
Assessment of Postnatal Pulmonary Adaption in Bovine Neonates Using Electric Impedance Tomography (EIT)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
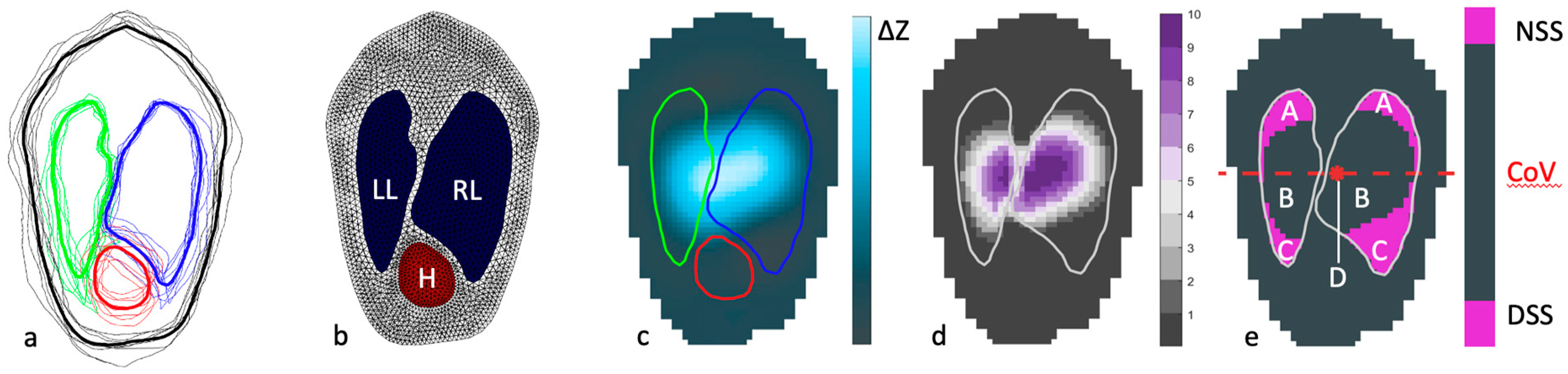
2.2. Measurements Using Electric Impedance Tomography
2.2.1. Centre of Ventilation
2.2.2. Relative Tidal Stretch and Silent Spaces
2.2.3. Global and Regional Ventilation of the Right and Left Lung
2.2.4. Respiration Rate, Tidal Impedance Change, and Minute Tidal Impedance Change
2.3. Blood Gas Analysis
2.3.1. Alveolar–Arterial Oxygen Difference
2.3.2. Oxygen Content-Based Index (Fshunt)
2.4. Statistical Analysis
3. Results
3.1. Clinical Variables
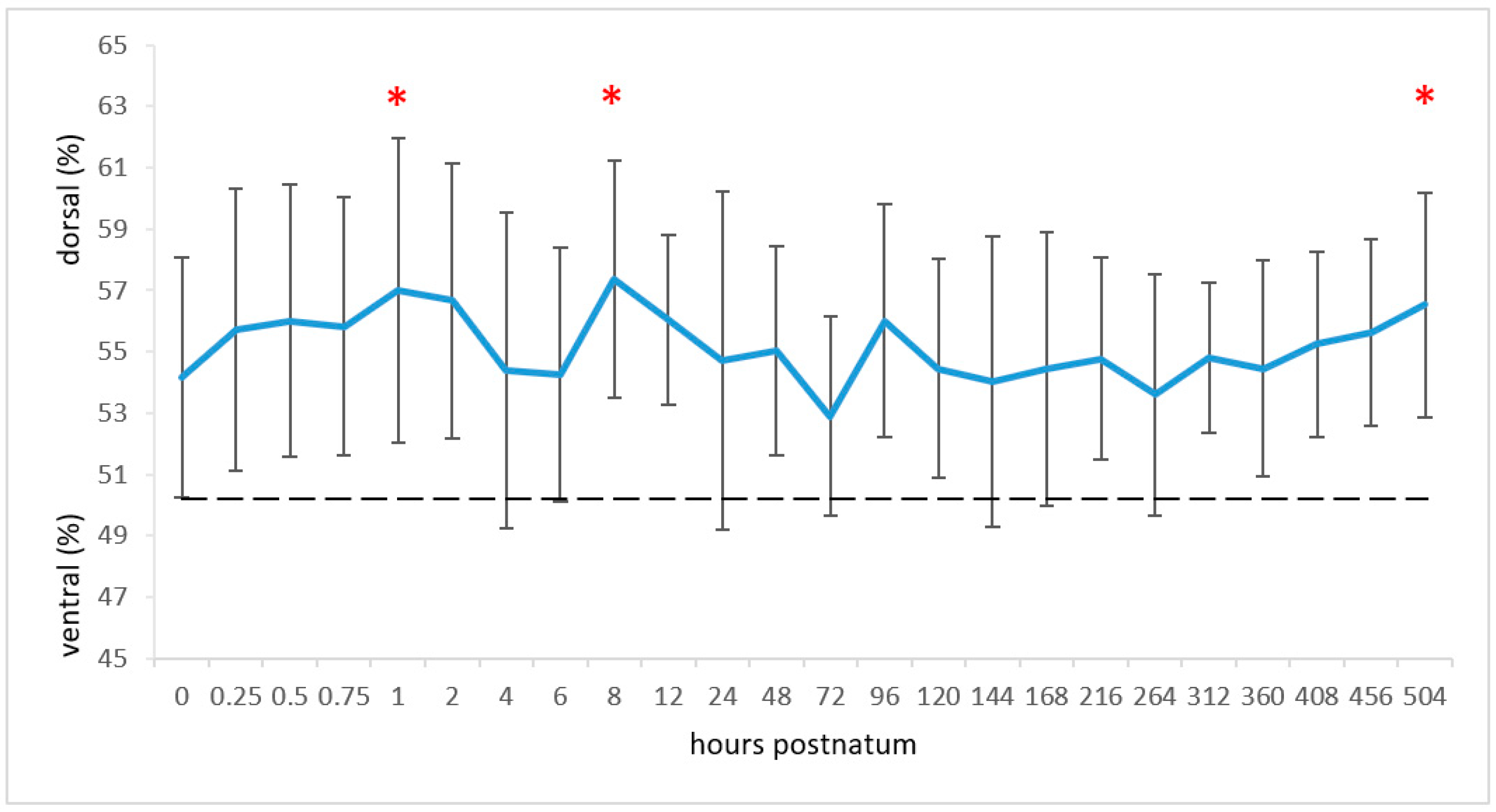
3.2. Centre of Ventilation
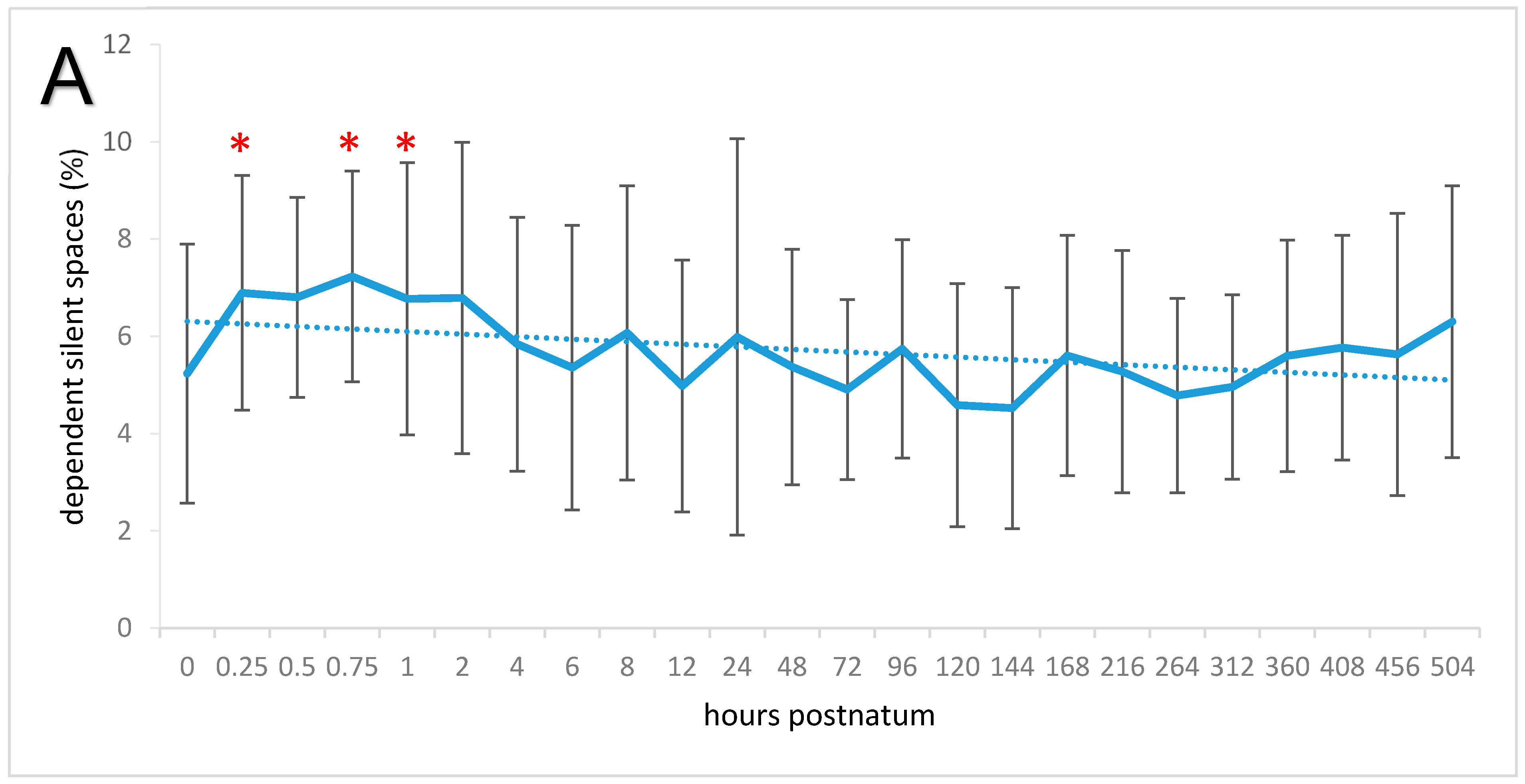
