
Genetic Diversity of Leuconostoc mesenteroides Isolates from Traditional Montenegrin Brine Cheese

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin and Cultivation of Isolates
2.2. Biochemical Testing
2.2.1. Growth at Different Temperatures
2.2.2. Production of CO2
2.2.3. Salt Tolerance
2.2.4. Acidification and Post-Acidification Ability in Milk
2.2.5. Catalase Testing
2.2.6. Ability for Formation of Exopolysaccharides (EPS)
2.3. Antimicrobial Susceptibility Testing
2.4. DNA Extraction and Whole Genome Sequencing
2.5. Sequence Data Analysis
2.6. Gene Presence and Absence
2.7. Nucleotide Sequence Accession Numbers
3. Results
3.1. Biochemical Properties
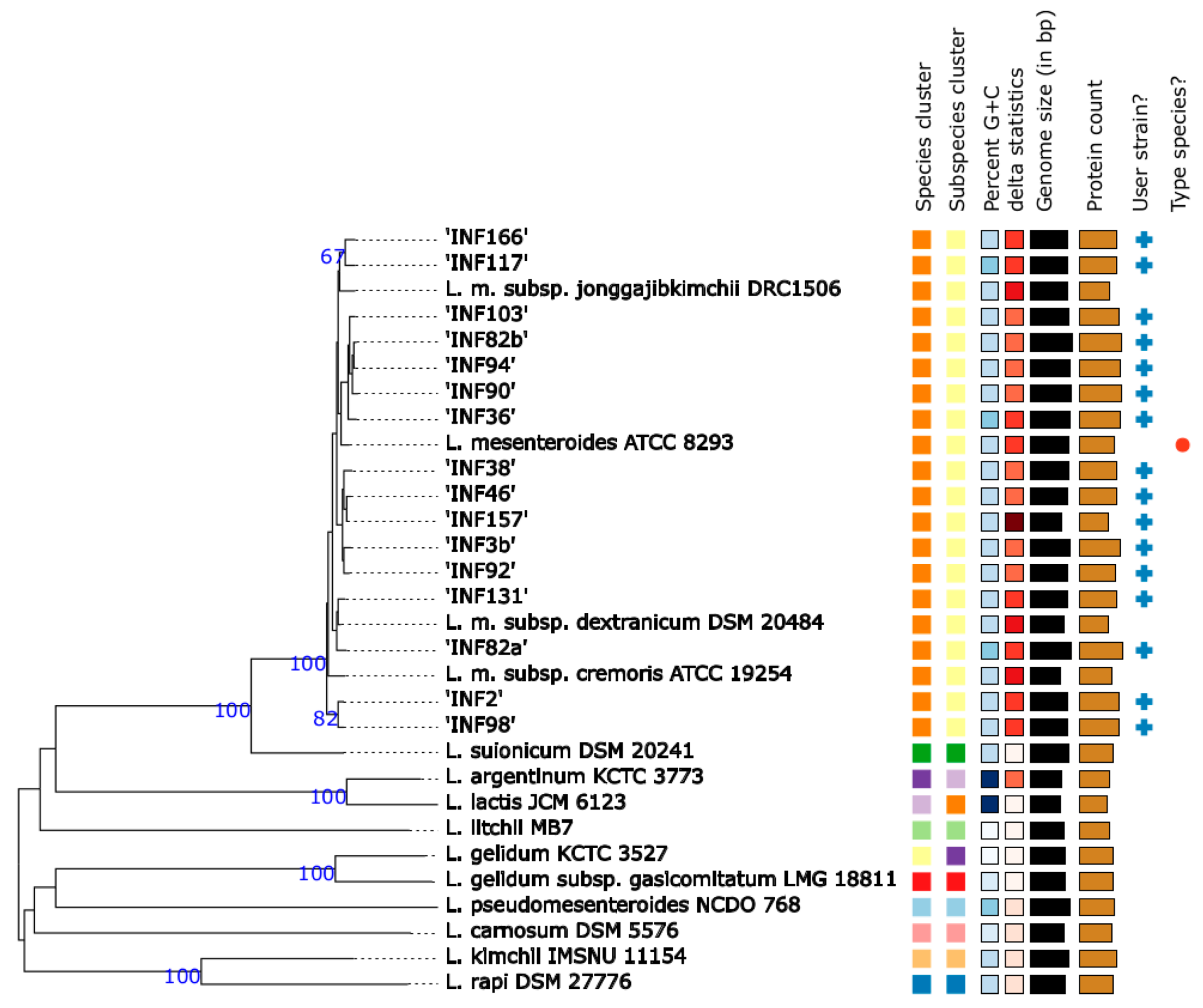
3.2. Species and Subspecies Identification
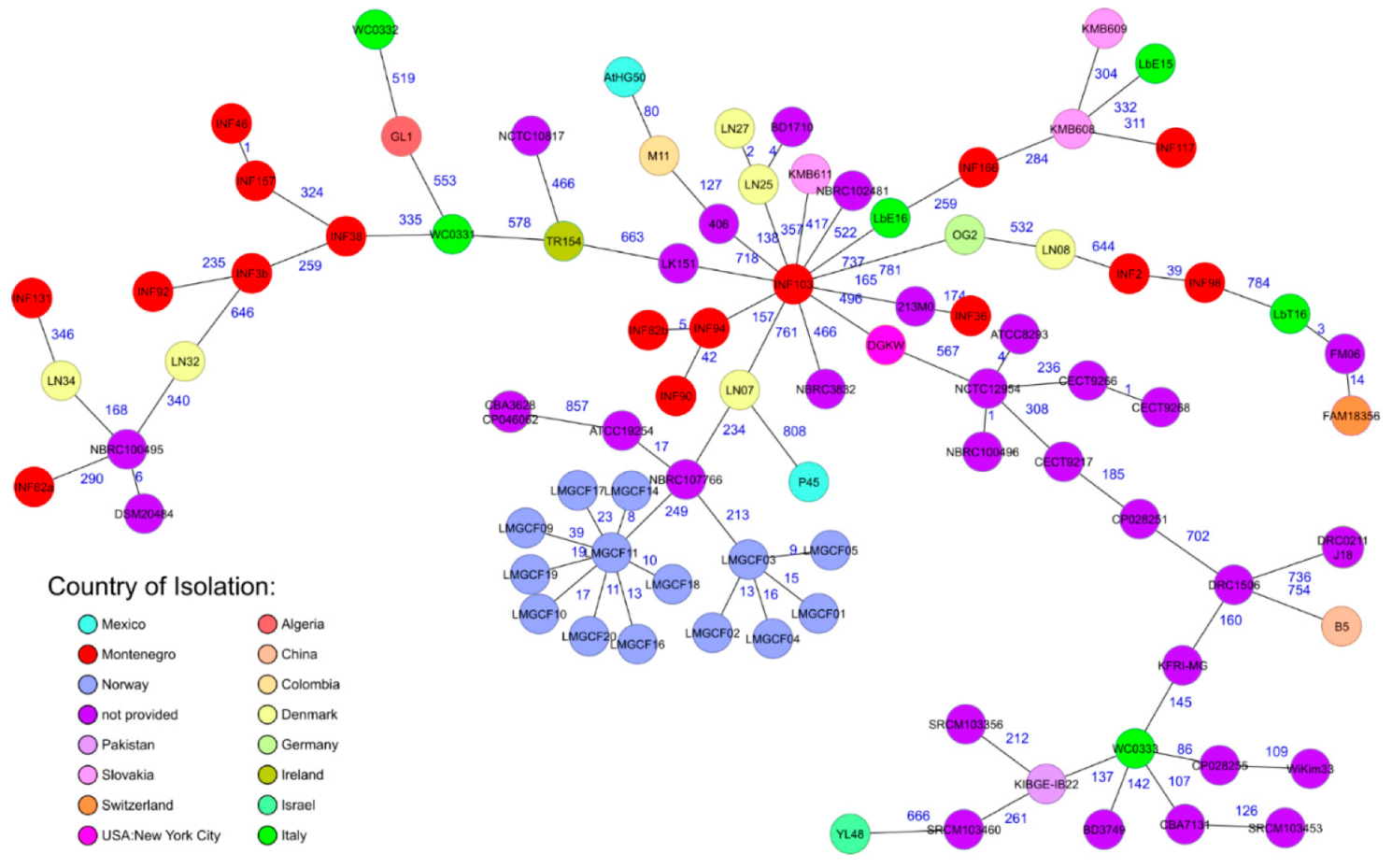
3.3. Whole Genome Sequence Based Subtyping
3.4. Detection of Bacteriocin and Secondary Metabolite Genes
3.5. Safety Evaluation
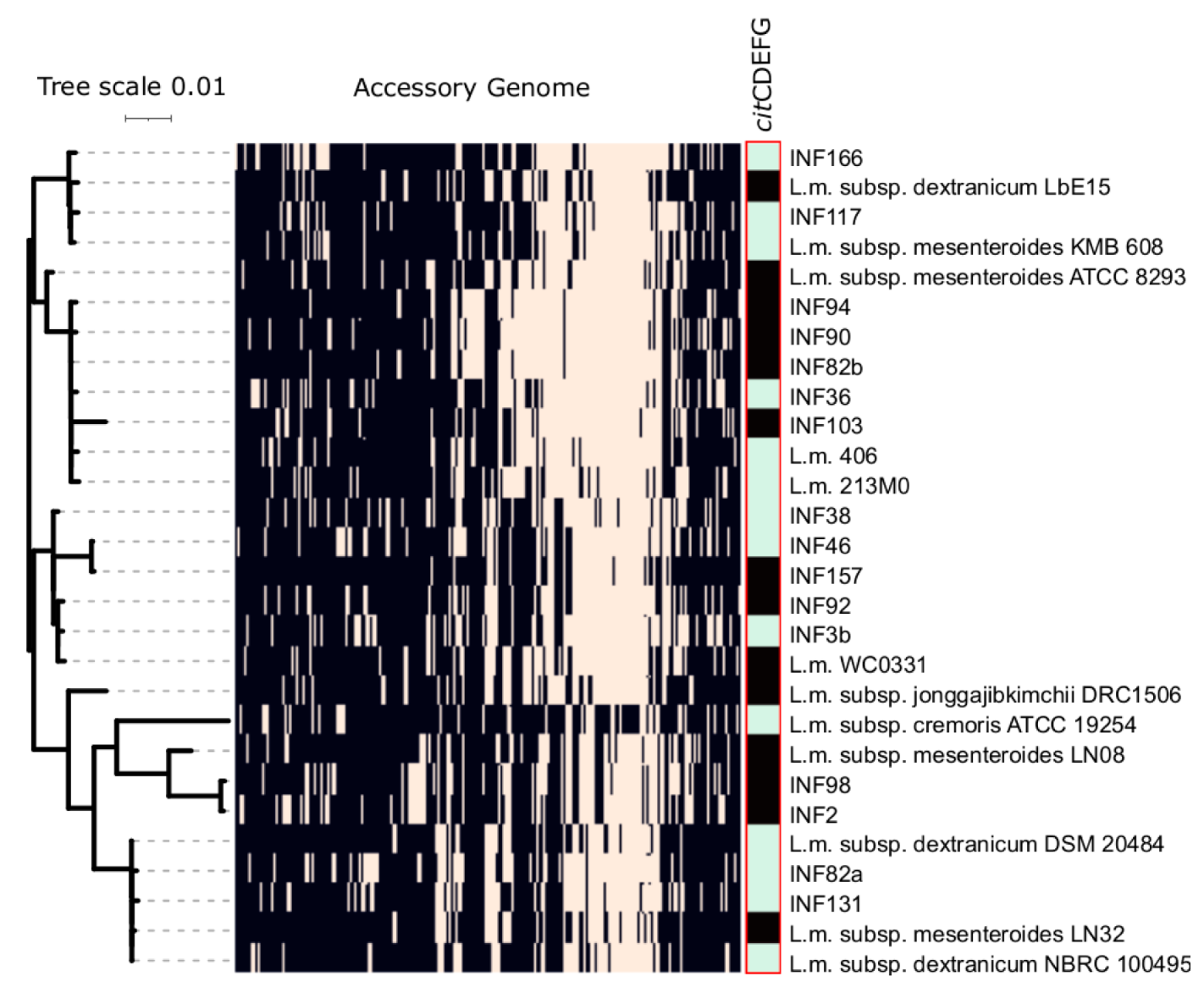
3.6. Orthofinder Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cogan, T.M.; Jordan, K.N. Metabolism of Leuconostoc bacteria. J. Dairy Sci. 1994, 77, 2704–2717. [Google Scholar] [CrossRef]
- Thunell, R.K. Taxonomy of the Leuconostocs. J. Dairy Sci. 1995, 78, 2514–2522. [Google Scholar] [CrossRef]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [Google Scholar] [CrossRef]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Nat. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [Green Version]
- Garvie, E.I. Proposal of neotype strains for Leuconostoc mesenteroides (Tsenkovskii) van Tieghem, Leuconostoc dextranicum (Beijerinck) Hucker and Pederson, and Leuconostoc cremoris (Knudsen and Sørensen) Garvie. Int. J. Syst. Evol. Microbiol. 1979, 29, 149–151. [Google Scholar]
- Garvie, E.I. NOTES: Leuconostoc mesenteroides subsp. cremoris (Knudsen and Sørensen) comb. nov. and Leuconostoc mesenteroides subsp. dextranicum (Beijerinck) comb. nov. Int. J. Syst. Evol. Microbiol. 1983, 33, 118–119. [Google Scholar] [CrossRef]
- Van Mastrigt, O.; Abee, T.; Smid, E.J. Complete Genome Sequences of Lactococcus lactis subsp. lactis bv. diacetylactis FM03 and Leuconostoc mesenteroides FM06 Isolated from Cheese. Genome Announc. 2017, 5, e00633-e17. [Google Scholar]
