
Activation of the JNK/MAPK Signaling Pathway by TGF-β1 Enhances Neonatal Fc Receptor Expression and IgG Transcytosis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Antibodies
2.2. Western Blotting
2.3. MAPKs Inhibition Assays
2.4. Construction of Reporter Plasmid and Luciferase Assays
2.5. Chromatin Immunoprecipitation
2.6. Statistical Analyses
3. Results
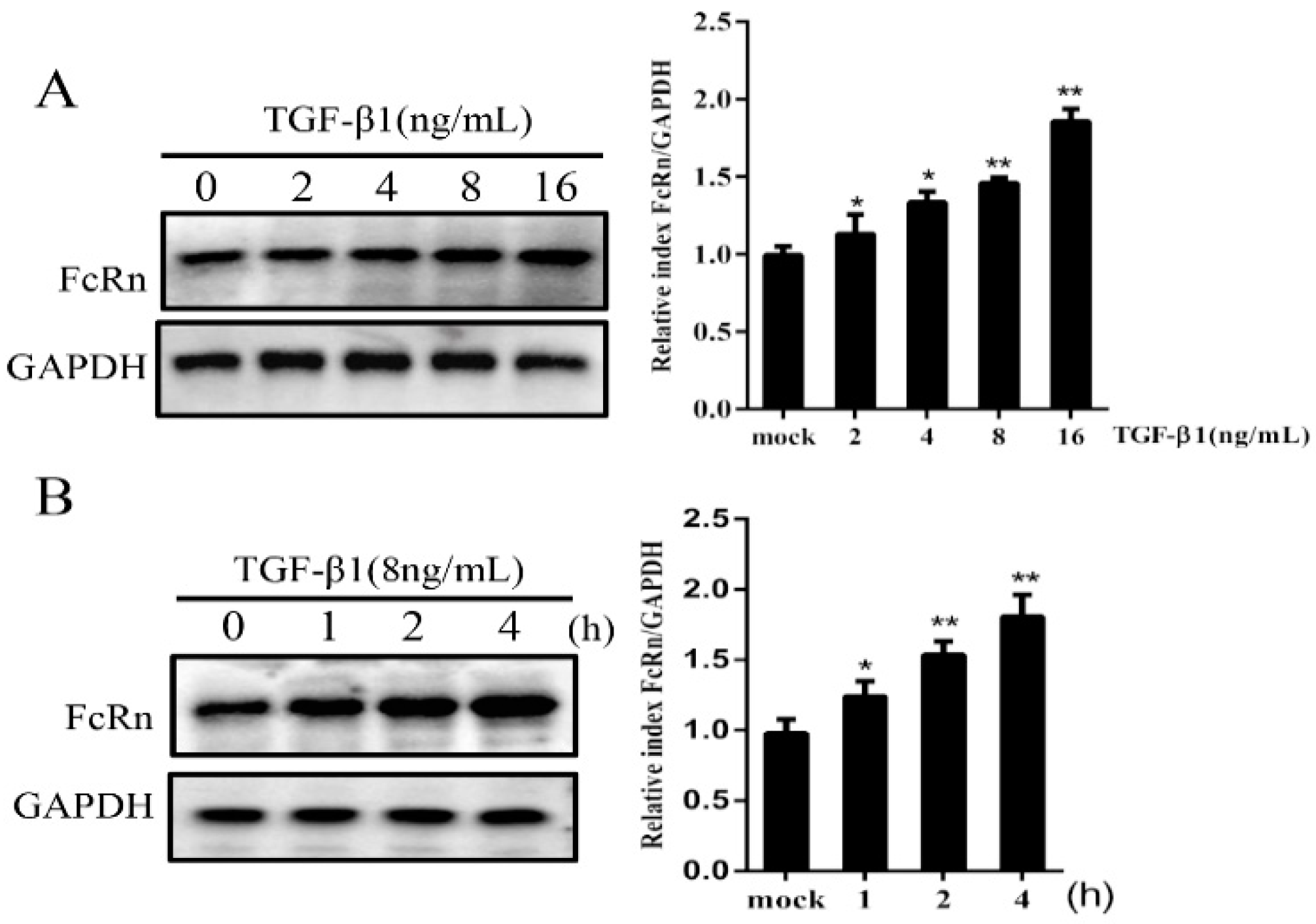
3.1. TGF-β1 Upregulated pFcRn Expression in IPEC-J2 Cells
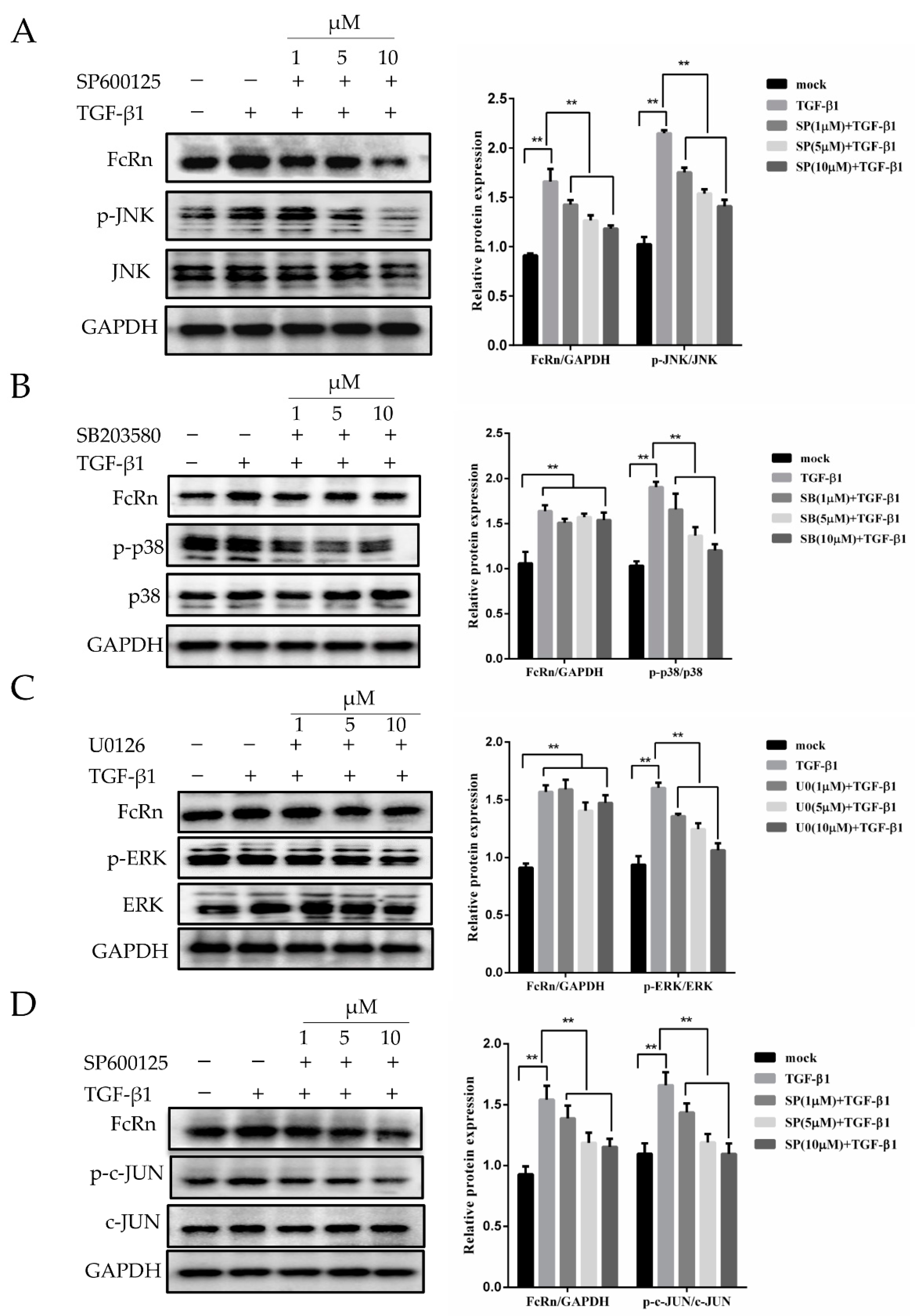
3.2. Effects of MAPK Inhibition on pFcRn Expression
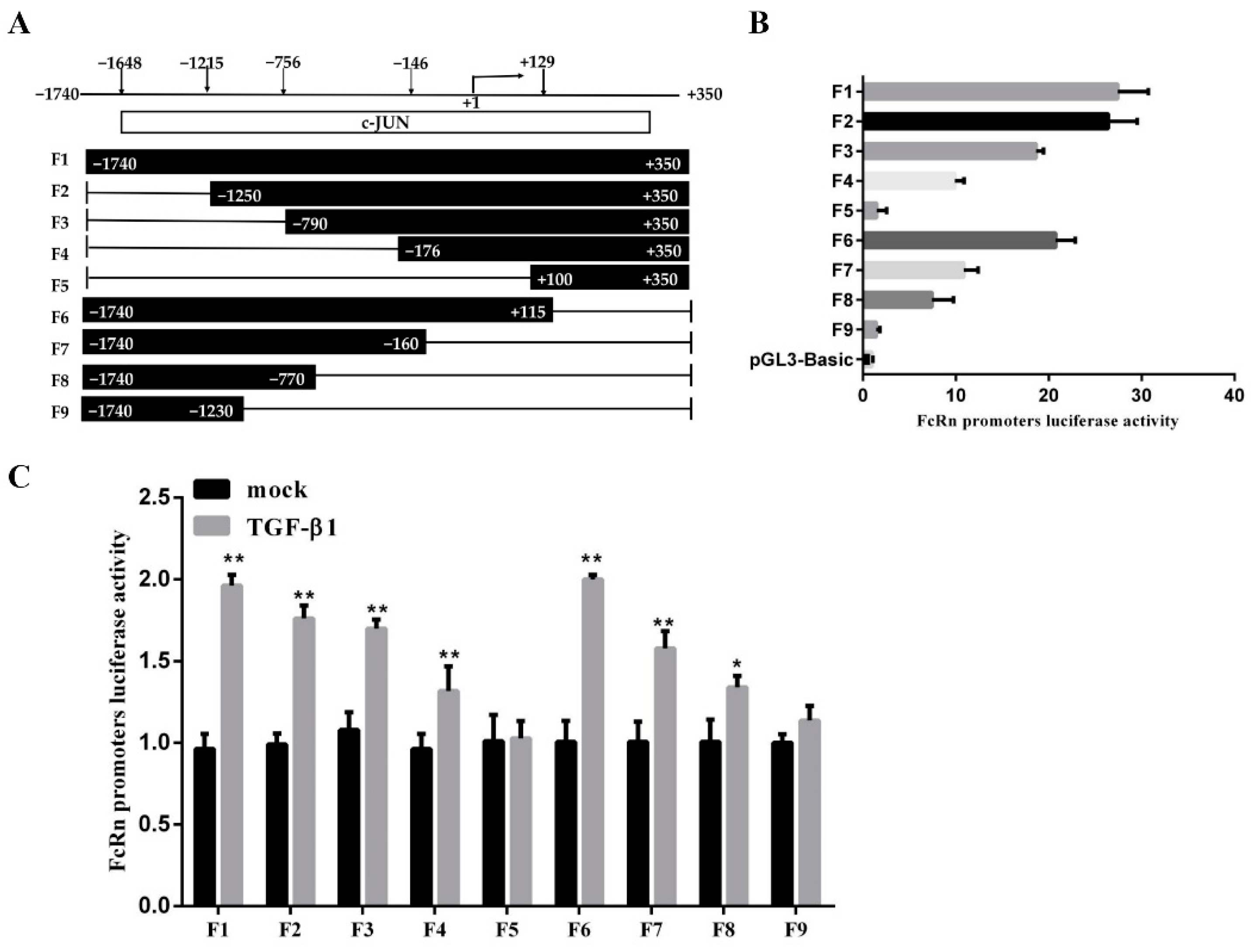
3.3. Screening for c-JUN Binding Sites Adjacent to the pFcRn Promoter
3.4. The pFcRn Promoter Contained Three c-JUN Binding Sites as Confirmed by ChIP
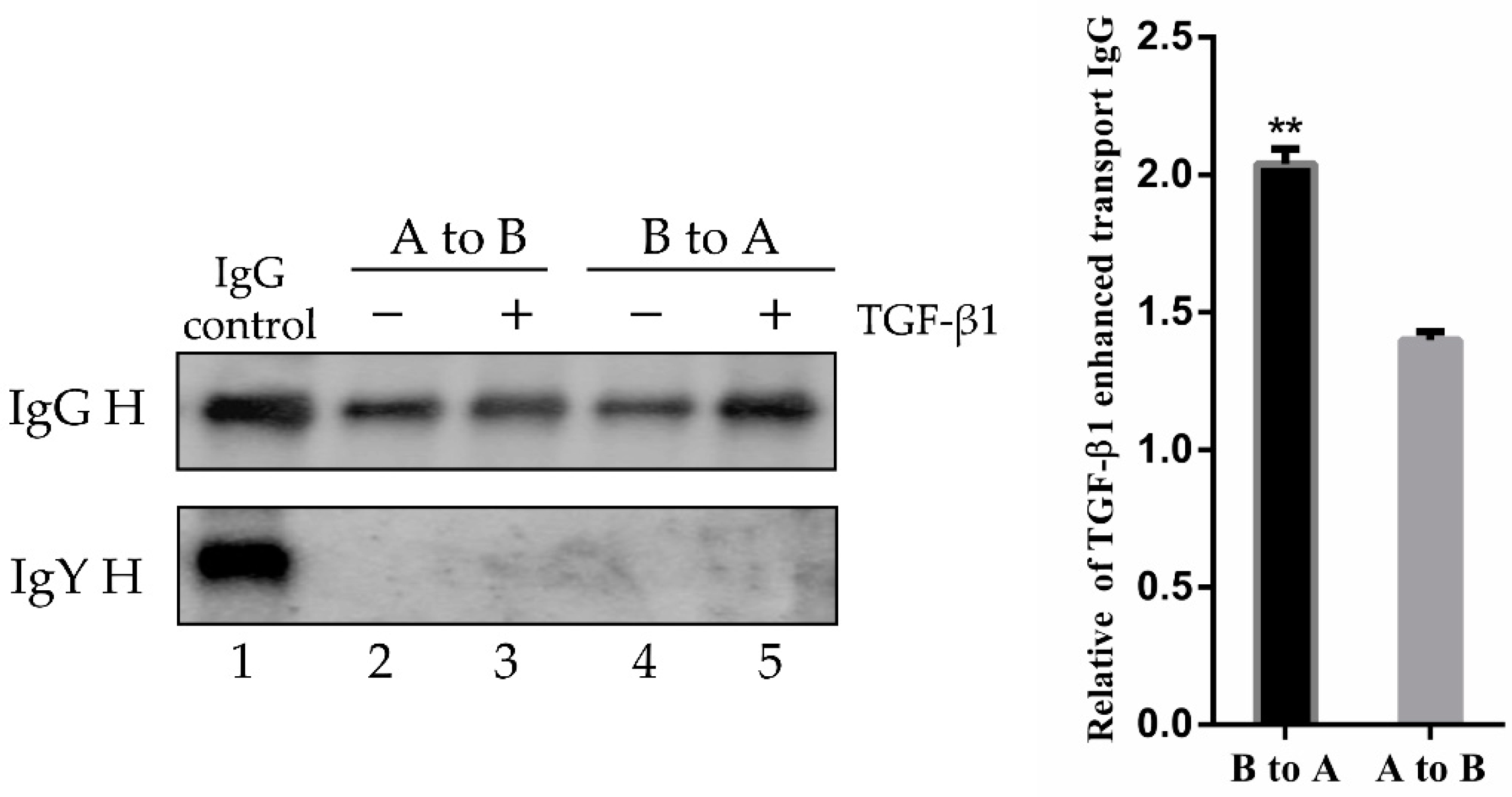
3.5. TGF-β1 Induced pFcRn-Mediated IgG Transcytosis in Polarized IPEC-J2 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ye, L.; Liu, X.; Rout, S.N.; Li, Z.; Yan, Y.; Lu, L.; Kamala, T.; Nanda, N.K.; Song, W.; Samal, S.K.; et al. The mhc class ii-associated invariant chain interacts with the neonatal fc gamma receptor and modulates its trafficking to endosomal/lysosomal compartments. J. Immunol. (Baltim. Md. 1950) 2008, 181, 2572–2585. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Palaniyandi, S.; Zeng, R.; Tuo, W.; Roopenian, D.C.; Zhu, X. Transfer of igg in the female genital tract by mhc class i-related neonatal fc receptor (fcrn) confers protective immunity to vaginal infection. Proc. Natl. Acad. Sci. USA 2011, 108, 