
Characterization and Analysis of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPRs) in Pandemic and Non-Pandemic Vibrio parahaemolyticus Isolates from Seafood Sources




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Detection of Hemolysin Genes
2.3. Group-Specific PCR (GS-PCR)
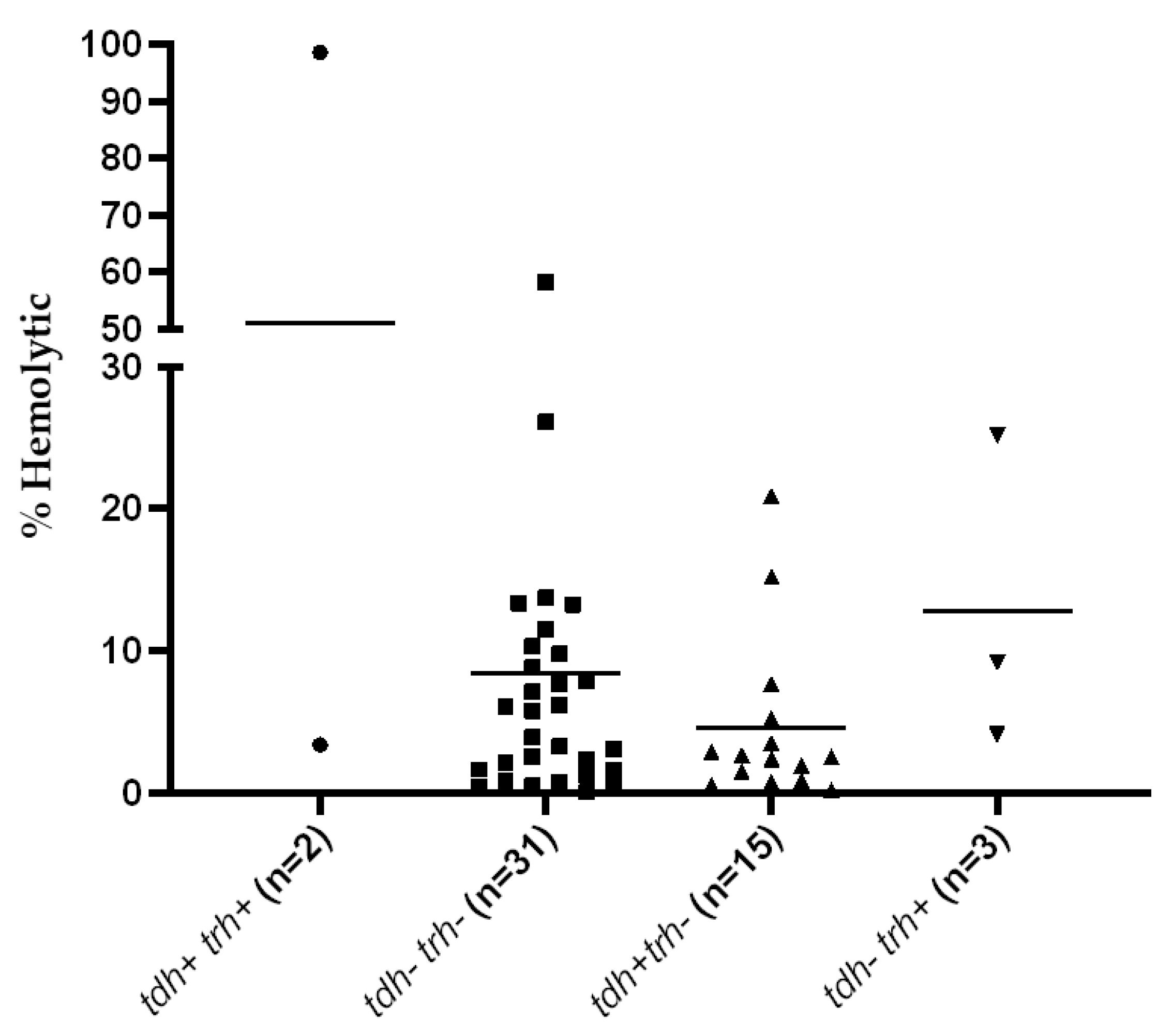
2.4. Determination of Hemolytic Activity
2.5. Kanagawa Phenomenon (KP) Assay
2.6. Antimicrobial Susceptibility Test
2.7. Determination of CRISPR Sequences
2.8. Analysis of CRISPR and CRISPR-Virulence Typing
2.9. Statistical Analyses
3. Results
3.1. Detection of Virulence Genes and Hemolytic Activity
3.2. Characterization of Antimicrobial Susceptibility
3.3. Analysis of CRISPR Repeat Sequences and Patterns
3.4. Spacer Sequence Analysis
3.5. Spacer Origin
3.6. CRISPR-Virulence Typing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelapati, S.; Nelapati, K.; Chinnam, B.K. Vibrio parahaemolyticus-An emerging foodborne pathogen—A Review. Vet. World 2012, 5, 48–62. [Google Scholar] [CrossRef]
- Broberg, C.A.; Calder, T.J.; Orth, K. Vibrio parahaemolyticus cell biology and pathogenicity determinants. Microbes Infect. 2011, 13, 992–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, C.; Okuda, J.; Ishibashi, M.; Iwanaga, M.; Garg, P.; Rammamurthy, T.; Wong, H.C.; Depaola, A.; Kim, Y.B.; Albert, M.J.; et al. Pandemic spread of an O3:K6 clone of Vibrio parahaemolyticus and emergence of related strains evidenced by arbitrarily primed PCR and toxRS sequence analyses. J. Clin. Microbiol. 2000, 38, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Meador-Kidd, C.; Parsons, M.; Bopp, C.; Gerner-Smidt, P.; Painter, J.; Vora, G. Virulence gene- and pandemic group-specific marker profiling of clinical Vibrio parahaemolyticus isolates. J. Clin. Microbiol. 2007, 45, 1133–1139. [Google Scholar] [CrossRef] [Green Version]
- Nishibuchi, M.; Kaper, J.B. Thermostable direct hemolysin gene of Vibrio parahaemolyticus: A virulence gene acquired by a marine bacterium. Infect. Immun. 1995, 63, 2093–2099. [Google Scholar] [CrossRef] [Green Version]
- Kishishita, M.; Matsuoka, N.; Kumagai, K.; Yamasaki, S.; Takeda, Y.; Nishibuchi, M. Sequence variation in the thermostable direct hemolysin-related hemolysin (trh) gene of Vibrio parahaemolyticus. Appl. Environ. Microbiol. 1992, 58, 2449–2457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terai, A.; Baba, K.; Shirai, H.; Yoshida, O.; Takeda, Y.; Nishibuchi, M. Evidence for insertion sequence-mediated spread of the thermostable direct hemolysin gene among Vibrio species. J. Bacteriol. 1991, 173, 5036–5046. [Google Scholar] [CrossRef] [Green Version]