- Özcan, E.; Selvi, S.S.; Nikerel, E.; Teusink, B.; Öner, E.T.; Çakır, T. A genome-scale metabolic network of the aroma bacterium Leuconostoc mesenteroides subsp. cremoris. Appl. Microbiol. Biotech. 2019, 103, 3153–3165. [Google Scholar] [CrossRef] [PubMed]
- Starrenburg, M.J.; Hugenholtz, J. Citrate Fermentation by Lactococcus and Leuconostoc spp. Appl. Environm. Microbiol. 1991, 57, 3535–3540. [Google Scholar] [CrossRef] [Green Version]
- De Paula, A.T.; Jeronymo-Ceneviva, A.B.; Todorov, S.D.; Penna, A.L.B. The two faces of Leuconostoc mesenteroides in food systems. Food Rev. Int. 2015, 31, 147–171. [Google Scholar] [CrossRef]
- Stiles, M.E. Bacteriocins produced by Leuconostoc species. J. Dairy Sci. 1994, 77, 2718–2724. [Google Scholar] [CrossRef]
- Arqués, J.L.; Rodríguez, E.; Langa, S.; Landete, J.M.; Medina, M. Antimicrobial activity of lactic acid bacteria in dairy products and gut: Effect on pathogens. BioMed Res. Int. 2015, 584183. [Google Scholar] [CrossRef]
- Thangavel, G.; Subramaniyam, T. Antimicrobial Efficacy of Leuconostoc spp. Isolated from Indian Meat against Escherichia coli and Listeria monocytogenes in Spinach Leaves. Food Sci. Anim. Resour. 2019, 39, 677–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deegan, L.H.; Cotter, P.D.; Hill, C.; Ross, P. Bacteriocins: Biological tools for bio-preservation and shelf-life extension. Int. Dairy J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]
- Terzić-Vidojević, A.; Veljović, K.; Tolinački, M.; Živković, M.; Lukić, J.; Lozo, J.; Fira, Đ.; Jovčić, B.; Strahinić, I.; Begović, J.; et al. Diversity of non-starter lactic acid bacteria in autochthonous dairy products from Western Balkan Countries—Technological and probiotic properties. Food Res. Int. 2020, 136, 109494. [Google Scholar] [CrossRef]
