, P.; Taboada, E.N.; Walker, M.; Reimer, A.; Christianson, S.; Nichani, A. PulseNet Canada Steering Committee, Nadon, C. The Validation and Implications of Using Whole Genome Sequencing as a Replacement for Traditional Serotyping for a National Salmonella Reference Laboratory. Front. Microbiol. 2017, 8, 1044. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barret, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on monitoring and assessment of the public health risk of “Salmonella Typhimurium-like” strains. EFSA J. 2010, 8, 1826. [Google Scholar] [CrossRef]
- Tennant, S.M.; Diallo, S.; Haim Levy, H.; Livio, S.; Sow, S.O.; Milagritos Tapia, M.; Fields, P.I.; Mikoleit, M.; Tamboura, B.; Kotloff, K.L.; et al. Identification by PCR of non-typhoidal Salmonella enterica serovars associated with invasive infections among febrile patients in Mali. PLoS Negl. Trop. Dis. 2010, 4, e621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barco, L.; Lettini, A.A.; Ramon, E.; Longo, A.; Saccardin, C.; Pozza, M.C.; Ricci, A. A rapid and sensitive method to identify and differentiate Salmonella enterica serotype Typhimurium and Salmonella enterica serotype 4,[5],12:i:- by combining traditional serotyping and multiplex polymerase chain reaction. Foodborne Pathog. Dis. 2011, 8, 741–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabsch, W.; Tschäpe, H.; Bäumler, A.J. Non-typhoidal salmonellosis: Emerging problems. Microbes Infect. 2001, 3, 237–247. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| O-serogroup | Serovar | N = 359 |
|---|---|---|
| O:2 | Paratyphi A | 23 |
| Kiel | 1 | |
| Koessen | 1 | |
| Nitra | 1 | |
| O:4 | Paratyphi B | 23 |
| Aberdeen | 1 | |
| Abony | 2 | |
| Agona | 6 | |
| Banana | 1 | |
| Brandenburg | 5 | |
| Bredeney | 1 | |
| Chester | 1 | |
| Coeln | 1 | |
| Derby | 8 | |
| Heidelberg | 2 | |
| Hessarek | 1 | |
| Indiana | 2 | |
| Kisangani | 1 | |
| Paratyphi B var. Java | 1 | |
| Saintpaul | 3 | |
| Schleissheim | 2 | |
| Schwarzengrund | 1 | |
| Stanleyville | 2 | |
| Typhimurium | 59 | |
| Typhimurium (monophasic variant) | 98 | |
| Subsp. salamae O:4 | 1 | |
| O:7 | Paratyphi C | 7 |
| Braenderup | 5 | |
| Choleraesuis | 2 | |
| Infantis | 11 | |
| Isangi | 2 | |
| Livingstone | 2 | |
| Mikawasima | 1 | |
| Montevideo | 3 | |
| Ohio | 2 | |
| Oranienburg | 3 | |
| Singapore | 1 | |
| Strathcona | 2 | |
| Thompson | 1 | |
| Virchow | 1 | |
| Subsp. houtanae O:7 | 2 | |
| O:9 | Typhi | 30 |
| Dublin | 1 | |
| Enteritidis | 19 | |
| Javiana | 1 | |
| Kapemba | 1 | |
| Napoli | 9 | |
| Panama | 2 | |
| Zaiman | 1 | |
| Subsp. salamae O:9,12 | 2 |
| O-groups | Training Set (N) | Testing Set CBA (N) | Testing Set TSA (N) | Testing Set CHO (N) | Testing Set MHA (N) |
|---|---|---|---|---|---|
| Paratyphi A (23) | 5 | 18 | 23 | 23 | 23 |
| Non-Paratyphi A (3) | 3 | 3 | 3 | 3 | 3 |
| Paratyphi B (23) | 5 | 18 | 23 | 23 | 23 |
| Non-Paratyphi B (199) | 27 | 199 | 199 | 199 | 199 |
| Paratyphi C (7) | 2 | 5 | 7 | 7 | 7 |
| Non-Paratyphi C (38) | 14 | 37 | 38 | 38 | 38 |
| Typhi (30) | 5 | 25 | 30 | 30 | - |
| Non-Typhi (36) | 14 | 36 | 36 | 36 | - |
| Agar Medium | Parameter | Classifier S. Paratyphi A | Classifier S. Paratyphi B | Classifier S. Paratyphi C | Classifier S. Typhi |
|---|---|---|---|---|---|
| CBA | Accuracy | 99.5% | 87.0% | 99% | 99.9% |
| Error rate | 0 | 10.8% | 0 | 0 | |
| Failed classification | 0.5% | 2.2% | 1% | 0.1% | |
| Sensitivity | 100% | 77.8% | 100% | 100% | |
| Specificity | 100% | 94.8% | 100% | 100% | |
| Positive predictive value # | 100% | 77.8% | 100% | 100% | |
| Negative predictive value # | 100% | 83.1% | 100% | 100% | |
| TSA | Accuracy | 97.0% | 87.8% | 95.5% | 99.6% |
| Error rate | 1% | 10.1% | 2.8% | 0 | |
| Failed classification | 2% | 2.1% | 1.7% | 0.4% | |
| CHO | Accuracy | 100% | 86.6% | 98.8% | 99.1% |
| Error rate | 0 | 12.0% | 1.2% | 0 | |
| Failed classification | 0 | 1.4% | 0 | 0.9% | |
| MHA | Accuracy | 94% | 84.9% | 94.7% | - |
| Error rate | 0 | 11.7% | 0 | - | |
| Failed classification | 6% | 3.4% | 5.3% | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordovana, M.; Mauder, N.; Kostrzewa, M.; Wille, A.; Rojak, S.; Hagen, R.M.; Ambretti, S.; Pongolini, S.; Soliani, L.; Justesen, U.S.; et al. Classification of Salmonella enterica of the (Para-)Typhoid Fever Group by Fourier-Transform Infrared (FTIR) Spectroscopy. Microorganisms 2021, 9, 853. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040853
Cordovana M, Mauder N, Kostrzewa M, Wille A, Rojak S, Hagen RM, Ambretti S, Pongolini S, Soliani L, Justesen US, et al. Classification of Salmonella enterica of the (Para-)Typhoid Fever Group by Fourier-Transform Infrared (FTIR) Spectroscopy. Microorganisms. 2021; 9(4):853. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040853
Chicago/Turabian StyleCordovana, Miriam, Norman Mauder, Markus Kostrzewa, Andreas Wille, Sandra Rojak, Ralf Matthias Hagen, Simone Ambretti, Stefano Pongolini, Laura Soliani, Ulrik S. Justesen, and et al. 2021. "Classification of Salmonella enterica of the (Para-)Typhoid Fever Group by Fourier-Transform Infrared (FTIR) Spectroscopy" Microorganisms 9, no. 4: 853. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040853