4388–4393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyzik, M.; Rath, T.; Kuo, T.T.; Win, S.; Baker, K.; Hubbard, J.J.; Grenha, R.; Gandhi, A.; Krämer, T.D.; Mezo, A.R.; et al. Hepatic fcrn regulates albumin homeostasis and susceptibility to liver injury. Proc. Natl. Acad. Sci. USA 2017, 114, 2862–2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhury, C.; Mehnaz, S.; Robinson, J.M.; Hayton, W.L.; Pearl, D.K.; Roopenian, D.C.; Anderson, C.L. The major histocompatibility complex-related fc receptor for igg (fcrn) binds albumin and prolongs its lifespan. J. Exp. Med. 2003, 197, 315–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghetie, V.; Ward, E.S. Multiple roles for the major histocompatibility complex class i- related receptor fcrn. Annu. Rev. Immunol. 2000, 18, 739–766. [Google Scholar] [CrossRef]
- Baker, K.; Qiao, S.W.; Kuo, T.T.; Aveson, V.G.; Platzer, B.; Andersen, J.T.; Sandlie, I.; Chen, Z.; de Haar, C.; Lencer, W.I.; et al. Neonatal fc receptor for igg (fcrn) regulates cross-presentation of igg immune complexes by cd8-cd11b+ dendritic cells. Proc. Natl. Acad. Sci. USA 2011, 108, 9927–9932. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Palaniyandi, S.; Zeng, R.; Bai, Y.; Liu, X.; Wang, Y.; Pauza, C.D.; Roopenian, D.C.; Zhu, X. A neonatal fc receptor-targeted mucosal vaccine strategy effectively induces hiv-1 antigen-specific immunity to genital infection. J. Virol. 2011, 85, 10542–10553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, L.; Zeng, R.; Bai, Y.; Roopenian, D.C.; Zhu, X. Efficient mucosal vaccination mediated by the neonatal fc receptor. Nat. Biotechnol. 