
The Divergent Key Residues of Two Agrobacterium fabrum (tumefaciens) CheY Paralogs Play a Key Role in Distinguishing Their Functions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Primers, and Bacterial Growth Conditions
2.2. DNA Manipulation, Construction of Gene-Deleted Mutants and Complemented Strains
2.3. Chemotaxis Assay
2.4. Observation and Analysis of Swimming Behavior
2.5. Construction of Plasmids for Expressing the Split-eGFP Fusion Proteins
2.6. Microscopic Observation of the Reconstituted Split-eGFP Fusions
2.7. Protein Overexpression and GST Pull-Down Assay
2.8. Electrophoresis and Immunoblotting Analysis
2.9. Site-Directed Mutation of CheY-Encoding Gene and the Test of the Function of Different CheY Variants
2.10. Statistical Analysis
3. Results
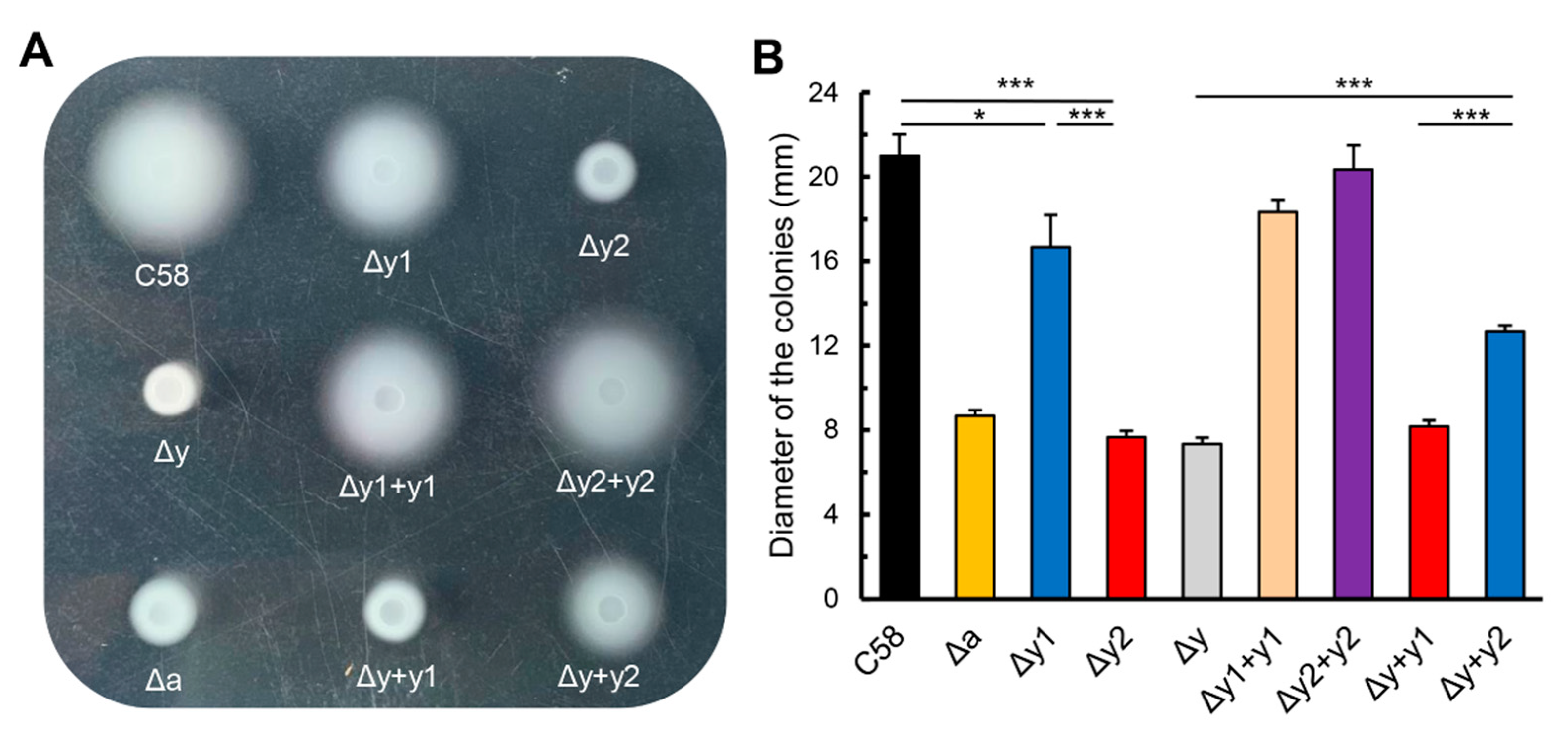
3.1. Deletion of Either cheY Gene Significantly Affects the Chemotactic Migration of A. fabrum in Swim Plate with cheY2 having Prominent Effect
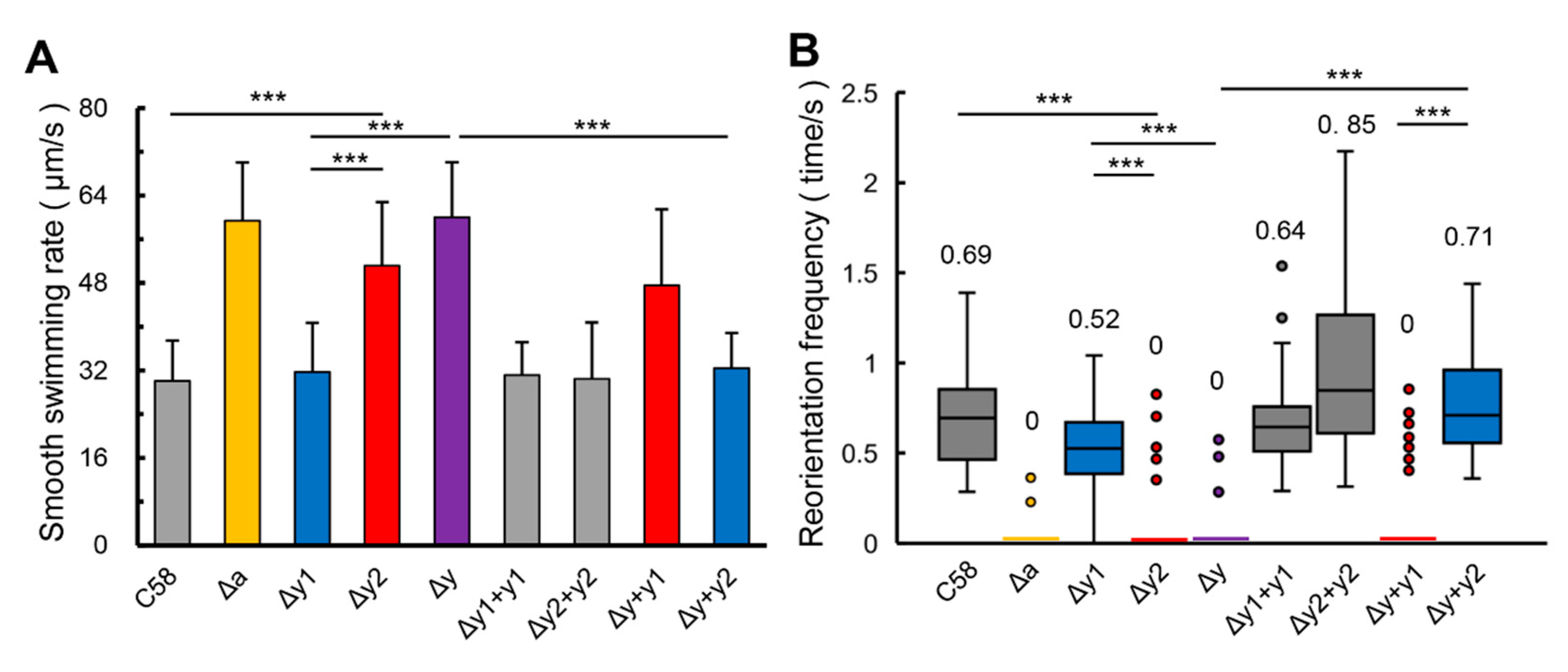
3.2. CheY1 and CheY2 have Different Effects on the Swimming Behavior of A. fabrum
3.3. Both CheY1 and CheY2 Interact with CheA at the Cell Pole Respectively
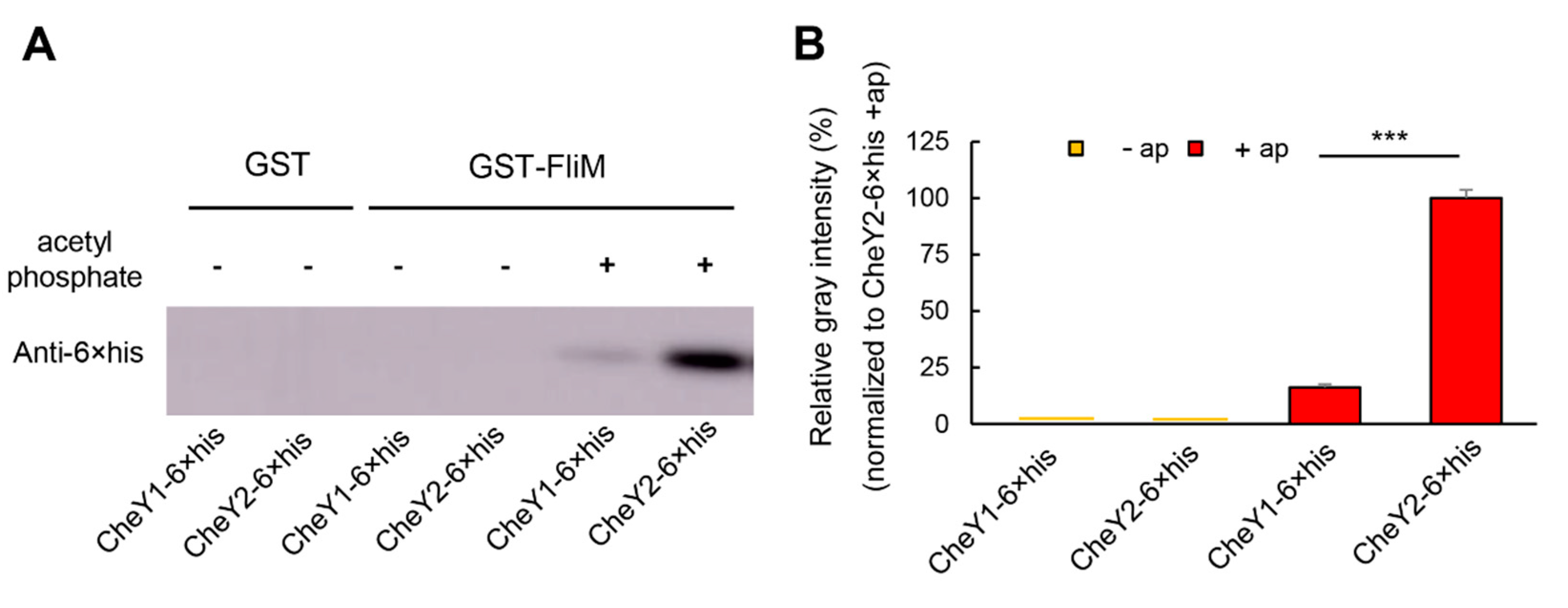
3.4. Two CheY Proteins Show Significantly Different Affinities to FliM
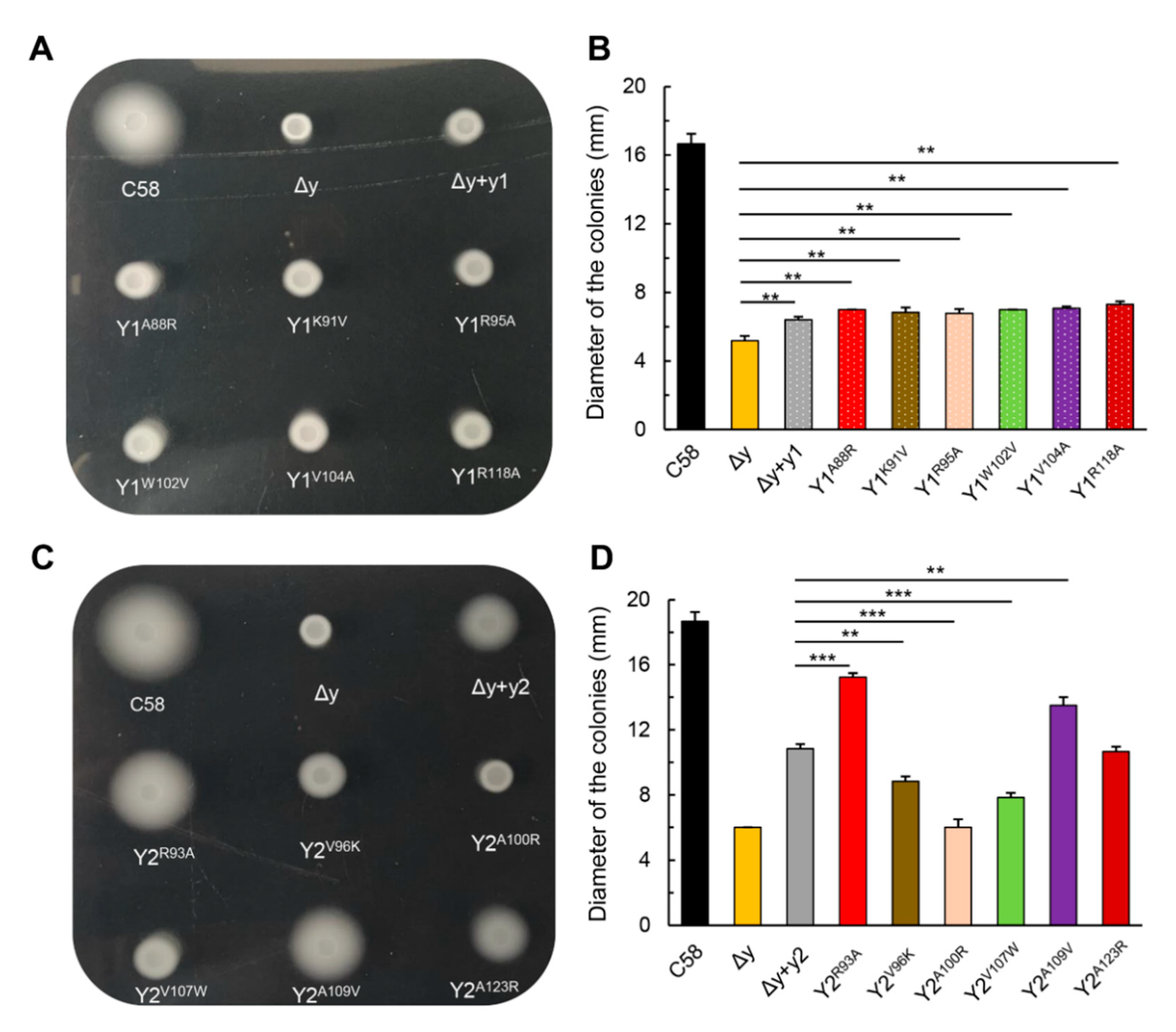
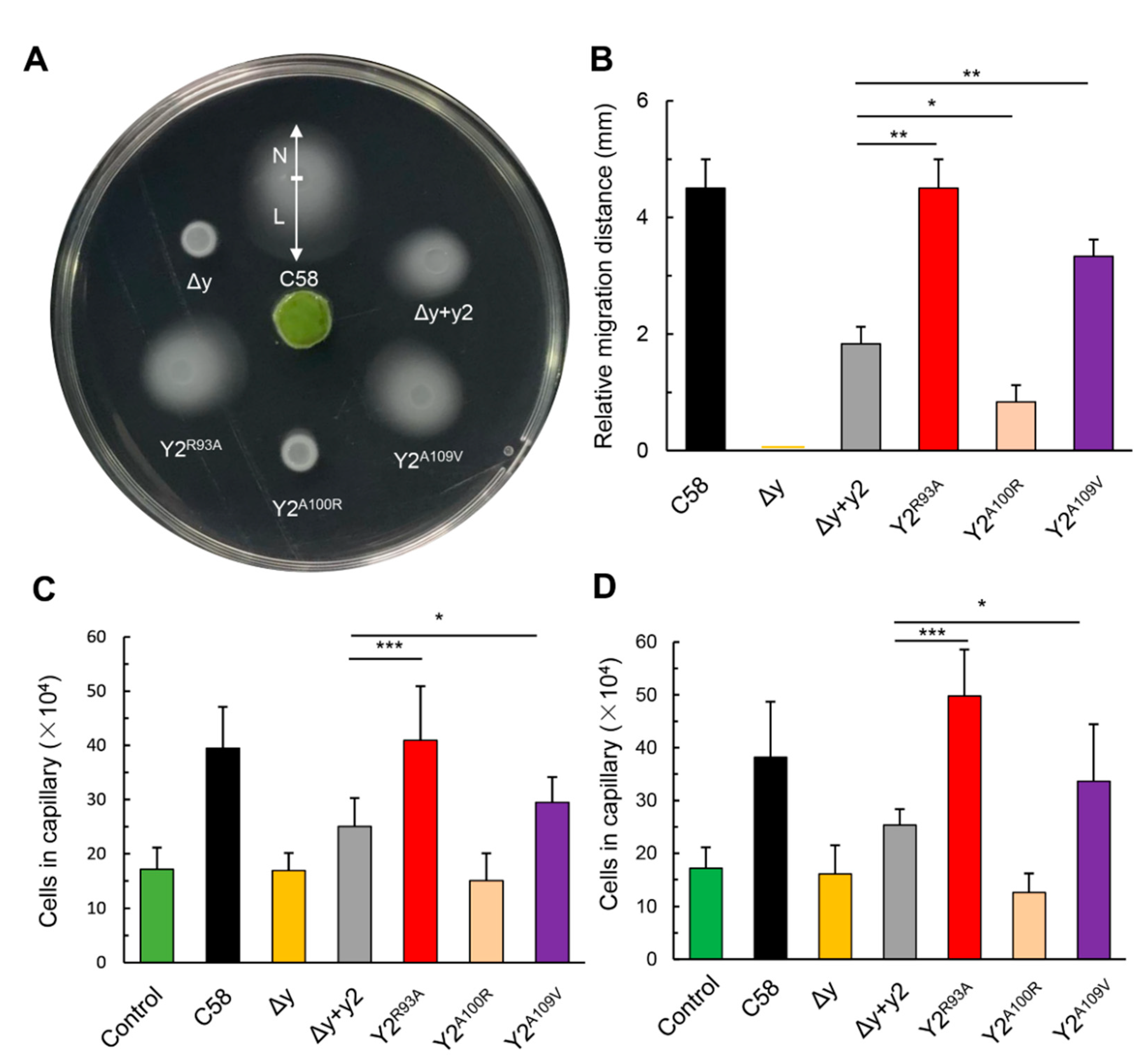
3.5. The Single Respective Replacements of R93 and A109 of CheY2 by the Residues of the Corresponding Sites in CheY1, Significantly Enhance the Function of CheY2 in the Absence of CheY1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acuna, G.; Shi, W.; Trudeau, K.; Zusman, D.R. The ‘CheA’ and ‘CheY’ domains of Myxococcus xanthus FrzE function independently in vitro as an autokinase and a phosphate acceptor, respectively. FEBS Lett. 1995, 358, 31–33. