
Effect of Coxsackievirus B4 Infection on the Thymus: Elucidating Its Role in the Pathogenesis of Type 1 Diabetes

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Thymic Structure and Function
2.1. Thymic Structure
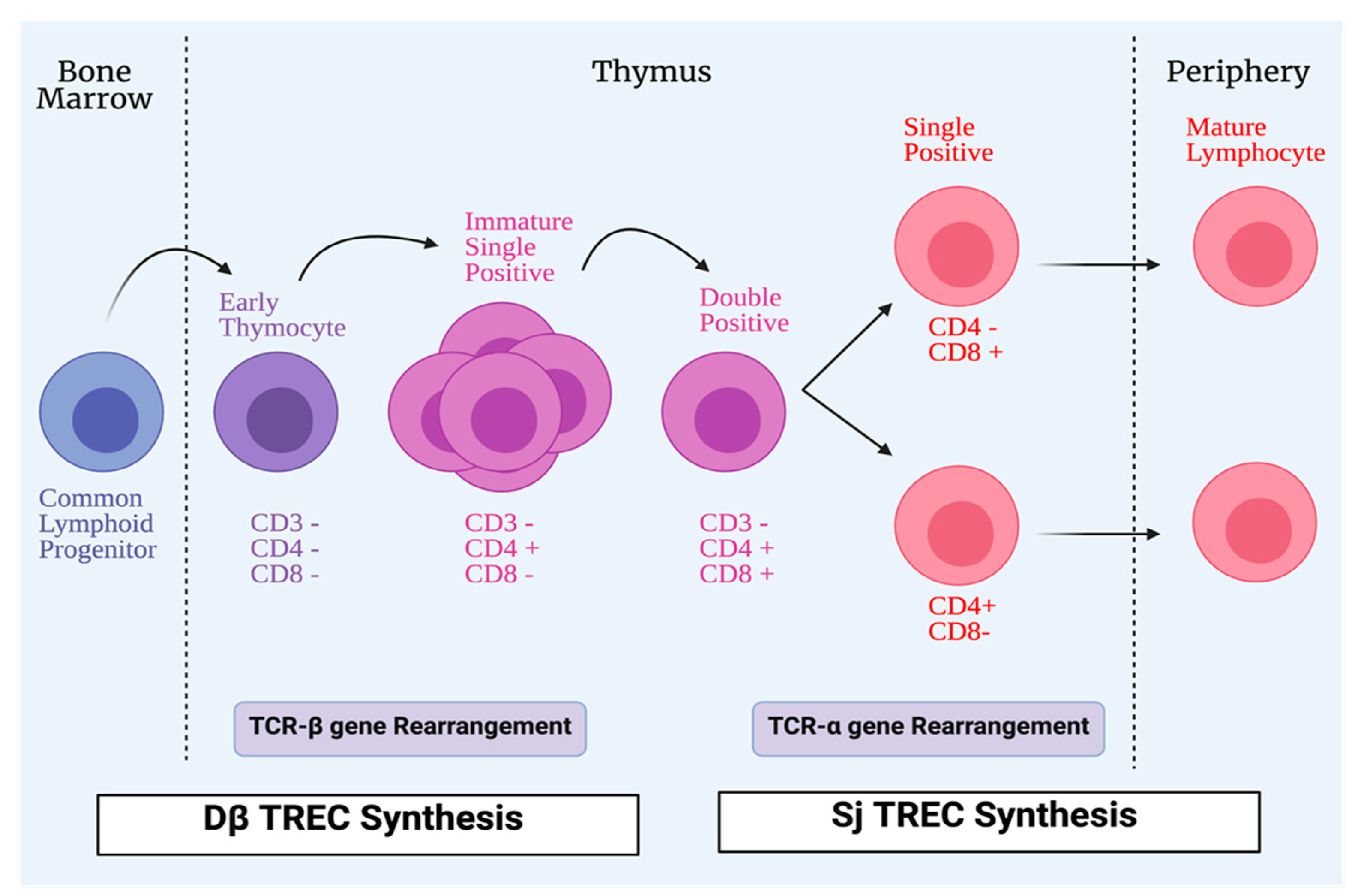
2.2. T-Cells Maturation and Differentiation
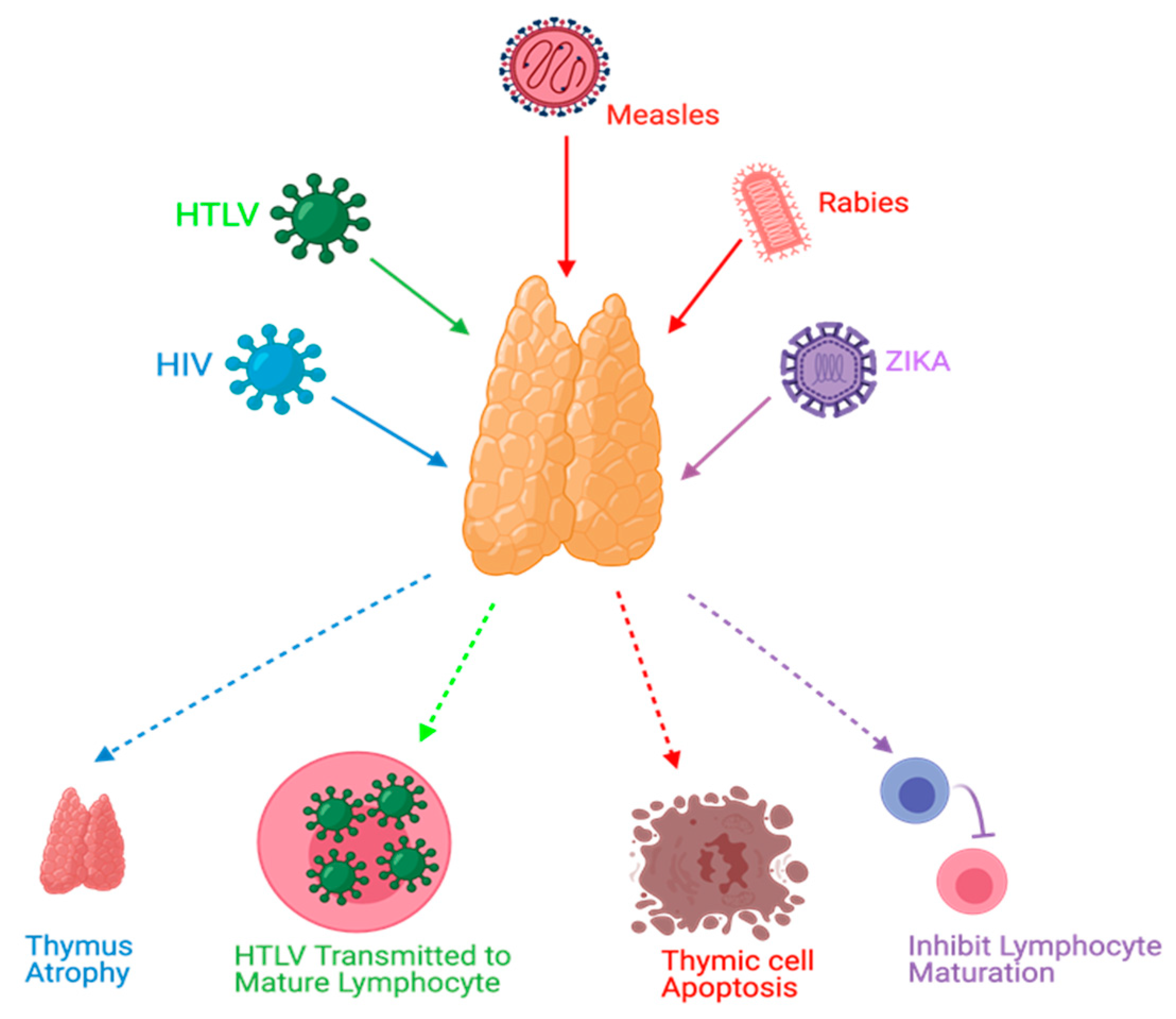
3. Viral Infections and the Thymus Gland
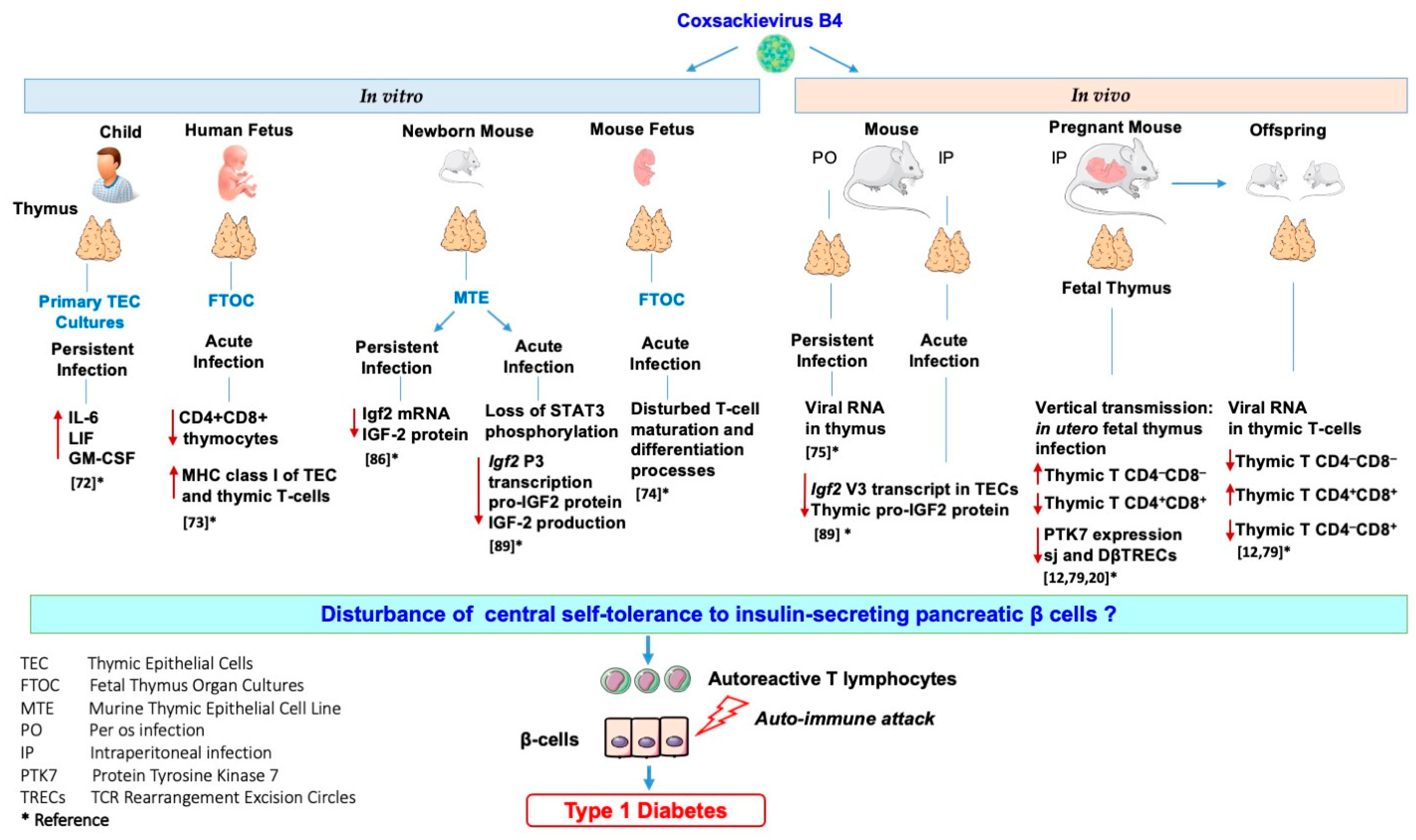
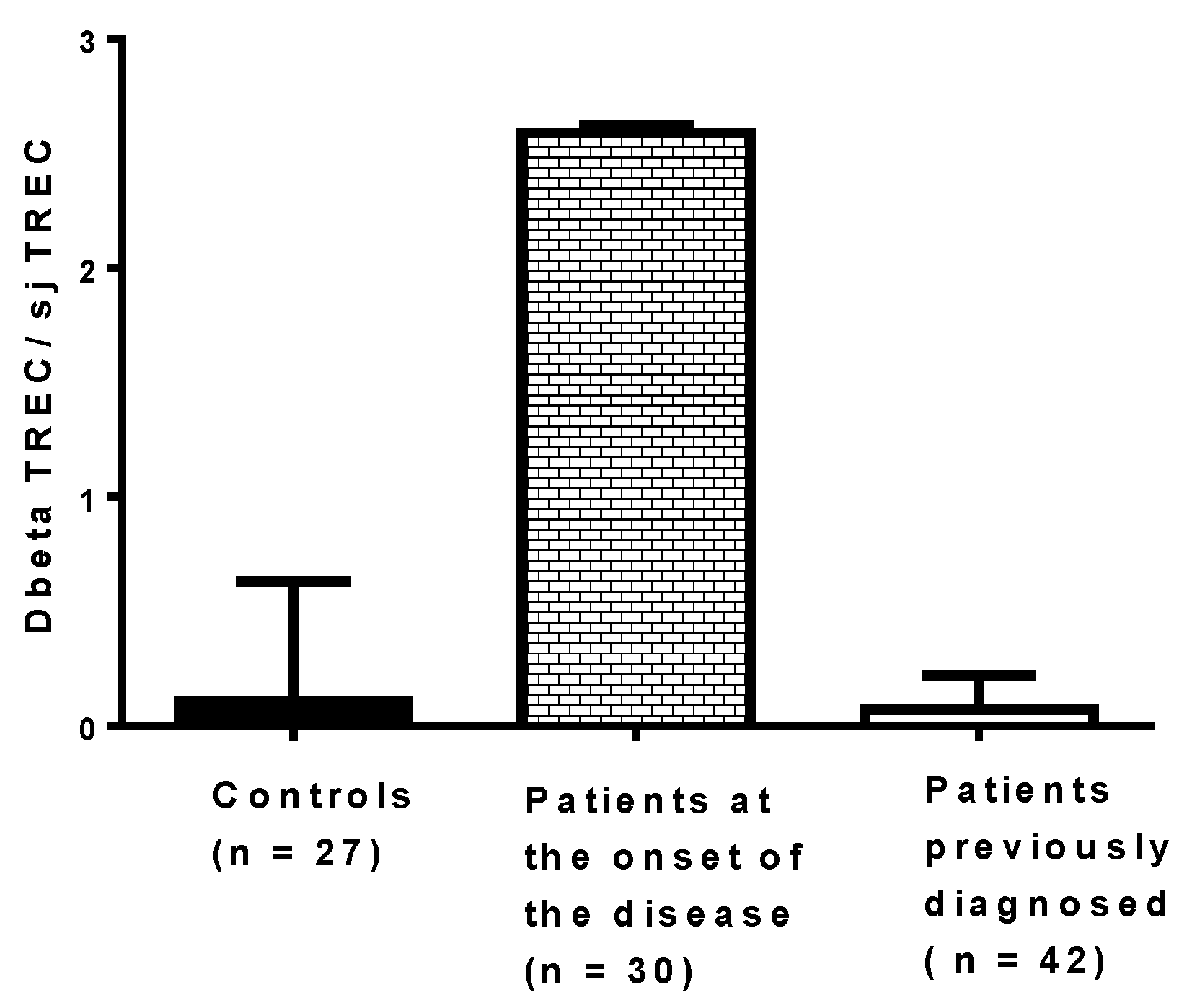
4. Coxsackievirus B4, the Thymus Gland, and Type 1 Diabetes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus Taxonomy Profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Tapparel, C.; Siegrist, F.; Petty, T.J.; Kaiser, L. Picornavirus and enterovirus diversity with associated human diseases. Infect. Genet. Evol. 2013, 14, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, M.H. Enterovirus infections: Diagnosis and treatment. Semin. Pediatr. Infect. Dis. 2002, 13, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Alhazmi, A.; Sane, F.; Lazrek, M.; Nekoua, M.P.; Badia-Boungou, F.; Engelmann, I.; Alidjinou, E.K.; Hober, D. Enteroviruses and Type 1 Diabetes Mellitus: An Overlooked Relationship in Some Regions. Microorganisms 2020, 8, 1458. [Google Scholar] [CrossRef]
- Stewart, T.A.; Hultgren, B.; Huang, X.; Pitts-Meek, S.; Hully, J.; MacLachlan, N.J. Induction of type I diabetes by interferon-alpha in transgenic mice. Science 1993, 260, 1942–1946. [Google Scholar] [CrossRef]
- Horwitz, M.S.; Bradley, L.M.; Harbertson, J.; Krahl, T.; Lee, J.; Sarvetnick, N. Diabetes induced by Coxsackie virus: Initiation by bystander damage and not molecular mimicry. Nat. Med. 1998, 4, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Filippi, C.; von Herrath, M. How viral infections affect the autoimmune process leading to type 1 diabetes. Cell. Immunol. 2005, 233, 125–132. [Google Scholar] [CrossRef]
- Von Herrath, M.G.; Fujinami, R.S.; Whitton, J.L. Microorganisms and autoimmunity: Making the barren field fertile? Nat. Rev. Microbiol. 2003, 1, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Albano, F.; Vecchio, E.; Renna, M.; Iaccino, E.; Mimmi, S.; Caiazza, C.; Arcucci, A.; Avagliano, A.; Pagliara, V.; Donato, G.; et al. Insights into Thymus Development and Viral Thymic Infections. Viruses 2019, 11, 836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alidjinou, E.K.; Sané, F.; Engelmann, I.; Geenen, V.; Hober, D. Enterovirus persistence as a mechanism in the pathogenesis of type 1 diabetes. Discov. Med. 2014, 18, 273–282. [Google Scholar]
- Tracy, S.; Smithee, S.; Alhazmi, A.; Chapman, N. Coxsackievirus can persist in murine pancreas by deletion of 5′ terminal genomic sequences. J. Med. Virol. 2014, 87, 240–247. [Google Scholar] [CrossRef]