- Espejo, R.T.; García, K.; Plaza, N. Insight Into the Origin and Evolution of the Vibrio parahaemolyticus Pandemic Strain. Front. Microbiol. 2017, 8, 1397. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Kato, T.; Obara, Y.; Akiyama, S.; Takizawa, K.; Yamai, S. In vitro hemolytic characteristic of Vibrio parahaemolyticus: Its close correlation with human pathogenicity. J. Bacteriol. 1969, 100, 1147–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igbinosa, E.O. Detection and antimicrobial resistance of Vibrio isolates in aquaculture environments: Implications for public health. Microb. Drug Resist. 2016, 22, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.; Embden, J.D.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Fabre, L.; Zhang, J.; Guigon, G.; Le Hello, S.; Guibert, V.; Accou-Demartin, M.; de Romans, S.; Lim, C.; Roux, C.; Passet, V.; et al. CRISPR typing and subtyping for improved laboratory surveillance of Salmonella infections. PLoS ONE 2012, 7, e36995. [Google Scholar]
- Dion, M.B.; Labrie, S.J.; Shah, S.A.; Moineau, S. CRISPRStudio: A User-Friendly software for rapid CRISPR array visualization. Viruses 2018, 10, 602. [Google Scholar] [CrossRef] [Green Version]
- Bangpanwimon, K.; Sottisuporn, J.; Mittraparp-Arthorn, P.; Ueaphatthanaphanich, W.; Rattanasupar, A.; Pourcel, C.; Vuddhakul, V. CRISPR-like sequences in Helicobacter pylori and application in genotyping. Gut Pathog. 2017, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Kongrueng, J.; Srinitiwarawong, K.; Nishibuchi, M.; Mittraparp-Arthorn, P.; Vuddhakul, V. Characterization and CRISPR-based genotyping of clinical trh-positive Vibrio parahaemolyticus. Gut Pathog. 2018, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Baliga, P.; Shekar, M.; Venugopal, M. Investigation of direct repeats, spacers and proteins associated with clustered regularly interspaced short palindromic repeat (CRISPR) system of Vibrio parahaemolyticus. Mol. Genet. Genom. 2019, 294, 253–262. [Google Scholar] [CrossRef]
- Sun, H.; Li, Y.; Shi, X.; Lin, Y.; Qiu, Y.; Zhang, J.; Liu, Y.; Jiang, M.; Zhang, Z.; Chen, Q.; et al. Association of CRISPR/Cas evolution with Vibrio parahaemolyticus virulence factors and genotypes. Foodborne Pathog. Dis. 2015, 12, 68–73. [Google Scholar] [CrossRef]
- Kim, Y.B.; Okuda, J.; Matsumoto, C.; Takahashi, N.; Hashimoto, S.; Nishibuchi, M. Identification of Vibrio parahaemolyticus strains at the species level by PCR targeted to the toxR gene. J. Clin. Microbiol. 1999, 37, 1173–1177. [Google Scholar] [CrossRef] [Green Version]
- Wootipoom, N.; Bhoopong, P.; Pomwised, R.; Nishibuchi, M.; Ishibashi, M.; Vuddhakul, V. A decrease in the proportion of infections by pandemic Vibrio parahaemolyticus in Hat Yai Hospital, southern Thailand. J. Med. Microbiol. 2007, 56, 1630–1638. [Google Scholar] [CrossRef]
- Tada, J.; Ohashi, T.; Nishimura, N.; Shirasaki, Y.; Ozaki, H.; Fukushima, S.; Takano, J.; Nishibuchi, M.; Takeda, Y. Detection of the thermostable direct hemolysin gene (tdh) and the thermostable direct hemolysin-related hemolysin gene (trh) of Vibrio parahaemolyticus by polymerase chain reaction. Mol. Cell. Probes 1992, 6, 477–487. [Google Scholar] [CrossRef]