2011, 29, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Rath, T.; Kuo, T.T.; Baker, K.; Qiao, S.W.; Kobayashi, K.; Yoshida, M.; Roopenian, D.; Fiebiger, E.; Lencer, W.I.; Blumberg, R.S. The immunologic functions of the neonatal fc receptor for igg. J. Clin. Immunol. 2013, 33 (Suppl. 1), 9–17. [Google Scholar] [CrossRef]
- Qiao, S.W.; Kobayashi, K.; Johansen, F.E.; Sollid, L.M.; Andersen, J.T.; Milford, E.; Roopenian, D.C.; Lencer, W.I.; Blumberg, R.S. Dependence of antibody-mediated presentation of antigen on fcrn. Proc. Natl. Acad. Sci. USA 2008, 105, 9337–9342. [Google Scholar] [CrossRef] [Green Version]
- Vidarsson, G.; Stemerding, A.M.; Stapleton, N.M.; Spliethoff, S.E.; Janssen, H.; Rebers, F.E.; de Haas, M.; van de Winkel, J.G. Fcrn: An igg receptor on phagocytes with a novel role in phagocytosis. Blood 2006, 108, 3573–3579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Claypool, S.M.; Wagner, J.S.; Mizoguchi, E.; Mizoguchi, A.; Roopenian, D.C.; Lencer, W.I.; Blumberg, R.S. Human neonatal fc receptor mediates transport of igg into luminal secretions for delivery of antigens to mucosal dendritic cells. Immunity 2004, 20, 769–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stapleton, N.M.; Einarsdóttir, H.K.; Stemerding, A.M.; Vidarsson, G. The multiple facets of fcrn in immunity. Immunol. Rev. 2015, 268, 253–268. [Google Scholar] [CrossRef]
- Hase, K.; Ohno, H. Epithelial cells as sentinels in mucosal immune barrier. Nihon Rinsho Men’eki Gakkai kaishi Jpn. J. Clin. Immunol. 2006, 29, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workman, L.M.; Habelhah, H. Tnfr1 signaling kinetics: Spatiotemporal control of three phases of ikk activation by posttranslational modification. Cell. Signal. 2013, 25, 1654–1664. [Google Scholar] [CrossRef] [Green Version]