- Jaïdane, H.; Halouani, A.; Jmii, H.; Elmastour, F.; Abdelkefi, S.; Bodart, G.; Michaux, H.; Chakroun, T.; Sane, F.; Mokni, M.; et al. In-utero coxsackievirus B4 infection of the mouse thymus. Clin. Exp. Immunol. 2017, 187, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.X.; Pan, W.; Zheng, L.; Zhong, X.P.; Tan, L.; Liang, Z.; He, J.; Feng, P.; Zhao, Y.; Qiu, Y.R. Thymic Epithelial Cells Contribute to Thymopoiesis and T Cell Development. Front. Immunol. 2019, 10, 3099. [Google Scholar] [CrossRef] [PubMed]
- Zdrojewicz, Z.; Pachura, E.; Pachura, P. The Thymus: A Forgotten, but Very Important Organ. Adv. Clin. Exp. Med. 2016, 25, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.F. The golden anniversary of the thymus. Nat. Rev. Immunol. 2011, 11, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Takaba, H.; Takayanagi, H. The Mechanisms of T Cell Selection in the Thymus. Trends Immunol. 2017, 38, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Bleul, C.C.; Corbeaux, T.; Reuter, A.; Fisch, P.; Mönting, J.S.; Boehm, T. Formation of a functional thymus initiated by a postnatal epithelial progenitor cell. Nature 2006, 441, 992–996. [Google Scholar] [CrossRef]
- Al-Harthi, L.; Marchetti, G.; Steffens, C.M.; Poulin, J.; Sékaly, R.; Landay, A. Detection of T cell receptor circles (TRECs) as biomarkers for de novo T cell synthesis using a quantitative polymerase chain reaction-enzyme linked immunosorbent assay (PCR-ELISA). J. Immunol. Methods 2000, 237, 187–197. [Google Scholar] [CrossRef]
- De Voeght, A.; Martens, H.; Renard, C.; Vaira, D.; Debruche, M.; Simonet, J.; Geenen, V.; Moutschen, M.; Darcis, G. Exploring the link between innate immune activation and thymic function by measuring sCD14 and TRECs in HIV patients living in Belgium. PLoS ONE 2017, 12, e0185761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halouani, A.; Jmii, H.; Bodart, G.; Michaux, H.; Renard, C.; Martens, H.; Aouni, M.; Hober, D.; Geenen, V.; Jaïdane, H. Assessment of Thymic Output Dynamics After. Front. Immunol. 2020, 11, 481. [Google Scholar] [CrossRef]
- Ferrando-Martinez, S.; De Pablo-Bernal, R.S.; De Luna-Romero, M.; De Ory, S.J.; Genebat, M.; Pacheco, Y.M.; Parras, F.J.; Montero, M.; Blanco, J.R.; Gutierrez, F.; et al. Thymic Function Failure Is Associated With Human Immunodeficiency Virus Disease Progression. Clin. Infect. Dis. 2017, 64, 1191–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, L.; Kyewski, B. “Promiscuous” expression of tissue antigens in the thymus: A key to T-cell tolerance and autoimmunity? J. Mol. Med. 2000, 78, 483–494. [Google Scholar] [CrossRef]
- Derbinski, J.; Schulte, A.; Kyewski, B.; Klein, L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol. 2001, 2, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Martens, H.; Goxe, B.; Geenen, V. The thymic repertoire of neuroendocrine self-antigens: Physiological implications in T-cell life and death. Immunol. Today 1996, 17, 312–317. [Google Scholar] [CrossRef]