- Rattanama, P.; Thompson, J.R.; Kongkerd, N.; Srinitiwarawong, K.; Vuddhakul, V.; Mekalanos, J.J. Sigma E regulators control hemolytic activity and virulence in a shrimp pathogenic Vibrio harveyi. PLoS ONE 2012, 7, e32523. [Google Scholar] [CrossRef]
- De Souza Santos, M.; Salomon, D.; Li, P.; Krachler, A.-M.; Orth, K. 8—Vibrio parahaemolyticus virulence determinants. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 4th ed.; Joseph, E., Alouf, J., Ladant, D., Popoff, M., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2015; pp. 230–260. [Google Scholar]
- The Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing CLSI Supplement M100S, 26th ed.; The Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016; pp. 74–75. [Google Scholar]
- Tan, C.W.; Rukayadi, Y.; Hasan, H.; Thung, T.Y.; Lee, E.; Rollon, W.D.; Hara, H.; Kayali, A.Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance patterns of Vibrio parahaemolyticus isolated from different types of seafood in Selangor, Malaysia. Saudi J. Biol. Sci. 2020, 27, 1602–1608. [Google Scholar] [CrossRef]
- Hofacker, I.L.; Fontana, W.; Stadler, P.F.; Bonhoeffer, L.S.; Tacker, M.; Schuster, P. Fast folding and comparison of RNA secondary structures. Mon. Chem. Chem. Mon. 1994, 125, 167–188. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibani, C.M.; Roth, O.; Liesegang, H.; Wendling, C.C. Genomic variation among closely related Vibrio alginolyticus strains is located on mobile genetic elements. BMC Genom. 2020, 21, 354. [Google Scholar] [CrossRef]
- Robert-Pillot, A.; Guénolé, A.; Lesne, J.; Delesmont, R.; Fournier, J.M.; Quilici, M.L. Occurrence of the tdh and trh genes in Vibrio parahaemolyticus isolates from waters and raw shellfish collected in two French coastal areas and from seafood imported into France. Int. J. Food Microbiol. 2004, 91, 319–325. [Google Scholar] [CrossRef]
- Mahoney, J.C.; Gerding, M.J.; Jones, S.H.; Whistler, C.A. Comparison of the pathogenic potentials of environmental and clinical Vibrio parahaemolyticus strains indicates a role for temperature regulation in virulence. Appl. Environ. Microbiol. 2010, 76, 7459–7465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Ueda, Y.; Furukawa, T.; Takegaki, Y.; Miyagi, K.; Noda, K.; Hirose, H.; Hashimoto, S.; Yano, S.; Ishibashi, M.; et al. Incidence of Kanagawa phenomenon-positive and -negative Vibrio parahaemolyticus strains isolated from traveller’s diarrhea and their relation to tdh and trh genes. Kansenshogaku Zasshi 1997, 71, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Lozano-León, A.; Torres, J.; Osorio, C.R.; Martínez-Urtaza, J. Identification of tdh-positive Vibrio parahaemolyticus from an outbreak associated with raw oyster consumption in Spain. FEMS Microbiol. Lett. 2003, 226, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Nishibuchi, M.; Kaper, J.B. Duplication and variation of the thermostable direct haemolysin (tdh) gene in Vibrio parahaemolyticus. Mol. Microbiol. 1990, 4, 87–99. [Google Scholar] [CrossRef]