- Guerrero, L.; Guàrdia, M.D.; Xicola, J.; Verbeke, W.; Vanhonacker, F.; Zakowska-Biemans, S.; Sajdakowska, M.; Sulmont-Rossé, C.; Issanchou, S.; Contel, M.; et al. Consumer-driven definition of traditional food products and innovation in traditional foods. A qualitative cross-cultural study. Appetite 2009, 52, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.F.; Kozak, I.M.; Flynn, S.; O’Gara, F. The Microbiome of an Active Meat Curing Brine. Front. Microbiol. 2019, 9, 3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardamone, L.; Quiberoni, A.; Mercanti, D.; Fornasari, M.; Reinheimer, J.; Guglielmotti, D.M. Adventitious dairy Leuconostoc strains with interesting technological and biological properties useful for adjunct starters. Dairy Sci. Technol. 2011, 91, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Chun, B.H.; Kim, K.H.; Jeon, H.H.; Lee, S.H.; Jeon, C.O. Pan-genomic and transcriptomic analyses of Leuconostoc mesenteroides provide insights into its genomic and metabolic features and roles in kimchi fermentation. Sci. Rep. 2017, 7, 11504. [Google Scholar] [CrossRef] [Green Version]
- Frantzen, C.A.; Kot, W.; Pedersen, T.B.; Ardö, Y.M.; Broadbent, J.R.; Neve, H.; Hansen, L.H.; Dal Bello, F.; Østlie, H.M.; Kleppen, H.P.; et al. Genomic Characterization of Dairy Associated Leuconostoc Species and Diversity of Leuconostocs in Undefined Mixed Mesophilic Starter Cultures. Front. Microbiol. 2017, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, V.K.; Koza, A.; Al-Nakeeb, K.; Oeregaard, G. Screening for texturing Leuconostoc and genomics behind polysaccharide production. FEMS Microbiol. Lett. 2020, 367, fnaa179. [Google Scholar] [CrossRef]
- Pedersen, T.B.; Kot, W.P.; Hansen, L.H.; Sørensen, S.J.; Broadbent, J.R.; Vogensen, F.K.; Ardö, Y. Genome Sequence of Leuconostoc mesenteroides subsp. cremoris Strain T26, Isolated from Mesophilic Undefined Cheese Starter. Genome Announc. 2014, 2, e00485-14. [Google Scholar]
- Zhang, W.; Liu, W.; Song, Y.; Xu, H.; Menghe, B.; Zhang, H.; Sun, Z. Multilocus sequence typing of a dairy-associated Leuconostoc mesenteroides population reveals clonal structure with intragenic homologous recombination. J. Dairy Sci. 2015, 98, 2284–2293. [Google Scholar] [CrossRef]
- Sharma, A.; Kaur, J.; Lee, S.; Park, Y.S. Analysis of Leuconostoc citreum strains using multilocus sequence typing. Food Sci. Biotechnol. 2018, 27, 1755–1760. [Google Scholar] [CrossRef]
