Intrafollicular and Systemic Dopamine, Noradrenaline and Adrenaline Concentrations in Cycling Mares

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Collection of Blood and Ovaries
2.3. Collection of Follicular Fluid
2.4. Catecholamine Assay
2.5. Statistical Analyses
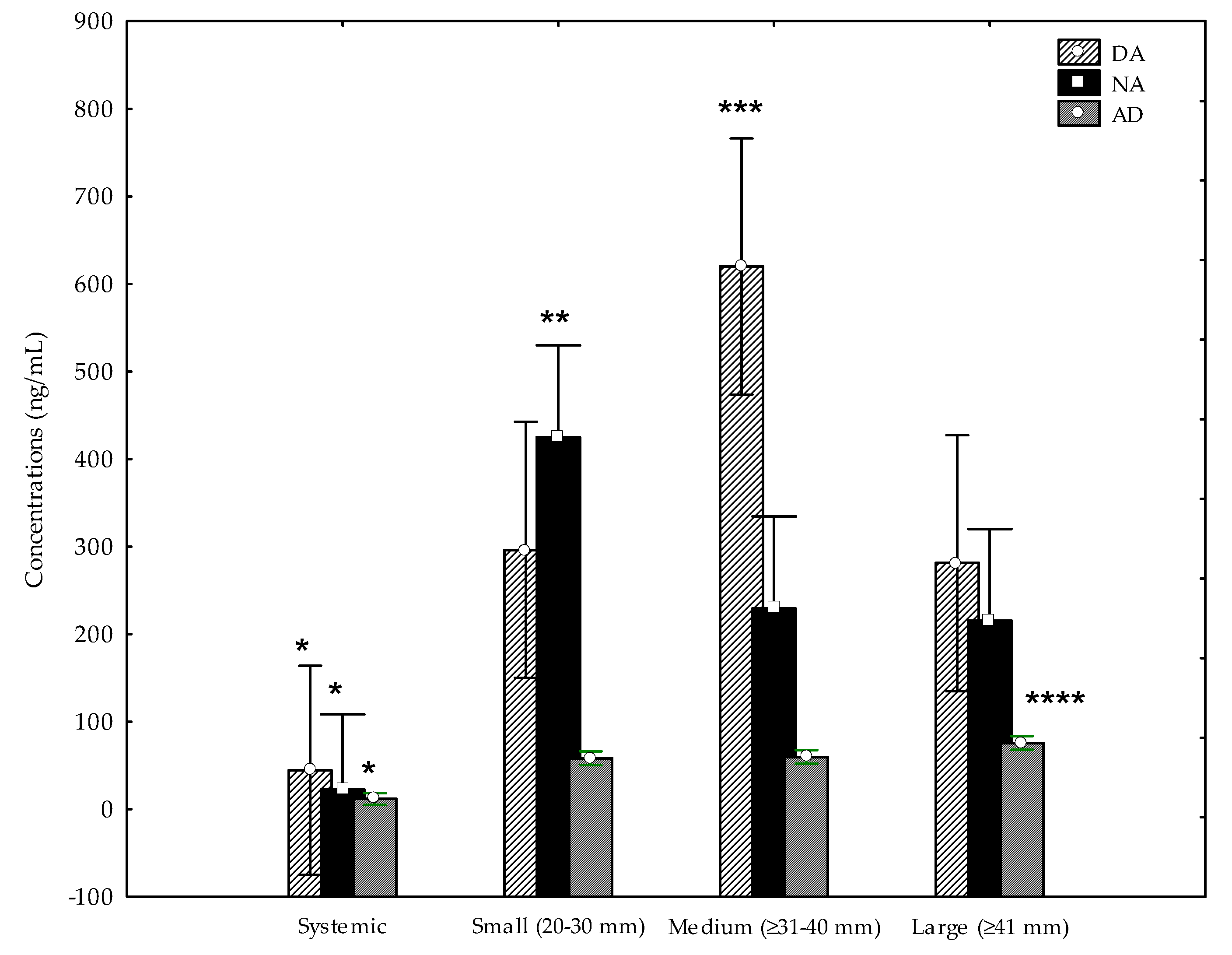
3. Results
4. Discussion
4.1. Intrafollicular Catecholamine Concentrations in Mares/Humans/Other Species
4.2. Ovarian Synthesis, Uptake, or Interconversion Mechanisms of Catecholamines in Mares/Primates/Other Species According to Receptors’ Expression
4.3. Effects of Systemic and Follicular Fluid Catecholamines on the Reproductive Physiology in Mares
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Melrose, P.A.; Walker, R.F.; Douglas, R.H. Dopamine in the cerebrospinal fluid of prepubertal and adult horses. Brain Behav. Evol. 1990, 35, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Daels, P.F.; Fatone, S.; Hansen, B.S.; Concannon, P.W. Dopamine antagonist-induced reproductive function in anoestrous mares: Gonadotrophin secretion and effects of environmental cues. J. Reprod. Fertil. 2000, 56, 173–183. [Google Scholar]
- Satué, K.; Gardon, J.C.; Marcilla, M. Physiology and Metabolic Anomalies of Dopamine in Horses: A Review; Open Access Peer-Reviewed Chapter; InTechOpen: London, UK, 2018; pp. 85–109. [Google Scholar] [CrossRef] [Green Version]
- Besognet, B.; Hansen, B.H.; Daels, P.F. Dopaminergic regulation of gonadotrophin secretion in seasonally anoestrous mares. J. Reprod. Fertil. 1996, 108, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besognet, B.; Hansen, B.H.; Daels, P.F. Induction of reproductive function in anestrous mares using a dopamine antagonist. Theriogenology 1997, 47, 467–480. [Google Scholar] [CrossRef]
- Bennett-Wimbush, K.; Loch, W.E.; Plata-Madrid, H.; Evans, T. The effect of perphenazine and bromocryptine on follicular dynamics and endocrine profiles in anestrous mares. Theriogenology 1998, 49, 717–733. [Google Scholar] [CrossRef]
- Panzani, D.; Zicchino, I.; Taras, A.; Marmorini, P.; Crisci, A.; Rota, A.; Camillo, F. Clinical use of dopamine antagonist sulpiride to advance first ovulation in transitional mares. Theriogenology 2011, 75, 138–143. [Google Scholar] [CrossRef]
- Thompson, D.L., Jr.; DePew, C.L. Prolactin, gonadotropin, and hair shedding responses to daily sulpiride administration in geldings in winter. J. Anim. Sci. 1997, 75, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Bódis, J.; Bognár, Z.; Hartmann, G.; Török, A.; Csaba, I.; Halvax, L. Analysis of noradrenaline, dopamine and serotonin levels in follicular fluid following superovulatory treatment. Orv. Hetil. 1991, 132, 2475–2477. [Google Scholar] [PubMed]
- Bódis, J.; Török, A.; Tinneberg, H.R.; Hanf, V.; Hamori, M.; Cledon, P. Influence of serotonin on progesterone and estradiol secretion of cultured human granulosa cells. Fertil. Steril. 1992, 57, 1008–1011. [Google Scholar] [CrossRef]