- Okuda, J.; Nishibuchi, M. Manifestation of the Kanagawa phenomenon, the virulence-associated phenotype, of Vibrio parahaemolyticus depends on a particular single base change in the promoter of the thermostable direct haemolysin gene. Mol. Microbiol. 1998, 30, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kümmerer, K. Antibiotics in the aquatic environment—A review—Part I. Chemosphere 2009, 75, 417–434. [Google Scholar] [CrossRef]
- Han, F.; Walker, R.D.; Janes, M.E.; Prinyawiwatkul, W.; Ge, B. Antimicrobial Susceptibilities of Vibrio parahaemolyticus and Vibrio vulnificus Isolates from Louisiana Gulf and Retail Raw Oysters. Appl. Environ. Microbiol. 2007, 73, 7096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Wu, Q.; Zhang, J.; Xu, X.; Cheng, J. Comparison of Vibrio parahaemolyticus isolates from aquatic products and clinical by antibiotic susceptibility, virulence, and molecular characterisation. Food Control 2017, 71, 315–321. [Google Scholar] [CrossRef]
- Okoh, A.I.; Igbinosa, E.O. Antibiotic susceptibility profiles of some Vibrio strains isolated from wastewater final effluents in a rural community of the Eastern Cape Province of South Africa. BMC Microbiol. 2010, 10, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preeprem, S.; Bhoopong, P.; Srinitiwarawong, K.; Vuddhakul, V.; Mittraparp-arthorn, P. Antibiogram profiles and virulence characteristics of pandemic Vibrio parahaemolyticus isolates from diarrheal patients in Hat Yai hospital, Southern Thailand. Southeast Asian J. Trop. Med. Public Health 2019, 50, 132–145. [Google Scholar]
- Basak, S.; Singh, P.; Rajurkar, M. Multidrug Resistant and Extensively Drug Resistant Bacteria: A Study. J. Pathog. 2016, 2016, 4065603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baliga, P.; Shekar, M.; Ahamed, S.; Venugopal, M. Antibiotic resistance pattern and its correlation to the presence of tdh gene and CRISPR-Cas system in Vibrio parahaemolyticus strains isolated from seafood. Indian J. Fish. 2019, 66, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Elexson, N.; Afsah-Hejri, L.; Rukayadi, Y.; Soopna, P.; Lee, H.Y.; Zainazor, T.T.; Ainy, M.N.; Nakaguchi, Y.; Mitsuaki, N.; Son, R. Effect of detergents as antibacterial agents on biofilm of antibiotics-resistant Vibrio parahaemolyticus isolates. Food Control 2014, 35, 378–385. [Google Scholar] [CrossRef]
- Chao, G.; Jiao, X.; Zhou, X.; Wang, F.; Yang, Z.; Huang, J.; Pan, Z.; Zhou, L.; Qian, X. Distribution of genes encoding four pathogenicity islands (VPaIs), T6SS, biofilm, and type I pilus in food and clinical strains of Vibrio parahaemolyticus in China. Foodborne Pathog. Dis. 2010, 7, 649–658. [Google Scholar] [CrossRef]