3.3. Relative Tidal Stretch and Silent Spaces
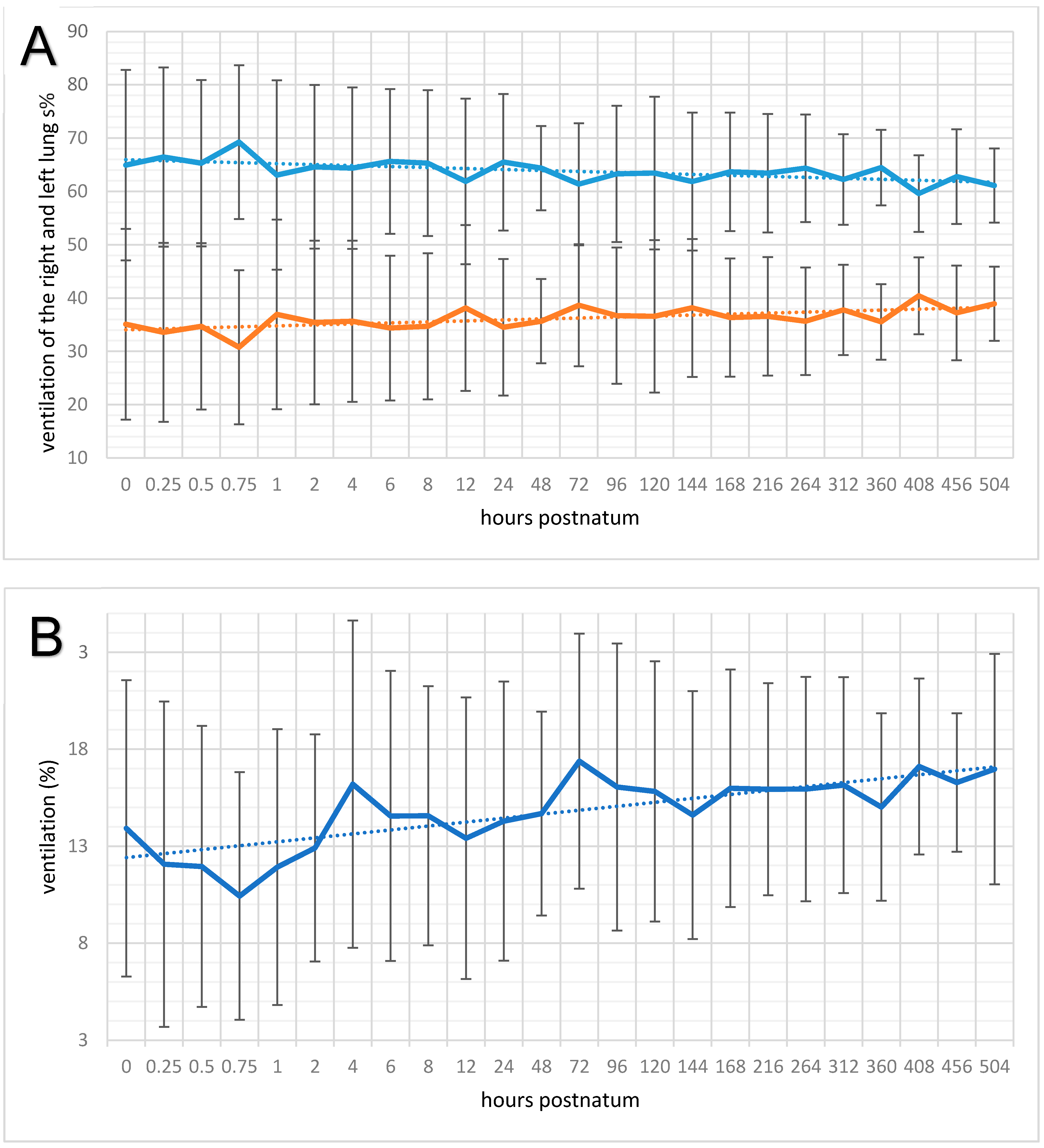
3.4. Regional Ventilation of the Right and Left Lung
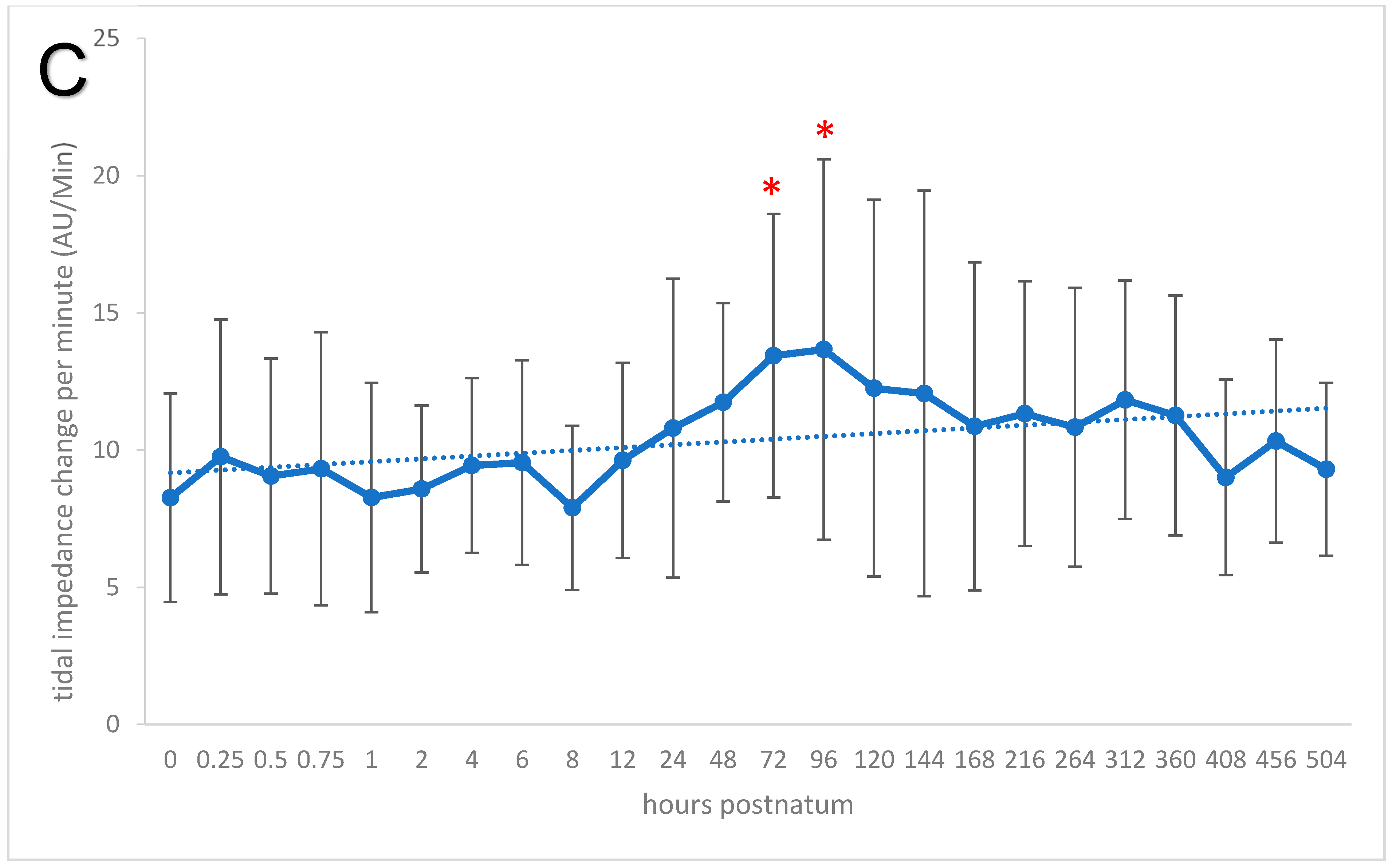
3.5. Respiration Rate, Global Ventilation (ΔZVT), and Minute Tidal Impedance Change
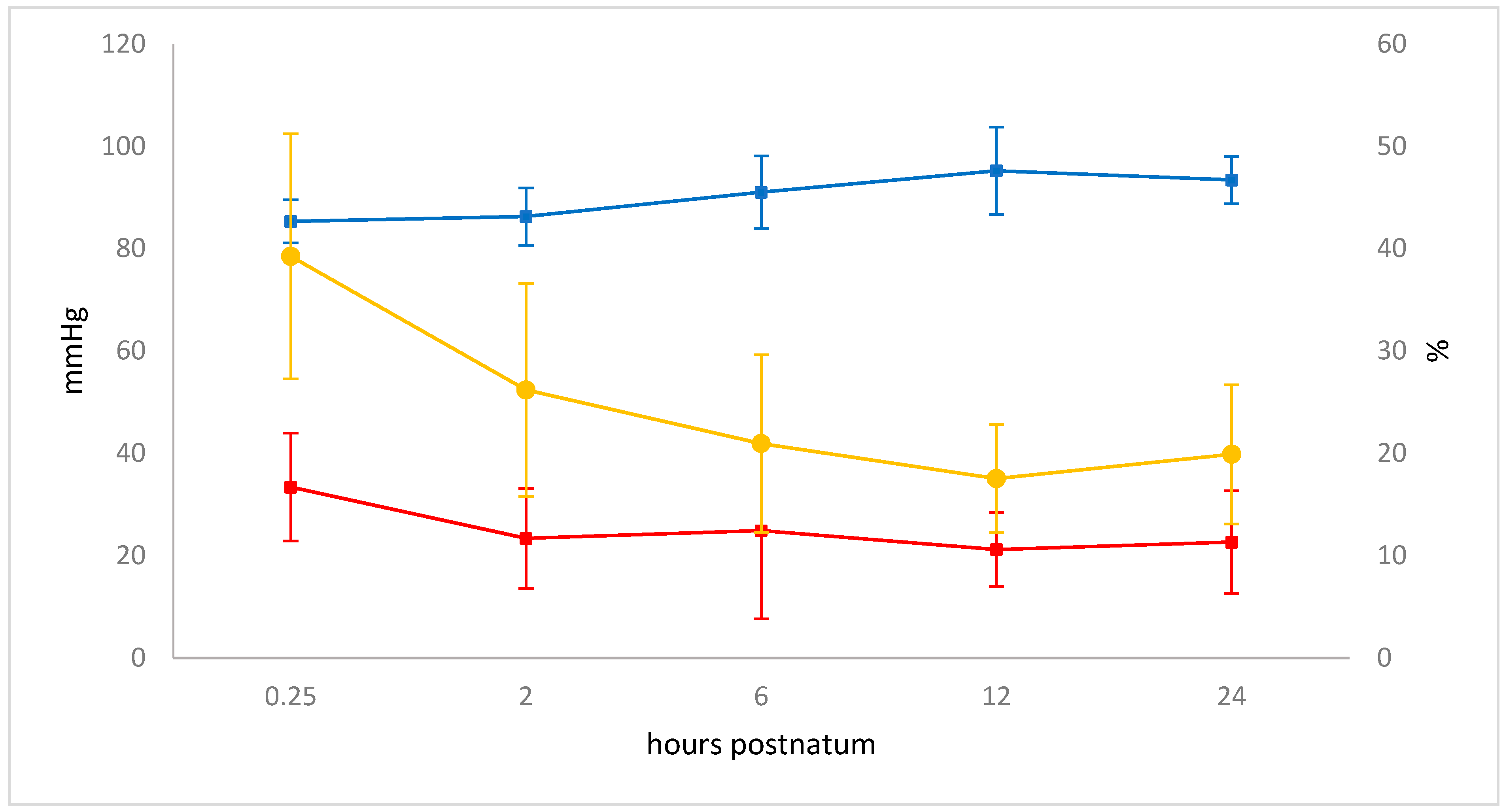
3.6. Blood Gas Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hooper, S.B.; Polglase, G.R.; Roehr, C.C. Cardiopulmonary Changes with Aeration of the Newborn Lung. Paediatr. Respir. Rev. 2015, 16, 147–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortola, J.P. Respiratory Physiology of Newborn Mammals: A Comparative Perspective; Johns Hopkins University Press: Baltimore, MD, USA, 2001; p. 344. [Google Scholar]
