
Enteric Ganglioneuritis, a Common Feature in a Subcutaneous TBEV Murine Infection Model

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Mice
2.3. Virus and Cell Culture
2.4. Experimental Design
2.5. RNA Extraction
2.6. Real-Time Quantitative RT-PCR
2.7. Histology and Immunohistochemistry
2.8. Statistical Analysis
3. Results
3.1. Course of Infection
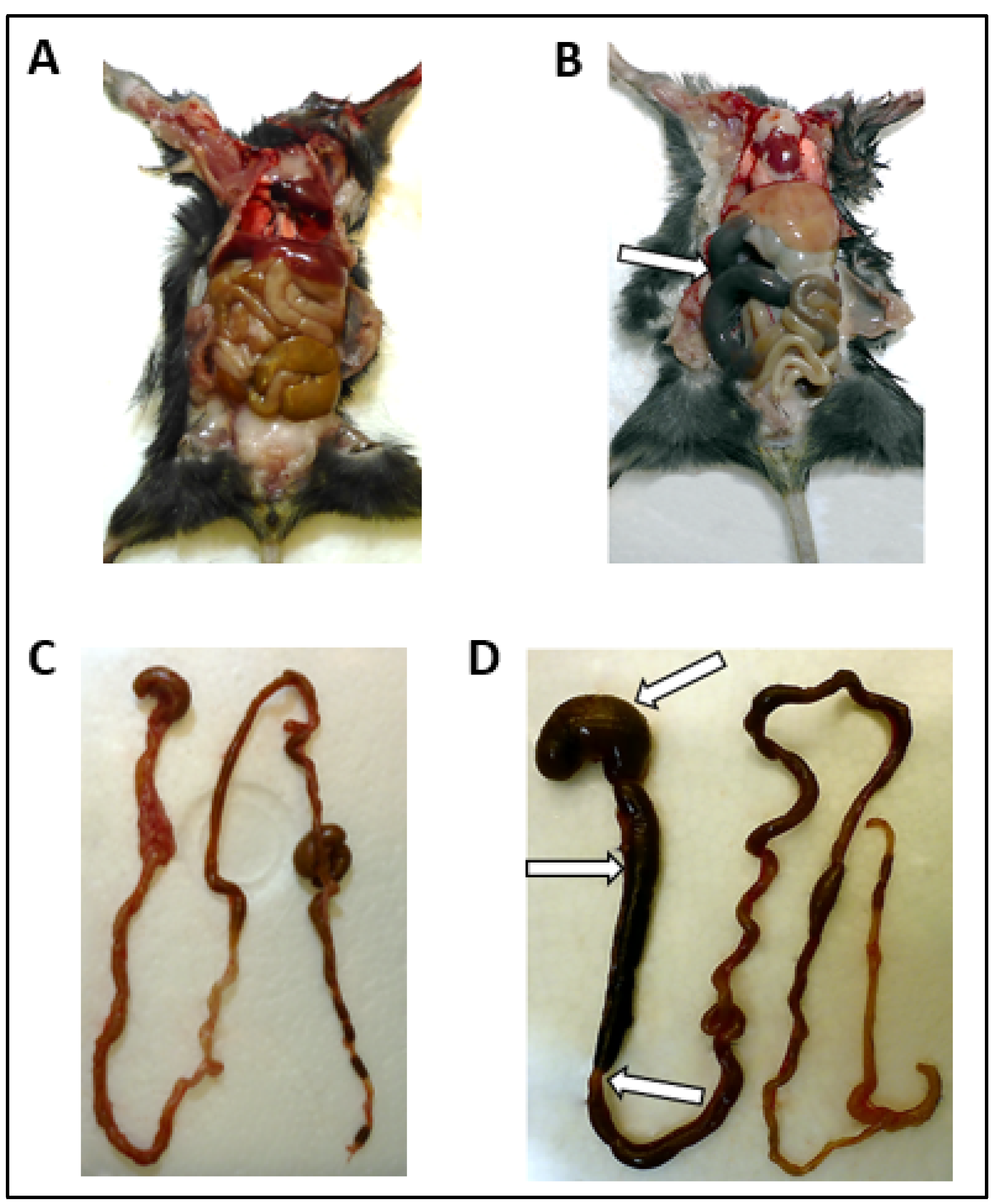
3.2. Macroscopic Alterations of the GI Tract
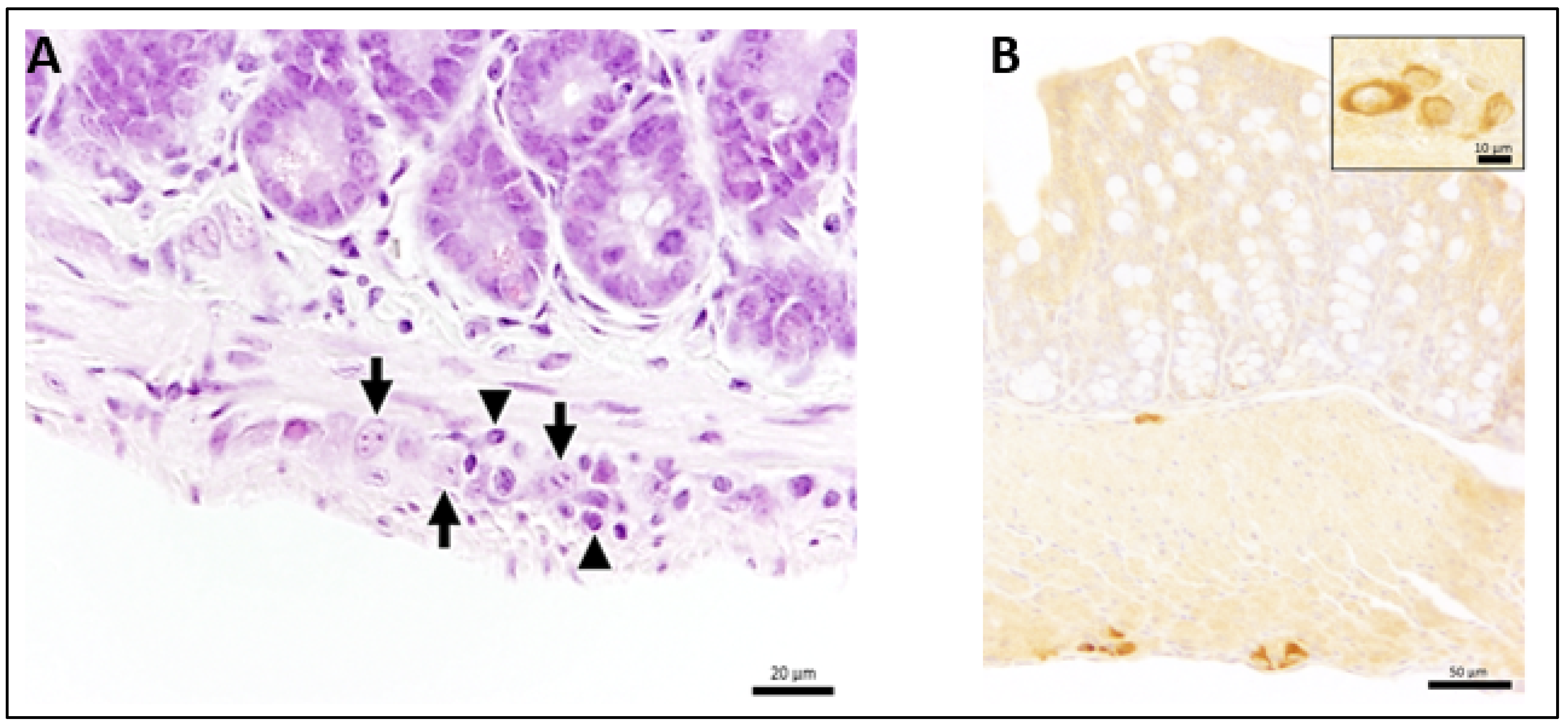
3.3. Ganglioneuritis in Intramural Ganglions of Small and Large Intestine and Viral Antigen in Enteric Plexus
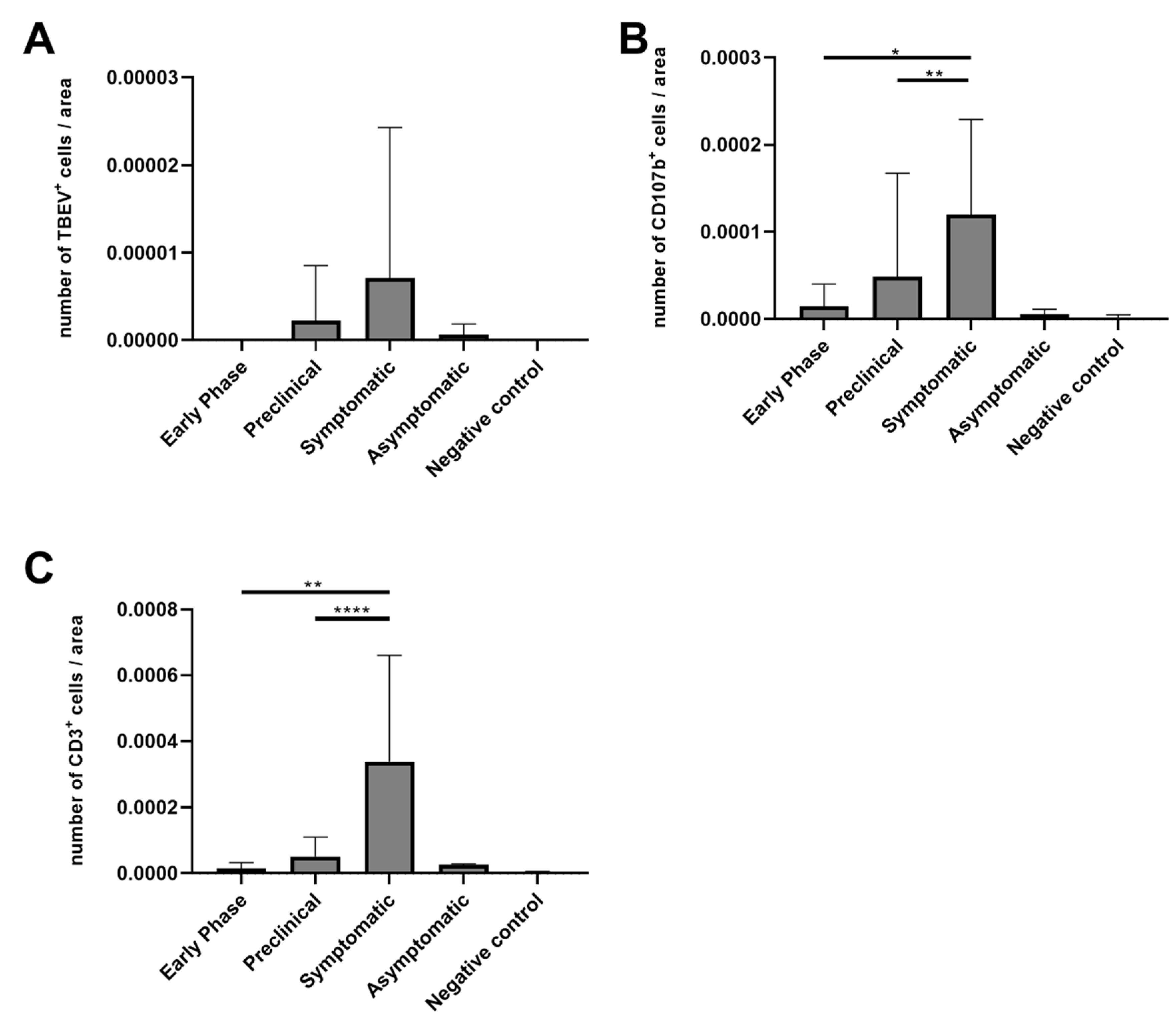
3.4. Numbers of CD3-Positive Cells and Macrophages Were Increased in Mice with Ganglioneuritis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gritsun, T.S.; Lashkevich, V.A.; Gould, E.A. Tick-borne encephalitis. Antivir. Res. 2003, 57, 129–146. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-borne encephalitis virus–A review of an emerging zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef]
- Ruzek, D.; Avsic Zupanc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy, and vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, L.; Vapalahti, O. Tick-borne encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Dai, X.; Shang, G.; Lu, S.; Yang, J.; Xu, J. A new subtype of eastern tick-borne encephalitis virus discovered in Qinghai-Tibet Plateau, China. Emerg. Microbes Infect. 