- Hurley, C.C.; Quirke, A.; Reen, F.J.; Boyd, E.F. Four genomic islands that mark post-1995 pandemic Vibrio parahaemolyticus isolates. BMC Genom. 2006, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of V. cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- Horvath, P.; Romero, D.A.; Coûté-Monvoisin, A.C.; Richards, M.; Deveau, H.; Moineau, S.; Boyaval, P.; Fremaux, C.; Barrangou, R. Diversity, Activity, and Evolution of CRISPR Loci in Streptococcus thermophilus. J. Bacteriol. 2008, 190, 1401. [Google Scholar] [CrossRef] [Green Version]
- McGinn, J.; Marraffini, L.A. CRISPR-Cas Systems Optimize Their Immune Response by Specifying the Site of Spacer Integration. Mol. Cell 2016, 64, 616–623. [Google Scholar] [CrossRef]
- Li, W.; Bian, X.; Evivie, S.E.; Huo, G.C. Comparative Analysis of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) of Streptococcus thermophilus St-I and its Bacteriophage-Insensitive Mutants (BIM) Derivatives. Curr. Microbiol. 2016, 73, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Virolle, C.; Goldlust, K.; Djermoun, S.; Bigot, S.; Lesterlin, C. Plasmid transfer by conjugation in gram-negative bacteria: From the cellular to the community level. Genes 2020, 11, 1239. [Google Scholar] [CrossRef] [PubMed]
- Nasko, D.J.; Ferrell, B.D.; Moore, R.M.; Bhavsar, J.D.; Polson, S.W.; Wommack, K.E. CRISPR spacers indicate preferential matching of specific virioplankton genes. mBio 2019, 10, e02651-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasu, H.; Iida, T.; Sugahara, T.; Yamaichi, Y.; Park, K.S.; Yokoyama, K.; Makino, K.; Shinagawa, H.; Honda, T. A filamentous phage associated with recent pandemic Vibrio parahaemolyticus O3:K6 strains. J. Clin. Microbiol. 2000, 38, 2156–2161. [Google Scholar] [CrossRef]
- Iida, T.; Hattori, A.; Tagomori, K.; Nasu, H.; Naim, R.; Honda, T. Filamentous phage associated with recent pandemic strains of Vibrio parahaemolyticus. Emerg. Infect. Dis. 2001, 7, 477–478. [Google Scholar] [CrossRef] [PubMed]
- Castillo, D.; Kauffman, K.; Hussain, F.; Kalatzis, P.; Rørbo, N.; Polz, M.F.; Middelboe, M. Widespread distribution of prophage-encoded virulence factors in marine Vibrio communities. Nature 2018, 8, 9973. [Google Scholar] [CrossRef] [PubMed]
- Sorek, R.; Kunin, V.; Hugenholtz, P. CRISPR-a widespread system that provides acquired resistance against phages in bacteria and archaea. Nature 2008, 6, 181–186. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hemolysin Genotype (n) | Isolates | Year of Isolation | Source | GS-PCR * | Kanagawa Phenomenon | MAR * Index | CRISPR-PCR |
|---|---|---|---|---|---|---|---|
| Seafood isolates | |||||||
| tdh+ trh− (15) | PSU166 | 2000 | Hard clam | + | + | 0.2 | + |
| PSU358 | 2001 | Mussel Mussel Mussel Mussel Hard clam | + | + | 0.2 | + | |
| PSU360 | + | + | 0.2 | + | |||
| PSU434 | + | + | 0.2 | + | |||