- Pal, K.; Kaetzel, C.S.; Brundage, K.; Cunningham, C.A.; Cuff, C.F. Regulation of polymeric immunoglobulin receptor expression by reovirus. J. Gen. Virol. 2005, 86, 2347–2357. [Google Scholar] [CrossRef]
- Johansen, F.E.; Kaetzel, C.S. Regulation of the polymeric immunoglobulin receptor and iga transport: New advances in environmental factors that stimulate pigr expression and its role in mucosal immunity. Mucosal Immunol. 2011, 4, 598–602. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ye, L.; Christianson, G.J.; Yang, J.Q.; Roopenian, D.C.; Zhu, X. Nf-kappab signaling regulates functional expression of the mhc class i-related neonatal fc receptor for igg via intronic binding sequences. J. Immunol. (Baltim. Md. 1950) 2007, 179, 2999–3011. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ye, L.; Bai, Y.; Mojidi, H.; Simister, N.E.; Zhu, X. Activation of the jak/stat-1 signaling pathway by ifn-gamma can down-regulate functional expression of the mhc class i-related neonatal fc receptor for igg. J. Immunol. (Baltim. Md. 1950) 2008, 181, 449–463. [Google Scholar] [CrossRef] [Green Version]
- Gohy, S.T.; Detry, B.R.; Lecocq, M.; Bouzin, C.; Weynand, B.A.; Amatngalim, G.D.; Sibille, Y.M.; Pilette, C. Polymeric immunoglobulin receptor down-regulation in chronic obstructive pulmonary disease. Persistence in the cultured epithelium and role of transforming growth factor-β. Am. J. Respir. Crit. Care Med. 2014, 190, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, C.; Guisset, A.; Detry, B.; Sibille, Y.; Pilette, C. Dual effect of neutrophils on pigr/secretory component in human bronchial epithelial cells: Role of tgf-beta. J. Biomed. Biotechnol. 