- Chen, J.; Lv, H.; Zhang, Z.; Zhang, H.; Zhang, B.; Wang, X.; Liu, Y.; Zhang, M.; Pang, H.; Qin, G.; et al. Multilocus Sequence Typing of Leuconostoc mesenteroides Strains from the Qinghai-Tibet Plateau. Front. Microbiol. 2021, 12, 614286. [Google Scholar] [CrossRef]
- Campedelli, I.; Flórez, A.B.; Salvetti, E.; Delgado, S.; Orrù, L.; Cattivelli, L.; Alegría, Á.; Felis, G.E.; Torriani, S.; Mayo, B. Draft Genome Sequence of Three Antibiotic-Resistant Leuconostoc mesenteroides Strains of Dairy Origin. Genome Announc. 2015, 3, e01018-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, G.S.; Hong, S.J.; Jung, B.K.; Lee, C.; Park, C.K.; Shin, J.H. The complete genome sequence of a lactic acid bacterium Leuconostoc mesenteroides ssp. dextranicum strain DSM 20484(T). J. Biotech. 2016, 219, 3–4. [Google Scholar] [CrossRef]
- O’Donnell, S.T.; Ross, R.P.; Stanton, C. The Progress of Multi-Omics Technologies: Determining Function in Lactic Acid Bacteria Using a Systems Level Approach. Front. Microbiol. 2020, 28, 3084. [Google Scholar] [CrossRef] [PubMed]
- Franz, E.; Gras, L.M.; Dallman, T. Significance of whole genome sequencing for surveillance, source attribution and microbial risk assessment of foodborne pathogens. Curr. Opin. Food Sci. 2016. [Google Scholar] [CrossRef]
- Martinovic, A.; Narvhus, J.; Abrahamsen, R.K.; Ostlie, H.M.; Skeie, S.B. Application of indigenous strains of lactic acid bacteria for semi-industrial production of autochthonous Montenegrin Njeguši cheese. Int. J. Dairy Technol. 2018, 71, 1–10. [Google Scholar] [CrossRef]
- De Bruyne, K.; Slabbinck, B.; Waegeman, W.; Vauterin, P.; De Baets, B.; Vandamme, P. Bacterial species identification from MALDI-TOF mass spectra through data analysis and machine learning. Syst. Appl. Microbiol. 2011, 34, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Bojanic Rasovic, M.; Mayrhofer, S.; Martinovic, A.; Dürr, K.; Domig, K.J. Lactococci of Local Origin as Potential Starter Cultures for Traditional Montenegrin Cheese Production. Food Technol. Biotechnol. 