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, S.; Garzon, A.; Parkinson, J.S. Mutational analysis of the P1 phosphorylation domain in Escherichia coli CheA, the signaling kinase for chemotaxis. J. Bacteriol. 2014, 196, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Zschiedrich, C.P.; Keidel, V.; Szurmant, H. Molecular mechanisms of two-component signal transduction. J. Mol. Biol. 2016, 428, 3752–3775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quax, T.E.F.; Altegoer, F.; Rossi, F.; Li, Z.; Rodriguez-Franco, M.; Kraus, F.; Bange, G.; Albers, S. Structure and function of the archaeal response regulator CheY. Proc. Natl. Acad. Sci. USA 2018, 115, E1259–E1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazelbauer, G.L.; Laim, W. Bacterial chemoreceptors: Providing enhanced features to two-component signaling. Curr. Opin. Microbiol. 2010, 13, 124–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, S.; Lai, L. Bacterial chemoreceptors and chemoeffectors. Cell. Mol. Life. Sci. 2015, 72, 691–708. [Google Scholar] [CrossRef]
- Bren, A.; Eisenbach, M. The N terminus of the flagellar switch protein, FliM, is the binding domain for the chemotactic response regulator, CheY. J. Mol. Biol. 1998, 278, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, M.K.; Paul, K.; Blair, D. Chemotaxis signaling protein CheY binds to the rotor protein FliN to control the direction of flagellar rotation in Escherichia coli. Proc. Natl. Acad. Sci. USA 2010, 107, 9370–9375. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Huang, Z.; Yang, J. Is there any crosstalk between the chemotaxis and virulence induction signaling in Agrobacterium tumefaciens? Biotechnol. Adv. 2017, 35, 505–511. [Google Scholar] [CrossRef]