2018, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Kovalev, S.Y.; Mukhacheva, T.A. Reconsidering the classification of tick-borne encephalitis virus within the Siberian subtype gives new insights into its evolutionary history. Infect. Genet. Evol. 2017, 55, 159–165. [Google Scholar] [CrossRef]
- Boelke, M.; Bestehorn, M.; Marchwald, B.; Kubinski, M.; Liebig, K.; Glanz, J.; Schulz, C.; Dobler, G.; Monazahian, M.; Becker, S.C. First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses 2019, 11, 462. [Google Scholar] [CrossRef] [Green Version]
- Holding, M.; Dowall, S.D.; Medlock, J.M.; Carter, D.P.; McGinley, L.; Curran-French, M.; Pullan, S.T.; Chamberlain, J.; Hansford, K.M.; Baylis, M.; et al. Detection of new endemic focus of tick-borne encephalitis virus (TBEV), Hampshire/Dorset border, England, September 2019. Eurosurveillance 2019, 24, 1900658. [Google Scholar] [CrossRef] [Green Version]
- Holding, M.; Dowall, S.D.; Medlock, J.M.; Carter, D.P.; Pullan, S.T.; Lewis, J.; Vipond, R.; Rocchi, M.S.; Baylis, M.; Hewson, R. Tick-Borne Encephalitis Virus, United Kingdom. Emerg. Infect. Dis. 2020, 26, 90–96. [Google Scholar] [CrossRef]
- Heinz, F.X.; Stiasny, K.; Holzmann, H.; Kundi, M.; Sixl, W.; Wenk, M.; Kainz, W.; Essl, A.; Kunz, C. Emergence of tick-borne encephalitis in new endemic areas in Austria: 42 years of surveillance. Eurosurveillance 2015, 20, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Fares, W.; Dachraoui, K.; Cherni, S.; Barhoumi, W.; Slimane, T.B.; Younsi, H.; Zhioua, E. Tick-borne encephalitis virus in Ixodes ricinus (Acari: Ixodidae) ticks, Tunisia. Ticks Tick Borne Dis. 2021, 12, 101606. [Google Scholar] [CrossRef]