2017, 55, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Veskovic-Moracanin, S.; Borovic, B.; Velebit, B. Morphological and biochemical characteristics of natural isolates of lactic acid bacteria isolated from Zlatar cheese. Tehnol. Mesa 2013, 54, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Martinović, A.; Radulović, Z.; Wind, A.; Janzen, T.; Obradović, D. Isolation and characterization of bacterial flora from farmhouse fermented milk products of Serbia and Montenegro. Acta Vet. 2005, 55, 307–318. [Google Scholar]
- European Food Safety Authority (EFSA) Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740–2750. [Google Scholar]
- Flórez, A.B.; Campedelli, I.; Delgado, S.; Alegría, Á.; Salvetti, E.; Felis, G.E.; Mayo, B.; Torriani, S. Antibiotic Susceptibility Profiles of Dairy Leuconostoc, Analysis of the Genetic Basis of Atypical Resistances and Transfer of Genes In Vitro and in a Food Matrix. PLoS ONE 2016, 11, e0145203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Ondov, B.D.; Treangen, T.J.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Fast genome and metagenome distance estimation using MinHash. bioRxiv 2015. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bliss, C.M.; Bennett, J.S.; Bratcher, H.B.; Brehony, C.; Colles, F.M.; Wimalarathna, H.; Harrison, O.B.; Sheppard, S.K.; Cody, A.J.; et al. Ribosomal multilocus sequence typing: Universal characterization of bacteria from domain to strain. Microbiology 2012, 158, 1005–1015. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Comm. 2018, 9, 5114. [Google Scholar] [CrossRef] [Green Version]
- Van Heel, A.J.; De Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly Web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kautsar, S.A.; Medema, M.H.; Weber, T. The antiSMASH database version 3: Increased taxonomic coverage and new query features for modular enzymes. Nucleic Acids Res. 2021, 49, D639–D643. [Google Scholar] [CrossRef]
- Cosentino, S.; Voldby Larsen, M.; Møller Aarestrup, F.; Lund, O. PathogenFinder—Distinguishing Friend from Foe Using Bacterial Whole Genome Sequence Data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Micobiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Martinović, A.; Mirecki, S. Food, nutrition, and health in Montenegro. In Nutritional and Health Aspects of Food in the Balkans; Lelieveld, H., Prakash, V., Prakash, J., Van der Meulen, B., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 165–185. [Google Scholar]