- Fawcitt, J.; Lind, J.; Wegelius, C. The First Breath: A Preliminary Communication Describing Some Methods of Investigation of the First Breath of a Baby and the Results Obtained from Them. Acta Paediatr. 1960, 49, 5–17. [Google Scholar] [CrossRef]
- Karlberg, P.; Cherry, R.B.; Escardó, F.E.; Koch, G. Respiratory Studies in Newborn Infants. II: Pulmonary Ventilation and Mechanics of Breathing in the First Minutes of Life, Including the Onset of Respiration. Acta Paediatr. 1962, 51, 121–136. [Google Scholar] [CrossRef]
- Lind, J. Changes in the Circulation and Lungs at Birth. Acta Paediatr. Supplementum. 1960, 49 (Suppl. 122), 39–52. [Google Scholar] [CrossRef]
- Milner, A.D.; Vyas, H. Lung Expansion at Birth. J. Pediatr. 1982, 101, 879–886. [Google Scholar] [CrossRef]
- Bleul, U.; Lejeune, B.; Schwantag, S.; Kahn, W. Blood gas and acid-base analysis of arterial blood in 57 newborn calves. Vet. Rec. 2007, 161, 688–691. [Google Scholar] [CrossRef]
- Neary, J.; Garry, F.; Raabis, S. Age-Related Changes in Arterial Blood-Gas Variables in Holstein Calves at Moderate Altitude. Open Access Anim. Physiol. 2014, 6, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Norton, J.; Munro, J.; Taylor, P.; Jackson, P. Measuring Arterial Oxygen Saturation in Neonatal Calves using Pulse Oximetry. Vet. Rec. 1996, 138, 157–164. [Google Scholar]
- Bartels, H.; Riegel, K.; Wenner, J.; Wulf, H. Perinatale Atmung: Physiologische Grundlagen und Therapeutische Konsequenzen; Springer: Berlin/Heidelberg, Germany, 1972. [Google Scholar]