2010, 2010, 428618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dignass, A.U. Mechanisms and modulation of intestinal epithelial repair. Inflamm. Bowel Dis. 2001, 7, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Barnard, J.A.; Beauchamp, R.D.; Coffey, R.J.; Moses, H.L. Regulation of intestinal epithelial cell growth by transforming growth factor type beta. Proc. Natl. Acad. Sci. USA 1989, 86, 1578–1582. [Google Scholar] [CrossRef] [Green Version]
- Endsley, M.A.; Njongmeta, L.M.; Shell, E.; Ryan, M.W.; Indrikovs, A.J.; Ulualp, S.; Goldblum, R.M.; Mwangi, W.; Estes, D.M. Human iga-inducing protein from dendritic cells induces iga production by naive igd+ b cells. J. Immunol. (Baltim. Md. 1950) 2009, 182, 1854–1859. [Google Scholar] [CrossRef]
- Huang, C.; Jacobson, K.; Schaller, M.D. Map kinases and cell migration. J. Cell Sci. 2004, 117, 4619–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Z.; An, K.; Xie, L.; Wu, W.; Zhang, R.; Wang, D.; Fang, Y.; Chen, H.; Xiao, S.; Fang, L. Transmissible gastroenteritis virus infection induces nf-κb activation through rlr-mediated signaling. Virology 2017, 507, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Dai, L.; Yu, Q.; Yang, Q. Persistent transmissible gastroenteritis virus infection enhances enterotoxigenic escherichia coli k88 adhesion by promoting epithelial-mesenchymal transition in intestinal epithelial cells. J. Virol. 