- Kyewski, B.; Klein, L. A central role for central tolerance. Annu. Rev. Immunol. 2006, 24, 571–606. [Google Scholar] [CrossRef] [PubMed]
- Thomas-Vaslin, V.; Damotte, D.; Coltey, M.; Le Douarin, N.M.; Coutinho, A.; Salaün, J. Abnormal T cell selection on nod thymic epithelium is sufficient to induce autoimmune manifestations in C57BL/6 athymic nude mice. Proc. Natl. Acad. Sci. USA 1997, 94, 4598–4603. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, H.; Sprent, J. A defect in central tolerance in NOD mice. Nat. Immunol. 2001, 2, 1025–1031. [Google Scholar] [CrossRef]
- Rosado-Sánchez, I.; Herrero-Fernández, I.; Genebat, M.; Ruiz-Mateos, E.; Leal, M.; Pacheco, Y.M. Thymic Function Impacts the Peripheral CD4/CD8 Ratio of HIV-Infected Subjects. Clin. Infect. Dis. 2017, 64, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Fiume, G.; Scialdone, A.; Albano, F.; Rossi, A.; Tuccillo, F.M.; Rea, D.; Palmieri, C.; Caiazzo, E.; Cicala, C.; Bellevicine, C.; et al. Impairment of T cell development and acute inflammatory response in HIV-1 Tat transgenic mice. Sci. Rep. 2015, 5, 13864. [Google Scholar] [CrossRef] [Green Version]
- Hartling, H.J.; Gaardbo, J.C.; Ronit, A.; Salem, M.; Laye, M.; Clausen, M.R.; Skogstrand, K.; Gerstoft, J.; Ullum, H.; Nielsen, S.D. Impaired thymic output in patients with chronic hepatitis C virus infection. Scand. J. Immunol. 2013, 78, 378–386. [Google Scholar] [CrossRef]
- Ye, P.; Kirschner, D.E.; Kourtis, A.P. The thymus during HIV disease: Role in pathogenesis and in immune recovery. Curr. HIV Res. 2004, 2, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Rb-Silva, R.; Nobrega, C.; Azevedo, C.; Athayde, E.; Canto-Gomes, J.; Ferreira, I.; Cheynier, R.; Yates, A.J.; Horta, A.; Correia-Neves, M. Thymic Function as a Predictor of Immune Recovery in Chronically HIV-Infected Patients Initiating Antiretroviral Therapy. Front. Immunol. 2019, 10, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho Barros, L.R.; Linhares-Lacerda, L.; Moreira-Ramos, K.; Ribeiro-Alves, M.; Machado Motta, M.C.; Bou-Habib, D.C.; Savino, W. HTLV-1-infected thymic epithelial cells convey the virus to CD4. Immunobiology 2017, 222, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Cardenas Palomo, L.F.; de Souza Matos, D.C.; Chaves Leal, E.; Bertho, A.L.; Marcovistz, R. Lymphocyte subsets and cell proliferation analysis in rabies-infected mice. J. Clin. Lab. Immunol. 1995, 46, 49–61. [Google Scholar] [PubMed]
- Marcovistz, R.; Bertho, A.L.; Matos, D.C. Relationship between apoptosis and thymocyte depletion in rabies-infected mice. Braz. J. Med. Biol. Res. 1994, 27, 1599–1603. [Google Scholar]