- Huang, Z.; Pan, X.; Xu, N.; Guo, M. Bacterial chemotaxis coupling protein: Structure, function and diversity. Microbiol. Res. 2019, 219, 40–48. [Google Scholar] [CrossRef]
- Wuichet, K.; Zhulin, I.B. Origins and diversification of a complex signal transduction system in prokaryotes. Sci. Signal. 2010, 3, ra50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motaleb, M.A.; Sultan, S.Z.; Miller, M.R.; Li, C.; Charon, N.W. CheY3 of Borrelia burgdorferi is the key response regulator essential for chemotaxis and forms a long-lived phosphorylated intermediate. J. Bacteriol. 2011, 193, 3332–3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Sultan, S.; Yerke, A.; Moon, K.H.; Mark Wooten, R.; Motaleb, M.A. Borrelia burgdorferi CheY2 is dispensable for chemotaxis or motility but crucial for the infectious life cycle of the spirochete. Infect. Immun. 2017, 85, e00264-e16. [Google Scholar] [CrossRef] [Green Version]
- Hyakutake, A.; Homma, M.; Austin, M.J.; Boin, M.A.; Hase, C.C.; Kawagishi, I. Only one of the five CheY homologs in Vibrio cholerae directly switches flagellar rotation. J. Bacteriol. 2005, 187, 8403–8410. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, T.; Kumar, D.; Burriss, N.; Xie, Z.; Alexandre, G. Azospirillum brasilense chemotaxis depends on two signaling pathways regulating distinct motility parameters. J. Bacteriol. 2016, 198, 1764–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, T.; Elmas, M.; Vo, L.; Alexiades, V.; Hong, T.; Alexandre, G. Multiple CheY homologs control swimming reversals and transient pauses in Azospirillum brasilense. Biophys. J. 2019, 116, 1527–1537. [Google Scholar] [CrossRef]