2017, 91, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, S.; Gao, Z.; Cao, R.; Yang, K.; Cui, Y.; Li, S.; Meng, X.; He, Q.; Li, Z. Transmissible gastroenteritis virus infection up-regulates fcrn expression via nucleocapsid protein and secretion of tgf-β in porcine intestinal epithelial cells. Front. Microbiol. 2019, 10, 3085. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Li, F.; He, Q.; Jin, H.; Liu, M.; Li, S.; Hu, S.; Xiao, Y.; Bi, D.; Li, Z. Neonatal fc receptor-mediated igg transport across porcine intestinal epithelial cells: Potentially provide the mucosal protection. DNA Cell Biol. 2016, 35, 301–309. [Google Scholar] [CrossRef]
- Guo, J.; Li, F.; Qian, S.; Bi, D.; He, Q.; Jin, H.; Luo, R.; Li, S.; Meng, X.; Li, Z. Tgev infection up-regulates fcrn expression via activation of nf-κb signaling. Sci. Rep. 2016, 6, 32154. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. CMLS 2013, 70, 631–659. [Google Scholar] [CrossRef]
- Pilette, C.; Ouadrhiri, Y.; Dimanche, F.; Vaerman, J.P.; Sibille, Y. Secretory component is cleaved by neutrophil serine proteinases but its epithelial production is increased by neutrophils through nf-kappa b- and p38 mitogen-activated protein kinase-dependent mechanisms. Am. J. Respir. Cell Mol. Biol. 2003, 28, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Pfundt, R.; Wingens, M.; Bergers, M.; Zweers, M.; Frenken, M.; Schalkwijk, J. Tnf-alpha and serum induce skalp/elafin gene expression in human keratinocytes by a p38 map kinase-dependent pathway. Arch. Dermatol. Res. 2000, 292, 180–187. [Google Scholar] [CrossRef]