- Labuda, M.; Austyn, J.M.; Zuffova, E.; Kozuch, O.; Fuchsberger, N.; Lysy, J.; Nuttall, P.A. Importance of localized skin infection in tick-borne encephalitis virus transmission. Virology 1996, 219, 357–366. [Google Scholar] [CrossRef]
- Kaiser, R. The clinical and epidemiological profile of tick-borne encephalitis in southern Germany 1994-98: A prospective study of 656 patients. Brain 1999, 122 Pt 11, 2067–2078. [Google Scholar] [CrossRef] [Green Version]
- Chrdle, A.; Chmelik, V.; Ruzek, D. Tick-borne encephalitis: What travelers should know when visiting an endemic country. Hum. Vaccines Immunother. 2016, 12, 2694–2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiter, I.; Steinbrecher, A.; Flugel, D.; Bogdahn, U.; Schulte-Mattler, W. Autonomic involvement in tick-borne encephalitis (TBE): Report of five cases. Eur. J. Med. Res. 2006, 11, 261–265. [Google Scholar] [PubMed]
- Versace, V.; Gutmann, B.; Sebastianelli, L.; Martignago, S.; Saltuari, L.; Nardone, R.; Wiedermann, C.J. Tick-borne viral encephalomyeloradiculitis complicated by severe autonomic myenteric involvement resulting in irreversible adynamic ileus. Clin. Auton. Res. 2017, 27, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Dobler, G.; Bestehorn, M.; Antwerpen, M.; Overby-Wernstedt, A. Complete Genome Sequence of a Low-Virulence Tick-Borne Encephalitis Virus Strain. Genome Announc. 