- Sourjik, V.; Schmitt, R. Different roles of CheY1 and CheY2 in the chemotaxis of Rhizobium meliloti. Mol. Microbiol. 1996, 22, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Bai, X.; Li, Y.; Min, J.; Kong, Y.; Hu, X. CheY1 and CheY2 of Azorhizobium caulinodans ORS571 regulate chemotaxis and competitive colonization with the host plant. Appl. Environ. Microbiol. 2020, 86, e00599-20. [Google Scholar] [CrossRef]
- Guo, M.; Bian, X.; Wu, X.; Wu, M. Agrobacterium-mediated genetic transformation: History and progress. In Genetic Transformation; Alvarez, M., Ed.; InTech: Rijeka, Croatia, 2011. [Google Scholar] [CrossRef] [Green Version]
- Kado, C.I. Historical account on gaining insights on the mechanism of crown gall tumorigenesis induced by Agrobacterium tumefaciens. Front. Microbiol. 2014, 5, 340. [Google Scholar] [CrossRef]
- Xu, N.; Wang, M.; Yang, X.; Xu, Y.; Guo, M. In silico analysis of the chemotactic system of Agrobacterium tumefaciens. Microb. Genom. 2020, 6, 000460. [Google Scholar] [CrossRef]
- Scharf, B.E.; Hynes, M.F.; Alexandre, G.M. Chemotaxis signaling systems in model beneficial plant–bacteria associations. Plant Mol. Biol. 2016, 90, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.D.; Yost, C.K.; Hynes, M.F.; Alexandre, G. The major chemotaxis gene cluster of Rhizobium leguminosarum bv. viciae is essential for competitive nodulation. Mol. Microbiol. 2007, 63, 348–362. [Google Scholar] [PubMed]
- Sourjik, V.; Schmitt, R. Phosphotransfer between CheA, CheY1, and CheY2 in the chemotaxis signal. Biochemistry 1998, 37, 2327–2335. [Google Scholar] [CrossRef]