| PSU474 | + | + | 0.2 | + | |||
| PSU476 | + | + | 0.1 | + | |||
| PSU479 | 2002 | Hard clam | + | + | 0.1 | + | |
| PSU579 | Cockle | + | + | 0.2 | + | ||
| PSU635 | Mussel | + | + | 0.3 | + | ||
| PSU637 | Mussel | + | + | 0.2 | + | ||
| PSU638 | Mussel | + | + | 0.2 | + | ||
| PSU3249 | 2006 | Mussel | + | − | 0.1 | + | |
| PSU4067 | 2008 | Cockle | + | + | 0.1 | + | |
| PSU4888 | 2010 | Hard clam | + | + | 0.1 | + | |
| PSU5382 | 2012 | Shellfish | + | + | 0.1 | + | |
| tdh− trh+ (3) | PSU3819 PSU3831 | 2007 | Crab Fish | − | − | 0.2 | + |
| − | − | 0.2 | + | ||||
| PSU5124 | 2011 | Shrimp | − | − | 0.1 | − | |
| tdh+ trh+ (2) | PSU582 | 2002 | Cockle | − | + | 0.2 | + |
| PSU4413 | 2008 | Cockle | − | − | 0.1 | − | |
| tdh− trh− (31) | PSU513 PSU571 PSU576 PSU578 | 2002 | Cockle Hard clam Cockle Cockle | − | − | 0.1 0.1 | − |
| − | − | 0.1 | − | ||||
| − | − | 0.1 | + | ||||
| PSU811 | 2003 | Mussel | − | − | 0.1 | − | |
| PSU2463 PSU2467 PSU2471 | 2005 | Cockle Cockle Cockle | − | − | 0.2 | − | |
| − | − | 0.2 | − | ||||
| − | − | 0.2 | − | ||||
| PSU3103 PSU3200 PSU3362 PSU3365 | 2006 | Hard clam Cockle Mussel Cockle | − | − | 0.1 0.2 0.2 0.1 | − | |
| PSU3858 PSU4055 PSU4058 PSU4062 PSU4075 PSU4091 PSU4094 | 2008 | Octopus Mussel Hard clam Cockle Hard clam Cockle Cockle | − | − | 0.4 | − | |
| − | − | 0.1 | − | ||||
| − | − | 0.1 | − | ||||
| − | − | 0.2 | − | ||||
| − | − | 0.1 | − | ||||
| − | − | 0.1 | − | ||||
| − | − | 0.1 | − | ||||
| PSU4415 PSU4418 PSU4425 PSU4446 PSU4459 PSU4460 PSU4575 | 2009 | Cockle Cockle Cockle Cockle Cockle Cockle Cockle | − | − | 0.1 | − | |
| − | − | 0.1 | − | ||||
| − | − | 0.1 | − | ||||
| − | − | 0.4 | − | ||||
| − | − | 0.1 | − | ||||
| − | − | 0.1 | − | ||||
| − | − | 0.1 | − | ||||
| PSU4869 PSU4879 PSU4885 PSU4895 | 2010 | Mussel Mussel Hard clam Hard clam | − | − | 0.2 | − | |
| − | − | 0.2 | − | ||||
| − | − | 0.1 | − | ||||
| − | − | 0.2 | − | ||||
| PSU5379 | 2012 | Shellfish | − | − | 0.1 | − | |
| Clinical isolates | |||||||
| tdh + trh− (3) | PSU3872 | 2008 | Clinical | + | + | 0.3 | + |
| PSU3949 | 2008 | Clinical | + | + | 0.2 | + | |
| PSU5126 | 2011 | Clinical | + | + | 0.1 | + | |
| tdh− trh+ (1) | ATCC17802 | 1965 | Clinical | − | − | 0.2 | − |
| Antimicrobial Drugs | No. of Isolates (%) | ||
|---|---|---|---|
| Susceptible | Intermediate | Resistant | |
| Ampicillin | 0 | 0 | 51 (100) |
| Gentamycin | 12 (23.5) | 39 (76.5) | 0 |
| Erythromycin | 0 | 29 (56.9) | 22 (43.1) |
| Sulfonamide | 49 (96) | 1 (2) | 1 (2) |
| Tetracycline | 49 (96) | 1 (2) | 1 (2) |
| Ciprofloxacin | 12 (23.5) | 38 (74.5) | 1 (2) |
| Trimethoprim/sulfamethoxazole | 36 (70.6) | 13 (25.5) | 2 (3.9) |
| Chloramphenicol | 51 (100) | 0 | 0 |
| Norfloxacin | 34 (66.7) | 16 (31.3) | 1 (2) |
| Hemolysin Genotype (n) | Type | Direct Repeats (DRs) Sequences * | No. of DRs | No. of Spacers (Sp Type) | CRISPR Locus Pattern (bp) * |