2016, 4, e01145-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autschbach, R.; Zaremba, A.; Ulrich, B. Pseudo-obstruction of the colon following tick bite injury. Chirurg 1989, 60, 365–367. [Google Scholar]
- Nagata, N.; Iwata-Yoshikawa, N.; Hayasaka, D.; Sato, Y.; Kojima, A.; Kariwa, H.; Takashima, I.; Takasaki, T.; Kurane, I.; Sata, T.; et al. The pathogenesis of 3 neurotropic flaviviruses in a mouse model depends on the route of neuroinvasion after viremia. J. Neuropathol. Exp. Neurol. 2015, 74, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Sasaki, M.; Okumura, M.; Kim, E.; Sawa, H. Flavivirus encephalitis: Pathological aspects of mouse and other animal models. Vet. Pathol. 2010, 47, 806–818. [Google Scholar] [CrossRef]
- White, J.P.; Xiong, S.; Malvin, N.P.; Khoury-Hanold, W.; Heuckeroth, R.O.; Stappenbeck, T.S.; Diamond, M.S. Intestinal Dysmotility Syndromes following Systemic Infection by Flaviviruses. Cell 2018, 175, 1198–1212. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H.A. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Shimizu, S. Routes of administration. In The Laboratory Mouse; Elsevier: Amsterdam, The Netherlands, 2004; pp. 527–541. [Google Scholar]
- National Centre for Replacement, Refinement& Reduction of Animals in Research, Humane Endpoints. Available online: https://www.nc3rs.org.uk/humane-endpoints (accessed on 8 April 2021).