- Bódis, J.; Hartmann, G.; Török, A.; Bognár, Z.; Tinneberg, H.R.; Cledon, P.; Hanf, V. Relationship between the monoamine and gonadotropin content in follicular fluid of preovulatory graafian follicles after superovulation treatment. Exp. Clin. Endocrinol. 1993, 101, 178–182. [Google Scholar] [CrossRef]
- Bódis, J.; Hartmann, G.; Tinneberg, H.R.; Török, A.; Hanf, V.; Papenfuss, F.; Schwarz, H. Relationship between the monoamine, progesterone and estradiol content in follicular fluid of preovulatory graafian follicles after superovulation treatment. Gynecol. Obstet. Investig. 1993, 35, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Itoh, M.T.; Ishizuka, B.; Kuribayashi, Y.; Abe, Y.; Sumi, Y. Noradrenaline concentrations in human preovulatory follicular fluid exceed those in peripheral plasma. Exp. Clin. Endocrinol. Diabetes 2000, 108, 506–509. [Google Scholar] [CrossRef]
- Saller, S.; Kunz, L.; Berg, D.; Berg, U.; Lara, H.; Urra, J.; Hecht, S.; Pavlik, R.; Thaler, C.J.; Mayerhofer, A. Dopamine in human follicular fluid is associated with cellular uptake and metabolism-dependent generation of reactive oxygen species in granulosa cells: Implications for physiology and pathology. Hum. Reprod. 2014, 29, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musalı, N.; Özmen, B.; Şükür, Y.E.; Ergüder, B.İ.; Atabekoğlu, C.S.; Sönmezer, M.; Berker, B.; Aytaç, R. Follicular fluid norepinephrine and dopamine concentrations are higher in polycystic ovary syndrome. Gynecol. Endocrinol. 2016, 32, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; Arbogast, L.A.; Rhoades, T.A.; Bahr, J.M. Norepinephrine in the rat ovary: Ontogeny and de novo synthesis. Endocrinology 1984, 115, 1426–1431. [Google Scholar] [CrossRef]
- Bahr, J.M.; Ben-Jonathan, N. Elevated catecholamines in porcine follicular fluid before ovulation. Endocrinology 1985, 117, 620–623. [Google Scholar] [CrossRef]
- Fernández-Pardal, J.; Gimeno, M.F.; Gimeno, A.L. Catecholamines in sow graafian follicles at proestrus and at diestrus. Biol. Reprod. 1986, 34, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Kozłowska, A.; Majewski, M.; Jana, B. Changes in the cholinergic innervation pattern of porcine ovaries with cysts induced by dexamethasone administration. J. Mol. Neurosci. 2014, 54, 10–19. [Google Scholar] [CrossRef] [Green Version]
- King, S.S.; Jones, K.L.; Mullenix, B.A.; Heath, D.T.; Everson, K.A.; Arbogast, L.A. Evidence for dopaminergic activity in equine follicles and ovarian germinal epithelium. Anim. Reprod. Sci. 2006, 94, 175–178. [Google Scholar]
- Henneke, D.R.; Potter, G.D.; Kreider, J.L.; Yeates, B.F. Relationship between condition scoring, physical measurement and body fat percentage in mares. Equine Vet. J. 1983, 15, 371–372. [Google Scholar] [CrossRef]