- Wiktor, T.J.; Doherty, P.C.; Koprowski, H. Suppression of cell-mediated immunity by street rabies virus. J. Exp. Med. 1977, 145, 1617–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirai, K.; Kawano, H.; Mifune, K.; Fujii, H.; Nishizono, A.; Shichijo, A.; Mannen, K. Suppression of cell-mediated immunity by street rabies virus infection. Microbiol. Immunol. 1992, 36, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Kasempimolporn, S.; Saengseesom, W.; Mitmoonpitak, C.; Akesowan, S.; Sitprija, V. Cell-mediated immunosuppression in mice by street rabies virus not restored by calcium ionophore or PMA. Asian Pac. J. Allergy Immunol. 1997, 15, 127–132. [Google Scholar]
- Godfraind, C.; Holmes, K.V.; Coutelier, J.P. Thymus involution induced by mouse hepatitis virus A59 in BALB/c mice. J. Virol. 1995, 69, 6541–6547. [Google Scholar] [CrossRef] [Green Version]
- Auwaerter, P.G.; Kaneshima, H.; McCune, J.M.; Wiegand, G.; Griffin, D.E. Measles virus infection of thymic epithelium in the SCID-hu mouse leads to thymocyte apoptosis. J. Virol. 1996, 70, 3734–3740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentin, H.; Azocar, O.; Horvat, B.; Williems, R.; Garrone, R.; Evlashev, A.; Toribio, M.L.; Rabourdin-Combe, C. Measles virus infection induces terminal differentiation of human thymic epithelial cells. J. Virol. 1999, 73, 2212–2221. [Google Scholar] [CrossRef] [Green Version]
- Yamanouchi, K.; Chino, F.; Kobune, F.; Kodama, H.; Tsuruhara, T. Growth of measles virus in the lymphoid tissues of monkeys. J. Infect. Dis. 1973, 128, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Moench, T.R.; Griffin, D.E.; Obriecht, C.R.; Vaisberg, A.J.; Johnson, R.T. Acute measles in patients with and without neurological involvement: Distribution of measles virus antigen and RNA. J. Infect. Dis. 1988, 158, 433–442. [Google Scholar] [CrossRef]
- Gibb, T.R.; Bray, M.; Geisbert, T.W.; Steele, K.E.; Kell, W.M.; Davis, K.J.; Jaax, N.K. Pathogenesis of experimental Ebola Zaire virus infection in BALB/c mice. J. Comp. Pathol. 2001, 125, 233–242. [Google Scholar] [CrossRef]
- Mocarski, E.S.; Bonyhadi, M.; Salimi, S.; McCune, J.M.; Kaneshima, H. Human cytomegalovirus in a SCID-hu mouse: Thymic epithelial cells are prominent targets of viral replication. Proc. Natl. Acad. Sci. USA 1993, 90, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messias, C.V.; Loss-Morais, G.; Carvalho, J.B.; González, M.N.; Cunha, D.P.; Vasconcelos, Z.; Arge, L.W.P.; Farias-de-Oliveira, D.A.; Gerber, A.L.; Portari, E.A.; et al. Zika virus targets the human thymic epithelium. Sci. Rep. 2020, 10, 1378. [Google Scholar] [CrossRef] [PubMed]
- Gul, K.A.; Sonerud, T.; Fjærli, H.O.; Nakstad, B.; Abrahamsen, T.G.; Inchley, C.S. Thymus activity measured by T-cell receptor excision circles in patients with different severities of respiratory syncytial virus infection. BMC Infect. Dis. 2017, 17, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.F.; Wang, H. Environmental Exposures and Autoimmune Diseases: Contribution of Gut Microbiome. Front. Immunol. 2019, 10, 3094. [Google Scholar] [CrossRef]
- Khan, M.F.; Wang, G. Environmental Agents, Oxidative Stress and Autoimmunity. Curr. Opin. Toxicol. 2018, 7, 22–27. [Google Scholar] [CrossRef]