- Dogra, G.; Purschke, F.G.; Wagner, V.; Haslbeck, M.; Kriehuber, T.; Hughes, J.G.; Van Tassell, M.L.; Gilbert, C.; Niemeyer, M.; Keith Ray, W.; et al. Sinorhizobium meliloti CheA Complexed with CheS Exhibits Enhanced Binding to CheY1, Resulting in Accelerated CheY1 Dephosphorylation. J. Bacteriol. 2011, 194, 1075–1087. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Maniatis, T.; Fritsch, E.F. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Huang, Z.; Zhou, Q.; Sun, P.; Yang, J.; Guo, M. Two Agrobacterium tumefaciens CheW proteins are incorporated into one chemosensory pathway with different efficiencies. Mol. Plant-Microbe Interact. 2018, 31, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelvin, S.B. Agrobacterium virulence gene induction. Methods Mol. Biol. 2006, 343, 77–84. [Google Scholar] [PubMed]
- Guo, M.; Hou, Q.M.; Hew, C.L.; Pan, S.Q. Agrobacterium VirD2-binding protein is involved in tumorigenesis and redundantly encoded in conjugative transfer gene clusters. Mol. Plant-Microbe Interact. 2007, 20, 1201–1212. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Zuo, J.; Gu, H.; Guo, M.; Yin, Y. Domain function dissection and catalytic properties of Listeria monocytogenes p60 protein with bacteriolytic activity. Appl. Microbiol. Biotechnol. 2015, 99, 10527–10537. [Google Scholar] [CrossRef]
- Yang, J.; Wu, M.; Zhang, X.; Guo, M.; Huang, Z. Expression of Agrobacterium homolog genes encoding T-complex recruiting protein under virulence induction conditions. Front. Microbiol. 2015, 6, 1379. [Google Scholar] [CrossRef]
- Goodner, B.; Hinkle, G.; Gattung, S.; Miller, N.; Blanchard, M.; Qurollo, B.; Goldman, B.S.; Cao, Y.; Askenazi, M.; Halling, C.; et al. Genome sequence of the plant pathogen and biotechnology agent Agrobacterium tumefaciens C58. Science 2001, 294, 2323–2328. [Google Scholar] [CrossRef]