- Hinrichs, K. Assisted reproduction techniques in the horse. Reprod. Fertil. Dev. 2012, 25, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Foss, R.; Ortis, H.; Hinrichs, K. Effect of potential oocyte transport protocols on blastocyst rates after intracytoplasmic sperm injection in the horse. Equine Vet. J. 2013, 45, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Marcilla, M.; Muñoz, A.; Satué, K. Longitudinal changes in serum catecholamines, dopamine, serotonin, ACTH and cortisol in pregnant Spanish mares. Res. Vet. Sci. 2017, 115, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Saller, S.; Merz-Lange, J.; Raffael, S.; Hecht, S.; Pavlik, R.; Thaler, C.; Berg, D.; Berg, U.; Kunz, L.; Mayerhofer, A. Norepinephrine, active norepinephrine transporter, and norepinephrine-metabolism are involved in the generation of reactive oxygen species in human ovarian granulosa cells. Endocrinology 2012, 153, 1472–1483. [Google Scholar] [CrossRef] [PubMed]
- Aguado, L.I. Role of the central and peripheral nervous system in the ovarian function. Microsc. Res. Tech. 2002, 59, 462–473. [Google Scholar] [CrossRef]
- Mayerhofer, A.; Smith, G.D.; Danilchik, M.; Levine, J.E.; Wolf, D.P.; Dissen, G.A.; Ojeda, S.R. Oocytes are a source of catecholamines in the primate ovary: Evidence for a cell-cell regulatory loop. Proc. Natl. Acad. Sci. USA 1998, 95, 10990–10995. [Google Scholar] [CrossRef] [Green Version]
- Paredes, A.H.; Salvetti, N.R.; Diaz, A.E.; Dallard, B.E.; Ortega, H.H.; Lara, H.E. Sympathetic nerve activity in normal and cystic follicles from isolated bovine ovary: Local effect of beta-adrenergic stimulation on steroid secretion. Reprod. Biol. Endocrinol. 2011, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Christian, C.A.; Moenter, S.M. The neurobiology of preovulatory and estradiol-induced gonadotropin-releasing hormone surges. Endocr. Rev. 2010, 31, 544–577. [Google Scholar] [CrossRef] [Green Version]
- Kotwica, G.; Kurowicka, B.; Franczak, A.; Grzegorzewski, W.; Wrobel, M.; Mlynarczuk, J.; Kotwica, J. The concentrations of catecholamines and oxytocin receptors in the oviduct and its contractile activity in cows during the estrous cycle. Theriogenology 2003, 60, 953–964. [Google Scholar] [CrossRef]
- Greiner, M.; Paredes, A.; Rey-Ares, V.; Saller, S.; Mayerhofer, A.; Lara, H.E. Catecholamine uptake, storage, and regulated release by ovarian granulosa cells. Endocrinology 2008, 149, 4988–4996. [Google Scholar] [CrossRef]
- Mari, G.; Morganti, M.; Merlo, B.; Castagnetti, C.; Parmeggiani, F.; Govoni, N.; Galeati, G.; Tamanini, C. Administration of sulpiride or domperidone for advancing the first ovulation in deep anestrous mares. Theriogenology 2009, 71, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Duchamp, J.; Daels, P.F. Combined effect of sulpiride and light treatment on the onset of cyclicity in anestrous mares. Theriogenology 2002, 58, 599–602. [Google Scholar]