- Shukla, S.K.; Singh, G.; Ahmad, S.; Pant, P. Infections, genetic and environmental factors in pathogenesis of autoimmune thyroid diseases. Microb. Pathog. 2018, 116, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Giwa, A.M.; Ahmed, R.; Omidian, Z.; Majety, N.; Karakus, K.E.; Omer, S.M.; Donner, T.; Hamad, A.R.A. Current understandings of the pathogenesis of type 1 diabetes: Genetics to environment. World J. Diabetes 2020, 11, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, K.A.; Hitman, G.A.; Rowe, R.E.; Hawa, M.; Huang, X.; Stewart, T.; Leslie, R.D. Concordance for type 1 diabetes in identical twins is affected by insulin genotype. Diabetes Care 2001, 24, 838–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaïdane, H.; Sané, F.; Hiar, R.; Goffard, A.; Gharbi, J.; Geenen, V.; Hober, D. Immunology in the clinic review series; focus on type 1 diabetes and viruses: Enterovirus, thymus and type 1 diabetes pathogenesis. Clin. Exp. Immunol. 2012, 168, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Hober, D.; Sane, F. Enteroviral pathogenesis of type 1 diabetes. Discov. Med. 2010, 10, 151–160. [Google Scholar] [CrossRef]
- Hober, D.; Sauter, P. Pathogenesis of type 1 diabetes mellitus: Interplay between enterovirus and host. Nat. Rev. Endocrinol. 2010, 6, 279–289. [Google Scholar] [CrossRef]
- Hober, D.; Alidjinou, E.K. Enteroviral pathogenesis of type 1 diabetes: Queries and answers. Curr. Opin. Infect. Dis. 2013, 26, 263–269. [Google Scholar] [CrossRef]
- Chehadeh, W.; Kerr-Conte, J.; Pattou, F.; Alm, G.; Lefebvre, J.; Wattré, P.; Hober, D. Persistent infection of human pancreatic islets by coxsackievirus B is associated with alpha interferon synthesis in beta cells. J. Virol. 2000, 74, 10153–10164. [Google Scholar] [CrossRef] [Green Version]
- Sane, F.; Caloone, D.; Gmyr, V.; Engelmann, I.; Belaich, S.; Kerr-Conte, J.; Pattou, F.; Desailloud, R.; Hober, D. Coxsackievirus B4 can infect human pancreas ductal cells and persist in ductal-like cell cultures which results in inhibition of Pdx1 expression and disturbed formation of islet-like cell aggregates. Cell. Mol. Life Sci. 2013, 70, 4169–4180. [Google Scholar] [CrossRef]
- Bertin, A.; Sane, F.; Gmyr, V.; Lobert, D.; Dechaumes, A.; Kerr-Conte, J.; Pattou, F.; Hober, D. Coxsackievirus B4 Infection of Human Primary Pancreatic Ductal Cell Cultures Results in Impairment of Differentiation into Insulin-Producing Cells. Viruses 2019, 11, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alidjinou, E.K.; Sané, F.; Engelmann, I.; Hober, D. Serum-dependent enhancement of coxsackievirus B4-induced production of IFNα, IL-6 and TNFα by peripheral blood mononuclear cells. J. Mol. Biol. 2013, 425, 5020–5031. [Google Scholar] [CrossRef] [PubMed]