- Hsieh, H.L.; Wang, H.H.; Wu, W.B.; Chu, P.J.; Yang, C.M. Transforming growth factor-β1 induces matrix metalloproteinase-9 and cell migration in astrocytes: Roles of ros-dependent erk- and jnk-nf-κb pathways. J. Neuroinflamm. 2010, 7, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervenak, J.; Doleschall, M.; Bender, B.; Mayer, B.; Schneider, Z.; Doleschall, Z.; Zhao, Y.; Bősze, Z.; Hammarström, L.; Oster, W.; et al. Nfκb induces overexpression of bovine fcrn: A novel mechanism that further contributes to the enhanced immune response in genetically modified animals carrying extra copies of fcrn. mAbs 2013, 5, 860–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikulska, J.E. Analysis of response elements involved in the regulation of the human neonatal fc receptor gene (fcgrt). PLoS ONE 2015, 10, e0139744. [Google Scholar]
- Newsted, D.; Fallahi, F.; Golshani, A.; Azizi, A. Advances and challenges in mucosal adjuvant technology. Vaccine 2015, 33, 2399–2405. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) |
|---|---|
| pFcRn-luc1-F | GCCGAGCTCAGTGTCCACAATCACATGAGCCA |
| pFcRn-luc1-R | CCCAAGCTTTCCTCCTCCTCCTCCTCCTCC |
| pFcRn-luc2-F | GCGAGCTCGACTGAGGTTCTTATCAGGGATGC |
| pFcRn-luc2-R | CCCAAGCTTTCCTCCTCCTCCTCCTCCTCC |
| pFcRn-luc3-F | GCGAGCTCCGACCTAGGCGAGGCCAA |
| pFcRn-luc3-R | CCCAAGCTTTCCTCCTCCTCCTCCTCCTCC |
| pFcRn-luc4-F | GCGAGCTCGCCGATCTCTAAAGGTGGGG |
| pFcRn-luc4-R | CCCAAGCTTTCCTCCTCCTCCTCCTCCTCC |
| pFcRn-luc5-F | GCGAGCTCAGGGATCGCGGCTGCTGT |
| pFcRn-luc5-R | CCCAAGCTTTCCTCCTCCTCCTCCTCCTCC |
| pFcRn-luc6-F | GCCGAGCTCAGTGTCCACAATCACATGAGCCA |