- McCue, P.M.; Buchanan, B.R.; Farquhar, V.J.; Squires, E.L.; Cross, D.L. Efficacy of domperidone on induction of ovulation in anestrous and transitional mares. Proc. Annu. Conv. Am. Assoc. Equine Pract. 1999, 45, 217–218. [Google Scholar]
- Nagy, P.; Guillaume, D.; Daels, P. Seasonality in mares. Anim. Reprod. Sci. 2000, 60–61, 245–262. [Google Scholar] [CrossRef]
- McCue, P.M. Management of seasonal anestrus: Hormone therapy. In Equine Reproductive Procedures; Dascanio, J.J., McCue, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 146–148. [Google Scholar]
- King, S.S.; Jones, K.L.; Mullenix, B.A.; Heath, D.T. Seasonal relationships between dopamine D1 and D2 receptor and equine FSH receptor mRNA in equine ovarian epithelium. Anim. Reprod. Sci. 2008, 108, 259–266. [Google Scholar] [CrossRef]
- Rey-Ares, V.; Lazarov, N.; Berg, D.; Berg, U.; Kunz, L.; Mayerhofer, A. Dopamine receptor repertoire of human granulosa cells. Reprod. Biol. Endocrinol. 2007, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Merz, C.; Saller, S.; Kunz, L.; Xu, J.; Yeoman, R.R.; Ting, A.Y.; Lawson, M.S.; Stouffer, R.L.; Hennebold, J.D.; Pau, F.; et al. Expression of the beta-2 adrenergic receptor (ADRB-2) in human and monkey ovarian follicles: A marker of growing follicles? J. Ovarian Res. 2015, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Lara, H.E.; Porcile, A.; Espinoza, J.; Romero, C.; Luza, S.M.; Fuhrer, J.; Miranda, C.; Roblero, L. Release of norepinephrine from human ovary: Coupling to steroidogenic response. Endocrine 2001, 15, 187–192. [Google Scholar] [CrossRef]
- Dees, W.L.; Hiney, J.K.; Schultea, T.D.; Mayerhofer, A.; Danilchik, M.; Dissen, G.A.; Ojeda, S.R. The primate ovary contains a population of catecholaminergic neuron-like cells expressing nerve growth factor receptors. Endocrinology 1995, 136, 5760–5768. [Google Scholar] [CrossRef]


{kind=link}
| Parameters | N (90) | r | Equation of Regression Line |
|---|---|---|---|
| DA (ng/mL) | 0.64 | Systemic/FF = 1.3557 + 0.99 * | |
| NA (ng/mL) | 0.67 | Systemic/FF = 1.3410 + 0.00154 * | |
| AD (ng/mL) | 0.93 | Systemic/FF = 0.90257 + 0.01575 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satué, K.; Fazio, E.; Rubio, M.D.; Cravana, C.; Medica, P. Intrafollicular and Systemic Dopamine, Noradrenaline and Adrenaline Concentrations in Cycling Mares. Animals 2020, 10, 1896. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10101896
Satué K, Fazio E, Rubio MD, Cravana C, Medica P. Intrafollicular and Systemic Dopamine, Noradrenaline and Adrenaline Concentrations in Cycling Mares. Animals. 2020; 10(10):1896. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10101896
Chicago/Turabian StyleSatué, Katiuska, Esterina Fazio, Maria Dolores Rubio, Cristina Cravana, and Pietro Medica. 2020. "Intrafollicular and Systemic Dopamine, Noradrenaline and Adrenaline Concentrations in Cycling Mares" Animals 10, no. 10: 1896. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10101896