- Wood, D.W.; Setubal, J.C.; Kaul, R.; Monks, D.E.; Kitajima, J.P.; Okura, V.K.; Zhou, Y.; Chen, L.; Wood, G.E.; Almeida, N.F., Jr.; et al. The genome of the natural genetic engineer Agrobacterium tumefaciens C58. Science 2001, 294, 2317–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Jin, S.; Sun, D.; Hew, C.L.; Pan, S.Q. Recruitment of conjugative DNA transfer substrate to Agrobacterium type IV secretion apparatus. Proc. Natl. Acad. Sci. USA 2007, 104, 20019–20024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Pan, X.; Xu, Y.; Li, Y.; Xu, N.; Huang, Z.; Ye, J.; Gao, D.; Guo, M. Agrobacterium tumefaciens ferritins play an important role in full virulence through regulating iron homeostasis and oxidative stress survival. Mol. Plant. Pathol. 2020, 21, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Zhu, Q.; Gao, D. Development and optimization of method for generating unmarked A. tumefaciens mutants. Prog. Biochem. Biophys. 2009, 36, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Cangelosi, G.A.; Best, E.A.; Martinetti, G.; Nester, E.W. Genetic analysis of Agrobacterium. Methods Enzymol. 1991, 204, 384–397. [Google Scholar]
- Sampedro, I.; Parales, R.E.; Krell, T.; Hill, J.E. Pseudomonas chemotaxis. FEMS Microbiol. Rev. 2015, 39, 17–46. [Google Scholar]
- Roggo, C.; Carraro, N.; Meer, J.R.V.D. Probing chemotaxis activity in Escherichia coli using fluorescent protein fusions. Sci. Rep. 2019, 9, 3845. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Bian, X.; Guo, M.; Wang, J.; Zhang, X. Identification and characterization of the biochemical function of Agrobacterium T-complex-recruiting protein Atu5117. FEBS J. 2013, 280, 4865–4875. [Google Scholar] [CrossRef]
- Volz, K.; Matsumura, P. Crystal structure of Escherichia coli CheY refined at 1.7-Å resolution. J. Biol. Chem. 1991, 266, 15511–15519. [Google Scholar] [CrossRef]