- Devirgiliis, C.; Zinno, P.; Perozzi, G. Update on antibiotic resistance in foodborne Lactobacillus and Lactococcus species. Front. Microbiol. 2013, 4, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. Scientific opinion of the panel on biological hazards on the request from EFSA on the maintenance of the list of QPS microorganisms intentionally added to food or feed. EFSA J. 2008, 928, 1–48. [Google Scholar]
- Menegueti, M.G.; Gaspar, G.G.; Laus, A.M.; Basile-Filho, A.; Bellissimo-Rodrigues, F.; Auxiliadora-Martins, M. Bacteremia by Leuconostoc mesenteroides in an immunocompetent patient with chronic Chagas disease: A case report. BMC Infect Dis. 2018, 18, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvetti, E.; Campedelli, I.; Larini, I.; Conedera, G.; Torriani, S. Exploring Antibiotic Resistance Diversity in Leuconostoc spp. by a Genome-Based Approach: Focus on the lsaA Gene. Microorganisms 2021, 9, 491. [Google Scholar] [CrossRef]
- Schmidt, C.; Löffler, B.; Ackermann, G. Antimicrobial phenotypes and molecular basis in clinical strains of Clostridium difficile. Diagn. Microbiol. Infect. Dis. 2007, 59, 1–5. [Google Scholar] [CrossRef]
- Kaushal, G.; Singh, S.P. Comparative genome analysis provides shreds of molecular evidence for reclassification of Leuconostoc mesenteroides MTCC 10508 as a strain of Leu. suionicum. Genomics 2020, 112, 4023–4031. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
 ) pH decrease wo/w 1% lactose, (
) pH decrease wo/w 1% lactose, (  ) pH increase wo/w 1% lactose, (
) pH increase wo/w 1% lactose, (  ) cit operon positive strains, (
) cit operon positive strains, (  ) no growth in minimum conditions, (
) no growth in minimum conditions, (  ) growth in minimum and/or maximum conditions.
) pH decrease wo/w 1% lactose, ( ) pH increase wo/w 1% lactose, ( ) cit operon positive strains, ( ) no growth in minimum conditions, ( ) growth in minimum and/or maximum conditions.