|---|---|---|---|---|---|
| Seafood isolates | |||||
| tdh+ trh− (15) | Typical repeat Terminal repeat | GTGAACTGCCGAATAGGTAGCTGATAAT GTGAACTGCCGCATAGGTAGAGAGAATC | 3 | 2 (1, 4) | 28-32-28-31-28 |
| tdh− trh+ (2) | Typical repeat Terminal repeat | GTGAACTGCCGAATAGGTAGCTGATAAT GTGAACTGCCGCATAGGTAGAGAGGATC | 3 | 2 (2, 4) | 28-31-28-31-28 |
| tdh+ trh+ (1) | Typical repeat Terminal repeat | GTGAACTGCCGAATAGGTAGCTGATAAT GTGAACTGCCGCATAGGTAGAGAGGATC | 2 | 1 (3) | 28-31-28 |
| tdh− trh− (1) | Typical repeat Terminal repeat | GTGAACTGCCGAATAGGTAGCTGATAAT GTGAACTGCCGAATAGGTAGAGAGGATC | 2 | 1 (1) | 28-32-28 |
| Clinical isolates | |||||
| tdh + trh− (3) | Typical repeat Terminal repeat | GTGAACTGCCGAATAGGTAGCTGATAAT GTGAACTGCCGCATAGGTAGAGAGAATC | 3 | 2 (1, 4) | 28-32-28-31-28 |
| Spacer Type (Length) | Spacer Sequences (5′ to 3′) | Spacer Homology | Query Cover/ Identities (%) | Accession Number/ Location |
|---|---|---|---|---|
| Sp1 (32) | GAGATACCACAAGCTCAAGCAGATGCTAACAG | Vibrio alginolyticus strain K08M3 plasmid pL300 | 93/96.7 | CP017915.1/ 184601-184630 |
| Sp2 (31) | TCATTCTCACGATCTAATTACAGTTGGTCAC | Vibrio alginolyticus strain K08M3 plasmid pL300 | 93/100 | CP017915.1/ 155899-155927 |
| Sp3 (31) | TGCAGACAAACAAAGAGCCATCGACGAGTGC | Vibrio alginolyticus strain K08M3 plasmid pL300 | 100/87.1 | CP017915.1/ 141985-142015 |
| Sp4 (31) | AGTCGGTCAACTGAGAATACGTTGTTGCCAA | - | - | - |
| Spacer Type | Isolates | Hemolysin Genotype * | Sources * | Identities (%) | E-Value | Accession Number |
|---|---|---|---|---|---|---|
| Sp1 | Vibrio parahaemolyticus strain RIMD 2210633 | tdh+ trh− | Clinical, Japan | 100 | 2 × 10−8 | BA000032.2 |
| Vibrio parahaemolyticus strain VPD14 | tdh+ trh− | Shrimp, China | 100 | 2 × 10−8 | CP031782.1 | |
| Vibrio parahaemolyticus strain FDAARGOS_191 | tdh+ trh− | Clinical, India | 100 | 2 × 10−8 | CP020428.2 | |
| Vibrio parahaemolyticus strain BB220P | tdh+ trh− | Environment, India | 96.88 | 1 × 10−6 | CP003973.1 | |
| Vibrio parahaemolyticus strain FORC_071 | tdh+ trh− | Clinical, South Korea | 100 | 1 × 10−6 | CP023486.1 | |
| Sp2 | Vibrio parahaemolyticus strain Vp17 | tdhunk trhunk | Clam, India | 100 | 7 × 10−8 | MG765521.1 |
| Vibrio parahaemolyticus strain Vp14 | tdhunk trhunk | Oyster, India | 100 | 7 × 10−8 | MG765520.1 | |
| Vibrio parahaemolyticus strain Vp8 | tdhunk trhunk | Shrimp, India | 100 | 9 × 10−7 | MG765517.1 | |
| Sp3 | Vibrio parahaemolyticus strain 10329 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP045795.1 |
| Vibrio parahaemolyticus strain MAVP-26 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP023247.1 | |
| Vibrio parahaemolyticus strain ST631 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP011885.1 | |
| Vibrio parahaemolyticus strain MAVP-QPI | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | MF066646.1 | |