- Lee, S.; Cho, H.S.; Pelton, J.G.; Yan, D.; Henderson, R.K.; King, D.S.; Huang, L.; Kustu, S.; Berry, E.A.; Wemmer, D.E. Crystal structure of an activated response regulator bound to its target. Nat. Struct. Biol. 2001, 8, 52–56. [Google Scholar] [CrossRef]
- Lam, K.H.; Ling, T.K.W.; Au, S.W.N. Crystal structure of activated CheY1 from Helicobacter pylori. J. Bacteriol. 2010, 192, 2324–2334. [Google Scholar] [CrossRef] [Green Version]
- Thakor, H.; Nicholas, S.; Porter, I.M.; Hand, N.; Stewart, R.C. Identification of an anchor residue for CheA-CheY interactions in the chemotaxis system of Escherichia coli. J. Bacteriol. 2011, 193, 3894–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harighi, B. Role of CheY1 and CheY2 in the chemotaxis of A. tumefaciens toward acetosyringone. Curr. Microbiol. 2008, 56, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Dyer, C.M.; Dahlquist, F.W. Switched or not? The structure of unphosphorylated CheY bound to the N terminus of FliM. J. Bacteriol. 2006, 188, 7354–7363. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Stephens, B.B.; Alexandre, G.; Farrand, S.K. Lon protease of the α-proteobacterium Agrobacterium tumefaciens is required for normal growth, cellular morphology and full virulence. Microbiology 2006, 152, 1197–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riepl, H.; Maurer, T.; Kalbitzer, H.R.; Meier, V.M.; Haslbeck, M.; Schmitt, R.; Scharf, B. Interaction of CheY2 and CheY2-P with the cognate CheA kinase in the chemosensory-signalling chain of Sinorhizobium meliloti. Mol. Microbiol. 2008, 69, 1373–1384. [Google Scholar] [CrossRef] [PubMed]