- Alidjinou, E.K.; Chehadeh, W.; Weill, J.; Vantyghem, M.C.; Stuckens, C.; Decoster, A.; Hober, C.; Hober, D. Monocytes of Patients with Type 1 Diabetes Harbour Enterovirus RNA. Eur. J. Clin. Investig. 2015, 45, 918–924. [Google Scholar] [CrossRef]
- Alidjinou, E.K.; Sané, F.; Trauet, J.; Copin, M.C.; Hober, D. Coxsackievirus B4 Can Infect Human Peripheral Blood-Derived Macrophages. Viruses 2015, 7, 6067–6079. [Google Scholar] [CrossRef] [Green Version]
- Benkahla, M.A.; Elmastour, F.; Sane, F.; Vreulx, A.C.; Engelmann, I.; Desailloud, R.; Jaidane, H.; Alidjinou, E.K.; Hober, D. Coxsackievirus B4E2 can infect monocytes and macrophages in vitro and in vivo. Virology 2018, 522, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Hober, D.; Chehadeh, W.; Weill, J.; Hober, C.; Vantyghem, M.C.; Gronnier, P.; Wattré, P. Circulating and cell-bound antibodies increase coxsackievirus B4-induced production of IFN-alpha by peripheral blood mononuclear cells from patients with type 1 diabetes. J. Gen. Virol. 2002, 83, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Chehadeh, W.; Lobert, P.E.; Sauter, P.; Goffard, A.; Lucas, B.; Weill, J.; Vantyghem, M.C.; Alm, G.; Pigny, P.; Hober, D. Viral protein VP4 is a target of human antibodies enhancing coxsackievirus B4- and B3-induced synthesis of alpha interferon. J. Virol. 2005, 79, 13882–13891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hober, D.; Sane, F.; Jaïdane, H.; Riedweg, K.; Goffard, A.; Desailloud, R. Immunology in the clinic review series; focus on type 1 diabetes and viruses: Role of antibodies enhancing the infection with Coxsackievirus B in the pathogenesis of type 1 diabetes. Clin. Exp. Immunol. 2012, 168, 47–51. [Google Scholar] [CrossRef]
- Elmastour, F.; Jaïdane, H.; Benkahla, M.; Aguech-Oueslati, L.; Sane, F.; Halouani, A.; Engelmann, I.; Bertin, A.; Mokni, M.; Gharbi, J.; et al. Anti-coxsackievirus B4 (CVB4) enhancing activity of serum associated with increased viral load and pathology in mice reinfected with CVB4. Virulence 2017, 8, 908–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmastour, F.; Jaidane, H.; Aguech-Oueslati, L.; Benkahla, M.A.; Aouni, M.; Gharbi, J.; Sane, F.; Hober, D. Immunoglobulin G-dependent enhancement of the infection with Coxsackievirus B4 in a murine system. Virulence 2016, 7, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brusko, T.M.; Wasserfall, C.H.; Clare-Salzler, M.J.; Schatz, D.A.; Atkinson, M.A. Functional defects and the influence of age on the frequency of CD4+ CD25+ T-cells in type 1 diabetes. Diabetes 2005, 54, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.S.; von Herrath, M. CD4 T cell differentiation in type 1 diabetes. Clin. Exp. Immunol. 2016, 183, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Nekoua, M.P.; Fachinan, R.; Fagninou, A.; Alidjinou, E.K.; Moutairou, K.; Hober, D.; Yessoufou, A. Does control of glycemia regulate immunological parameters in insulin-treated persons with type 1 diabetes? Diabetes Res. Clin. Pract. 2019, 157, 107868. [Google Scholar] [CrossRef]