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick borne encephalitis virus (TBEV) RNA. J. Clin. Virol. 2003, 27, 136–145. [Google Scholar] [CrossRef]
- Wilhelmi, V.; Simon, C.O.; Podlech, J.; Bohm, V.; Daubner, T.; Emde, S.; Strand, D.; Renzaho, A.; Lemmermann, N.A.; Seckert, C.K.; et al. Transactivation of cellular genes involved in nucleotide metabolism by the regulatory IE1 protein of murine cytomegalovirus is not critical for viral replicative fitness in quiescent cells and host tissues. J. Virol. 2008, 82, 9900–9916. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Leitzen, E.; Goebbels, S.; Nave, K.A.; Baumgartner, W.; Hansmann, F. Comparison of Theiler’s Murine Encephalomyelitis Virus Induced Spinal Cord and Peripheral Nerve Lesions Following Intracerebral and Intraspinal Infection. Int. J. Mol. Sci. 2019, 20, 5134. [Google Scholar] [CrossRef] [Green Version]
- Leitzen, E.; Raddatz, B.B.; Jin, W.; Goebbels, S.; Nave, K.A.; Baumgartner, W.; Hansmann, F. Virus-triggered spinal cord demyelination is followed by a peripheral neuropathy resembling features of Guillain-Barre Syndrome. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Zivcec, M.; Safronetz, D.; Scott, D.; Robertson, S.; Ebihara, H.; Feldmann, H. Lethal Crimean-Congo hemorrhagic fever virus infection in interferon α/β receptor knockout mice is associated with high viral loads, proinflammatory responses, and coagulopathy. J. Infect. Dis. 2013, 207, 1909–1921. [Google Scholar] [CrossRef]
- Lieskovská, J.; Páleníková, J.; Langhansová, H.; Chmelař, J.; Kopecký, J. Saliva of Ixodes ricinus enhances TBE virus replication in dendritic cells by modulation of pro-survival Akt pathway. Virology 2018, 514, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Egyed, L.; Zöldi, V.; Szeredi, L. Subclinical Tick-Borne Encephalitis Virus in Experimentally Infected Apodemus agrarius. Intervirology 2015, 58, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Klem, F.; Wadhwa, A.; Prokop, L.J.; Sundt, W.J.; Farrugia, G.; Camilleri, M.; Singh, S.; Grover, M. Prevalence, Risk Factors, and Outcomes of Irritable Bowel Syndrome After Infectious Enteritis: A Systematic Review and Meta-analysis. Gastroenterology 2017, 152, 1042–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furness, J.B. The Enteric Nervous System; Blackwell Pub.: Malden, MA, USA, 2006; p. xiii. 274p. [Google Scholar]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Wright, C.M.; Heuckeroth, R.O. Unexpected Roles for the Second Brain: Enteric Nervous System as Master Regulator of Bowel Function. Annu. Rev. Physiol. 2019, 81, 235–259. [Google Scholar] [CrossRef] [PubMed]
- Gelpi, E.; Preusser, M.; Laggner, U.; Garzuly, F.; Holzmann, H.; Heinz, F.X.; Budka, H. Inflammatory response in human tick-borne encephalitis: Analysis of postmortem brain tissue. J. Neurovirol. 2006, 12, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, D.; Nagata, N.; Fujii, Y.; Hasegawa, H.; Sata, T.; Suzuki, R.; Gould, E.A.; Takashima, I.; Koike, S. Mortality following peripheral infection with Tick-borne encephalitis virus results from a combination of central nervous system pathology, systemic inflammatory and stress responses. Virology 2009, 390, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Ruzek, D.; Salat, J.; Palus, M.; Gritsun, T.S.; Gould, E.A.; Dykova, I.; Skallova, A.; Jelinek, J.; Kopecky, J.; Grubhoffer, L. CD8+ T-cells mediate immunopathology in tick-borne encephalitis. Virology 2009, 384, 1–6. [Google Scholar] [CrossRef]