) growth in minimum and/or maximum conditions.
) pH decrease wo/w 1% lactose, ( ) pH increase wo/w 1% lactose, ( ) cit operon positive strains, ( ) no growth in minimum conditions, ( ) growth in minimum and/or maximum conditions.| P | Cheese ID | Strain ID | Subsp.-Id | Gram | Cat | EPS | CO2 | pH Growth Medium after (h) wo/w 1% Lactose | Cfu/g Cheese | Growth with % NaCl | Growth °C | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TYGS | (4 h) | (6 h) | (8 h) | (24 h) | (48 h) | 2% | 3% | 4.5% | 6.5% | 4 | 10 | 15 | 30 | 45 | ||||||||
| A | 2-1406 | INF2 | Ln. m. cremoris? | + | - | - | + | 6.6/6.4 | 6.6/6.4 | 6.5/5.8 | 5.8/4.6 | 4.8/4.4 | 89 | + | pg | - | - | - | + | + | + | - |
| 3-1406 | INF3b | Ln. m. mesenteroides | + | - | - | + | 6.6/6.4 | 6.6/6.4 | 6.6/5.2 | 5.9/4.4 | 5.3/4.4 | 39 | + | pg | - | - | + | + | + | + | - (+) | |
| B | 1-1404 | INF36 | Ln. m. mesenteroides | + | - | - | + | 6.5/6.7 | 6.4/6.6 | 6.4/5.8 | 5.8/5.0 | 5.3/4.6 | 58 | + | + | + | + | + | + | + | + | - (+) |
| 2-1404 | INF38 | Ln. m. mesenteroides | + | - | - | + | 6.5/6.5 | 6.4/6.3 | 6.4/6.1 | 5.9/5.2 | 5.8/4.9 | 61 | + | + | + | - | - | + | + | + | - | |
| C | 1453 | INF46 | Ln. m. mesenteroides | + | - | - | + | 6.4/6.6 | 6.4/6.6 | 6.0/6.1 | 5.7/5.4 | 5.4/4.7 | 41 | + | + | + | + | + | + | + | + | + |
| D | 1-3763 | INF82a | Ln. m. dextranicum | + | - | - | + | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| INF82b | Ln. m. mesenteroides | + | - | - | + | 5.5/5.8 | 5.4/5.7 | 5.3/5.5 | 5.2/5.4 | 5.1/5.3 | 72 | - | pg | + | pg | - | + | + | + | + | ||
| 3-3763 | INF90 | Ln m. mesenteroides | + | - | - | + | 5.8/6.0 | 5.9/5.8 | 5.8/5.4 | 5.7/5.3 | 5.7/5.1 | 53 | + | + | + | + | - | + | + | + | + | |
| INF92 | Ln. m. mesenteroides | + | - | - | - | 6.1/5.9 | 6.0/5.8 | 6.0/5.7 | 5.8/5.6 | 5.6/5.1 | 73 | - | + | + | pg | - | + | + | + | + | ||
| INF94 | Ln. m. mesenteroides | + | - | - | - | 5.7/6.0 | 5.7/5.7 | 5.6/5.4 | 5.4/5.2 | 5.2/5.2 | 47 | + | + | pg | pg | - | + | - | + | - | ||
| 4-3763 | INF98 | Ln. m. cremoris? | + | - | - | + | 5.4/6.0 | 5.4/5.9 | 5.4/5.8 | 5.2/5.8 | 5.2/5.5 | 63 | + | + | + | pg | - | + | + | + | + | |
| E | 4-3758 | INF103 | Ln. m. mesenteroides | + | - | - | - | 5.4/5.5 | 5.2/5.5 | 5.2/5.5 | 5.1/5.3 | 5.0/5.2 | 89 | + | + | pg | pg | - | - | + | + | + |
| F | 4-14246 | INF117 | Ln. m. jonggajibkimchii? | + | - | - | + | 6.0/6.2 | 5.9/5.7 | 5.9/5.7 | 5.8/5.6 | 5.7/5.5 | 92 | - | pg | + | + | - | + | + | + | + |
| G | Ž | INF 131 | Ln. m. dextranicum | + | - | - | + | 5.6/6.4 | 5.3/6.3 | 4.6/6.2 | 4.3/4.6 | 4.2/4.4 | 72 | + | + | + | + | - | + | + | + | - |
| H | 3-1387 | INF157 | Ln. m. mesenteroides | + | - | - | + | 6.4/6.6 | 6.3/6.6 | 6.2/5.8 | 5.8/5.5 | 5.4/4.9 | 41 | + | + | + | + | - | + | + | + | + |
| I | 4-1383 | INF166 | Ln. m. jonggajibkimchii? | + | - | - | + | 6.7/6.4 | 6.5/6.4 | 5.8/5.9 | 5.1/4.6 | 4.8/4.4 | 42 | + | + | + | - | - | + | + | + | - |