- Brilot, F.; Chehadeh, W.; Charlet-Renard, C.; Martens, H.; Geenen, V.; Hober, D. Persistent infection of human thymic epithelial cells by coxsackievirus B4. J. Virol. 2002, 76, 5260–5265. [Google Scholar] [CrossRef] [Green Version]
- Brilot, F.; Geenen, V.; Hober, D.; Stoddart, C.A. Coxsackievirus B4 infection of human fetal thymus cells. J. Virol. 2004, 78, 9854–9861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brilot, F.; Jaïdane, H.; Geenen, V.; Hober, D. Coxsackievirus B4 infection of murine foetal thymus organ cultures. J. Med. Virol. 2008, 80, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaïdane, H.; Gharbi, J.; Lobert, P.E.; Lucas, B.; Hiar, R.; M’hadheb, M.B.; Brilot, F.; Geenen, V.; Aouni, M.; Hober, D. Prolonged viral RNA detection in blood and lymphoid tissues from coxsackievirus B4 E2 orally-inoculated Swiss mice. Microbiol. Immunol. 2006, 50, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Euscher, E.; Davis, J.; Holzman, I.; Nuovo, G.J. Coxsackie virus infection of the placenta associated with neurodevelopmental delays in the newborn. Obstet. Gynecol. 2001, 98, 1019–1026. [Google Scholar] [CrossRef]
- Abzug, M.J. Presentation, diagnosis, and management of enterovirus infections in neonates. Pediatr. Drugs 2004, 6, 1–10. [Google Scholar] [CrossRef]
- Satosar, A.; Ramirez, N.C.; Bartholomew, D.; Davis, J.; Nuovo, G.J. Histologic correlates of viral and bacterial infection of the placenta associated with severe morbidity and mortality in the newborn. Hum. Pathol. 2004, 35, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Jaïdane, H.; Halouani, A.; Jmii, H.; Elmastour, F.; Mokni, M.; Aouni, M. Coxsackievirus B4 vertical transmission in a murine model. Virol. J. 2017, 14, 16. [Google Scholar] [CrossRef] [Green Version]
- Geenen, V.; Hober, D. For Debate: Programing of the Autoimmune Diabetogenic Response in the Thymus during Fetal and Perinatal Life. Pediatr. Endocrinol. Rev. 2019, 17, 78–83. [Google Scholar] [CrossRef]
- Halouani, A.; Jmii, H.; Michaux, H.; Renard, C.; Martens, H.; Pirottin, D.; Mastouri, M.; Aouni, M.; Geenen, V.; Jaïdane, H. Housekeeping Gene Expression in the Fetal and Neonatal Murine Thymus Following Coxsackievirus B4 Infection. Genes 2020, 11, 279. [Google Scholar] [CrossRef] [Green Version]
- Geenen, V.; Trussart, C.; Michaux, H.; Halouani, A.; Jaïdane, H.; Collée, C.; Renard, C.; Daukandt, M.; Ledent, P.; Martens, H. The presentation of neuroendocrine self-peptides in the thymus: An essential event for individual life and vertebrate survival. Ann. N. Y. Acad. Sci. 2019, 1455, 113–125. [Google Scholar] [CrossRef]