| pFcRn-luc6-R | CCCAAGCTTCTCACAGCAGCCGCGATC |
| pFcRn-luc7-F | GCCGAGCTCAGTGTCCACAATCACATGAGCCA |
| pFcRn-luc7-R | CCCAAGCTTCACCTTTAGAGATCGGCGCA |
| pFcRn-luc8-F | GCCGAGCTCAGTGTCCACAATCACATGAGCCA |
| pFcRn-luc8-R | CCCAAGCTTTTTTTGGCCTCGCCTAGGTC |
| pFcRn-luc9-F | GCCGAGCTCAGTGTCCACAATCACATGAGCCA |
| pFcRn-luc9-R | CCCAAGCTTTCCCTGATAAGAACCTCAGTCGG |
| Primer | Sequence (5′ to 3′) |
|---|---|
| Luc-146-F | TGACGAGGTAAGAAGGGGGC |
| Luc-146-R | GAGGGTGCCGGCGATCCA |
| Luc-790-F | TGCTGCGGCTCTGATTACACC |
| Luc-790-R | TGTGTCAAAACTTCATTTCTTTTTTG |
| Luc-1215-F | GTGTGTTAAGAACCCGACTGAGG |
| Luc-1215-R | GCTGAAGCTGTAGATATCAGCCTATAC |
| Luc-1684-F | TCTTTCTATATATATGCATACATCCTGTTG |
| Luc-1684-R | TTCTTTTGGATATATAGCTGGGAGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, S.; Li, C.; Liu, X.; Jia, X.; Xiao, Y.; Li, Z. Activation of the JNK/MAPK Signaling Pathway by TGF-β1 Enhances Neonatal Fc Receptor Expression and IgG Transcytosis. Microorganisms 2021, 9, 879. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040879
Qian S, Li C, Liu X, Jia X, Xiao Y, Li Z. Activation of the JNK/MAPK Signaling Pathway by TGF-β1 Enhances Neonatal Fc Receptor Expression and IgG Transcytosis. Microorganisms. 2021; 9(4):879. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040879
Chicago/Turabian StyleQian, Shaoju, Chenxi Li, Xi Liu, Xiangchao Jia, Yuncai Xiao, and Zili Li. 2021. "Activation of the JNK/MAPK Signaling Pathway by TGF-β1 Enhances Neonatal Fc Receptor Expression and IgG Transcytosis" Microorganisms 9, no. 4: 879. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040879