| Isolate-ID | Bacteriocin | Betalactone | T3PKS | Other |
|---|---|---|---|---|
| INF2 | IIc | + | + | Enterocin_x_chain_beta |
| INF3b | IIc, MesentericinY105 | - | + | Enterocin_x_chain_beta |
| INF36 | IIc | + | + | Enterocin_x_chain_beta |
| INF38 | IIc | + | + | Enterocin_x_chain_beta |
| INF46 | IIc | + | + | Enterocin_x_chain_beta |
| INF82a | IIc | + | + | Enterocin_x_chain_beta |
| INF90 | IIc | + LomaiviticinA//C-E | + | Enterocin_x_chain_beta |
| INF82b | IIc | LomaiviticinA//C-E | + | Enterocin_x_chain_beta |
| INF94 | IIc | LomaiviticinA//C-E | + | Enterocin_x_chain_beta |
| INF92 | IIc | - | + | Enterocin_x_chain_beta |
| INF117 | IIc | - | + | - |
| INF103 | IIc | - | + | - |
| INF98 | IIc | - | + | Enterocin_x_chain_beta |
| INF131 | MesentericinB105 | + | + | Enterocin_x_chain_beta |
| INF157 | IIc | - | + | Enterocin_x_chain_beta |
| INF166 | - | + | + | - |
| Strain ID | AMR | Plasmid Finder | Virulence Finder | Pathogen Finder | ||
|---|---|---|---|---|---|---|
| CARD | ResFinder | Susceptibility Testing | ||||
| INF2 | - | - | nd | Rep3 | - | - |
| INF3b | - | - | nd | Rep3 | - | - |
| INF36 | Clostridioides difficile 23S rRNA with C656T mutation conferring resistance to erythromycin and clindamycin, 98.97% identity, 10.00% coverage | - | S Ery (15 µg) Inhz 26 mm | - | - | - |
| INF38 | - | - | nd | - | - | - |
| INF46 | - | - | nd | - | - | - |
| INF82a | - | - | nd | - | - | - |
| INF82b | - | - | nd | - | - | - |
| INF90 | - | - | nd | - | - | - |
| INF92 | - | - | nd | Rep3 | - | - |
| INF94 | - | - | nd | - | - | - |
| INF98 | - | - | nd | Rep3 | - | - |
| INF103 | - | - | nd | - | - | - |
| INF117 | 97.5% identity to blaTEM 33, 55.94% coverage (using loose criteria) | beta lactam resistance 99.04% identity to blaTEM 141, 60.51% coverage | S Amp (10 µg) Inhz 19.5 mm Pen (1U) Inhz 21 mm | - | - | - |
| INF131 | - | - | nd | - | - | - |
| INF157 | - | - | nd | - | - | - |
| INF166 | - | - | nd | RepA_N | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruppitsch, W.; Nisic, A.; Hyden, P.; Cabal, A.; Sucher, J.; Stöger, A.; Allerberger, F.; Martinović, A. Genetic Diversity of Leuconostoc mesenteroides Isolates from Traditional Montenegrin Brine Cheese. Microorganisms 2021, 9, 1612. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081612
Ruppitsch W, Nisic A, Hyden P, Cabal A, Sucher J, Stöger A, Allerberger F, Martinović A. Genetic Diversity of Leuconostoc mesenteroides Isolates from Traditional Montenegrin Brine Cheese. Microorganisms. 2021; 9(8):1612. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081612
Chicago/Turabian StyleRuppitsch, Werner, Andjela Nisic, Patrick Hyden, Adriana Cabal, Jasmin Sucher, Anna Stöger, Franz Allerberger, and Aleksandra Martinović. 2021. "Genetic Diversity of Leuconostoc mesenteroides Isolates from Traditional Montenegrin Brine Cheese" Microorganisms 9, no. 8: 1612. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081612