| Vibrio parahaemolyticus strain MAVP-Q | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP022472.1 | |
| Vibrio parahaemolyticus strain FDAARGOS_662 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP044070.1 | |
| Vibrio parahaemolyticus strain FDAARGOS_51 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP026042.1 | |
| Vibrio parahaemolyticus strain 2014V-1125 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP046777.1 | |
| Vibrio parahaemolyticus strain 2014V-1066 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP046780.1 | |
| Vibrio parahaemolyticus strain 2015AW-0174 | tdh+ trh+ | Clinical, USA | 100 | 7 × 10−8 | CP046753.1 | |
| Vibrio parahaemolyticus strain 2010V-1106 | tdhunk trh+ | Clinical, USA | 100 | 7 × 10−8 | CP046827.1 | |
| Vibrio parahaemolyticus strain 2013V-1146 | tdhunk trh+ | Clinical, USA | 100 | 7 × 10−8 | CP046809.1 | |
| Vibrio parahaemolyticus strain 2013V-1181 | tdhunk trh+ | Clinical, USA | 100 | 7 × 10−8 | CP046784.1 | |
| Sp4 | Vibrio parahaemolyticus strain RIMD 2210633 | tdh+ trh− | Clinical, Japan | 100 | 7 × 10−8 | BA000032.2 |
| Vibrio parahaemolyticus strain VPD14 | tdh+ trh− | Shrimp, China | 100 | 7 × 10−8 | CP031782.1 | |
| Vibrio parahaemolyticus strain FDAARGOS_191 | tdh+ trh− | Clinical, India | 100 | 7 × 10−8 | CP020428.2 | |
| Vibrio parahaemolyticus strain BB22OP | tdh+ trh− | Environment, Bangladesh | 100 | 7 × 10−8 | CP003973.1 | |
| Vibrio parahaemolyticus strain FORC_071 | tdh+ trh− | Clinical, South Korea | 100 | 7 × 10−8 | CP023486.1 | |
| Vibrio parahaemolyticus strain Vp14 | tdhunk trhunk | Oyster, India | 100 | 7 × 10−8 | MG765520.1 | |
| Vibrio parahaemolyticus strain Vp9 | tdhunk trhunk | Oyster, India | 100 | 7 × 10−8 | MG765518.1 | |
| Vibrio parahaemolyticus strain Vp8 | tdhunk trhunk | Shrimp, India | 100 | 7 × 10−8 | MG765517.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jingjit, N.; Preeprem, S.; Surachat, K.; Mittraparp-arthorn, P. Characterization and Analysis of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPRs) in Pandemic and Non-Pandemic Vibrio parahaemolyticus Isolates from Seafood Sources. Microorganisms 2021, 9, 1220. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061220
Jingjit N, Preeprem S, Surachat K, Mittraparp-arthorn P. Characterization and Analysis of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPRs) in Pandemic and Non-Pandemic Vibrio parahaemolyticus Isolates from Seafood Sources. Microorganisms. 2021; 9(6):1220. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061220
Chicago/Turabian StyleJingjit, Nawaporn, Sutima Preeprem, Komwit Surachat, and Pimonsri Mittraparp-arthorn. 2021. "Characterization and Analysis of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPRs) in Pandemic and Non-Pandemic Vibrio parahaemolyticus Isolates from Seafood Sources" Microorganisms 9, no. 6: 1220. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061220