- Hansenne, I.; Renard-Charlet, C.; Greimers, R.; Geenen, V. Dendritic cell differentiation and immune tolerance to insulin-related peptides in Igf2-deficient mice. J. Immunol. 2006, 176, 4651–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, X.R.; Yang, G.; Li, M.; Song, J.P.; Liu, Z.Q.; Qiu, S.; Liu, Z.; Yang, P.C. Insulin-like growth factor-2 enhances functions of antigen (Ag)-specific regulatory B cells. J. Biol. Chem. 2014, 289, 17941–17950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Geng, X.R.; Song, J.P.; Wu, Y.; Yan, H.; Zhan, Z.; Yang, L.; He, W.; Liu, Z.Q.; Qiu, S.; et al. Insulin-like growth factor 2 enhances regulatory T-cell functions and suppresses food allergy in an experimental model. J. Allergy Clin. Immunol. 2014, 133, 1702–1708. [Google Scholar] [CrossRef] [PubMed]
- Jaïdane, H.; Caloone, D.; Lobert, P.E.; Sane, F.; Dardenne, O.; Naquet, P.; Gharbi, J.; Aouni, M.; Geenen, V.; Hober, D. Persistent infection of thymic epithelial cells with coxsackievirus B4 results in decreased expression of type 2 insulin-like growth factor. J. Virol. 2012, 86, 11151–11162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kecha, O.; Brilot, F.; Martens, H.; Franchimont, N.; Renard, C.; Greimers, R.; Defresne, M.P.; Winkler, R.; Geenen, V. Involvement of insulin-like growth factors in early T cell development: A study using fetal thymic organ cultures. Endocrinology 2000, 141, 1209–1217. [Google Scholar] [CrossRef]
- Michaux, H.; Martens, H.; Jaïdane, H.; Halouani, A.; Hober, D.; Geenen, V. How Does Thymus Infection by Coxsackievirus Contribute to the Pathogenesis of Type 1 Diabetes? Front. Immunol. 2015, 6, 338. [Google Scholar] [CrossRef] [Green Version]
- Michaux, H.; Halouani, A.; Trussart, C.; Renard, C.; Jaïdane, H.; Martens, H.; Geenen, V.; Hober, D. Modulation of IGF2 Expression in the Murine Thymus and Thymic Epithelial Cells Following Coxsackievirus B4 Infection. Microorganisms 2021, 9, 402. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhazmi, A.; Nekoua, M.P.; Michaux, H.; Sane, F.; Halouani, A.; Engelmann, I.; Alidjinou, E.K.; Martens, H.; Jaidane, H.; Geenen, V.; et al. Effect of Coxsackievirus B4 Infection on the Thymus: Elucidating Its Role in the Pathogenesis of Type 1 Diabetes. Microorganisms 2021, 9, 1177. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061177
Alhazmi A, Nekoua MP, Michaux H, Sane F, Halouani A, Engelmann I, Alidjinou EK, Martens H, Jaidane H, Geenen V, et al. Effect of Coxsackievirus B4 Infection on the Thymus: Elucidating Its Role in the Pathogenesis of Type 1 Diabetes. Microorganisms. 2021; 9(6):1177. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061177
Chicago/Turabian StyleAlhazmi, Abdulaziz, Magloire Pandoua Nekoua, Hélène Michaux, Famara Sane, Aymen Halouani, Ilka Engelmann, Enagnon Kazali Alidjinou, Henri Martens, Hela Jaidane, Vincent Geenen, and et al. 2021. "Effect of Coxsackievirus B4 Infection on the Thymus: Elucidating Its Role in the Pathogenesis of Type 1 Diabetes" Microorganisms 9, no. 6: 1177. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061177