- Cluzel, P.; Surette, M.; Leibler, S. An ultrasensitive bacterial motor revealed by monitoring signaling proteins in single cells. Science 2000, 287, 1652–1655. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Branch, R.W.; Hosu, B.G.; Berg, H.C. Adaptation at the output of the chemotaxis signalling pathway. Nature 2012, 484, 233–235. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Tao, A.; Zhang, R.; Yuan, J. Robustness in an ultrasensitive motor. mBio 2020, 11, e03050-19. [Google Scholar] [CrossRef] [Green Version]
- Hawes, M.C.; Smith, L.Y. Requirement for chemotaxis in pathogenicity of Agrobacterium tumefaciens on roots of soil-grown pea plants. J. Bacteriol. 1989, 171, 5668–5671. [Google Scholar] [CrossRef] [Green Version]
- Merritt, P.M.; Danhorn, T.; Fuqua, C. Motility and chemotaxis in Agrobacterium tumefaciens surface attachment and biofilm formation. J. Bacteriol. 2007, 189, 8005–8014. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, D.; Zong, R.; Huang, Z.; Ye, J.; Wang, H.; Xu, N.; Guo, M. The Divergent Key Residues of Two Agrobacterium fabrum (tumefaciens) CheY Paralogs Play a Key Role in Distinguishing Their Functions. Microorganisms 2021, 9, 1134. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061134
Gao D, Zong R, Huang Z, Ye J, Wang H, Xu N, Guo M. The Divergent Key Residues of Two Agrobacterium fabrum (tumefaciens) CheY Paralogs Play a Key Role in Distinguishing Their Functions. Microorganisms. 2021; 9(6):1134. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061134
Chicago/Turabian StyleGao, Dawei, Renjie Zong, Zhiwei Huang, Jingyang Ye, Hao Wang, Nan Xu, and Minliang Guo. 2021. "The Divergent Key Residues of Two Agrobacterium fabrum (tumefaciens) CheY Paralogs Play a Key Role in Distinguishing Their Functions" Microorganisms 9, no. 6: 1134. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061134