- Gelpi, E.; Preusser, M.; Garzuly, F.; Holzmann, H.; Heinz, F.X.; Budka, H. Visualization of Central European tick-borne encephalitis infection in fatal human cases. J. Neuropathol. Exp. Neurol. 2005, 64, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Maximova, O.A.; Faucette, L.J.; Ward, J.M.; Murphy, B.R.; Pletnev, A.G. Cellular inflammatory response to flaviviruses in the central nervous system of a primate host. J. Histochem. Cytochem. 2009, 57, 973–989. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| 1st Antibody | Pre-Treatment | 2nd Antibody | |||
|---|---|---|---|---|---|
| Antigen, Target | Product Name | Clonality, Host Species, Clone | Dilution | ||
| CD3, T lymphocytes | Dako A0452 | Polyclonal, rabbit | 1:200 | Citrate buffer, microwave | Goat-anti-rabbit biotinylated |
| CD107b, microglia/macrophages | BioRad MCA2293B | Monoclonal, rat, clone M3/84 | 1:200 | Citrate buffer, microwave | Rabbit-anti-rat biotinylated |
| CD45R, B lymphocytes | BD Bioscience 553085 | Monoclonal rat, clone B220- biotinylated | 1:1000 | Citrate buffer, microwave | - |
| Anti-TBEV-rabbit serum | Center for Virology, Medical University of Vienna, Austria | Polyclonal, rabbit | 1:800 | Proteinase K | Goat-anti-rabbit biotinylated |
| Animal ID | dpi | Clinical Score | Ganglioneuritis | Viral Antigen | Macroscopic Alterations | qRT-PCR Intestine | |||
|---|---|---|---|---|---|---|---|---|---|
| Stomach | Proximal | Mid | Distal | ||||||
| M8 | 2 | 0 | + | − | − | − | − | − | − |
| M11 | 4 | 0 | + | − | − | − | − | − | (+) |
| M15 | 7 | 0 | + | + | − | − | − | − | + |
| M28 | 7 | 0 | (+) | − | − | − | − | − | + |
| M24 | 8 | HEP | n.d. | n.d. | − | + | − | − | + |
| M25 | 8 | HEP | +++ | + | − | + | + | − | + |
| M26 | 8 | HEP | ++ | + | + | + | (+) | − | + |
| M18 | 9 | HEP | + | + | (+) | + | − | − | + |
| M19 | 9 | HEP | ++ | − | − | + | + | − | + |
| M17 | 10 | 6 | ++ | + | − | − | + | + | + |
| M22 | 10 | HEP | ++ | + | + | + | (+) | − | + |
| M27 | 10 | 9 | ++ | + | − | (+) | + | − | + |
| M21 | 11 | HEP | +++ | − | − | − | − | − | + |
| M20 | 12 | HEP | + | − | − | + | + | − | + |
| M23 | 12 | HEP | + | − | + | + | (+) | − | + |
| M16 | 14 | 0 | + | + | − | − | − | − | − |
| M29 | 6 | HEP | + | − | − | − | − | − | + |
| M30 | 6 | HEP | + | + | − | − | − | − | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boelke, M.; Puff, C.; Becker, K.; Hellhammer, F.; Gusmag, F.; Marks, H.; Liebig, K.; Stiasny, K.; Dobler, G.; Baumgärtner, W.; et al. Enteric Ganglioneuritis, a Common Feature in a Subcutaneous TBEV Murine Infection Model. Microorganisms 2021, 9, 875. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040875
Boelke M, Puff C, Becker K, Hellhammer F, Gusmag F, Marks H, Liebig K, Stiasny K, Dobler G, Baumgärtner W, et al. Enteric Ganglioneuritis, a Common Feature in a Subcutaneous TBEV Murine Infection Model. Microorganisms. 2021; 9(4):875. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040875
Chicago/Turabian StyleBoelke, Mathias, Christina Puff, Kathrin Becker, Fanny Hellhammer, Frederic Gusmag, Hannah Marks, Katrin Liebig, Karin Stiasny, Gerhard Dobler, Wolfgang Baumgärtner, and et al. 2021. "Enteric Ganglioneuritis, a Common Feature in a Subcutaneous TBEV Murine Infection Model" Microorganisms 9, no. 4: 875. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040875