- Karlberg, P.; Koch, G. Respiratory Studies in Newborn Infants. III.: Development of Mechanics of Breathing during the First Week of Life. A Longitudinal Study1. Acta Paediatr. 1962, 51, 121–129. [Google Scholar] [CrossRef]
- Jung, C. Sonographie der Lunge und des Abdomens beim Bovinen Neonaten unter Besonderer Berücksichtigung Pathologischer Veränderungen; Justus-Liebig University: Giessen, Germany, 2002. [Google Scholar]
- Abdelmegeid, M.K.; Kutasi, O.; Nassiff, M.N.; Wirth, K.; Bodai, E.; Farrag, F.; Molnár, L.; Szenci, O. Radiographic Assessment of Pulmonary Fluid Clearance and Lung Aeration in Newborn Calves Delivered by Elective Caesarean Section. Reprod. Domest. Anim. 2017, 52, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Linke, B.; Bostedt, H.; Richter, A. Computer Tomographic Illustration of the Development of the Pulmonary Function in Bovine Neonates until the Twenty-First Day Postnatum. Vet. Med. Int. 2013, 2013, 12. [Google Scholar] [CrossRef] [Green Version]
- Frerichs, I.; Amato, M.B.P.; van Kaam, A.H.; Tingay, D.G.; Zhao, Z.; Grychtol, B.; Bodenstein, M.; Gagnon, H.; Böhm, S.H.; Teschner, E.; et al. Chest Electrical Impedance Tomography Examination, Data Analysis, Terminology, Clinical Use and Recommendations: Consensus Statement of the Translational EIT Development Study Group. Thorax 2017, 72, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Sacks, M.; Byrne, D.; Herteman, N.; Secombe, C.; Adler, A.; Raisis, A.; Mosing, M. Comparison of distribution of ventilation between healthy horses and horses with left-sided cardiac volume overload measured by Electrical Impedance Tomography. J. Vet. Int. Med. 2021, 35, 2511–2523. [Google Scholar] [CrossRef] [PubMed]
- Waldmann, A.; Meira, C.; Auer, U.; Böhme, S.; Braun, C.; Bohm, S.; Mosing, M. Construction of a robust beagle model for eit applications. In Proceedings of the 17th International Conference on Electrical Impedance Tomography, Stockholm, Sweden, 19–23 June 2016; p. 158. [Google Scholar]
- Sophocleous, L.; Frerichs, I.; Miedema, M.; Kallio, M.; Papadouri, T.; Karaoli, C.; Becher, T.; Tingay, D.G.; van Kaam, A.H.; Bayford, R.; et al. Clinical performance of a novel textile interface for neonatal chest electrical impedance tomography. Physiol. Meas. 2018, 39, 044004. [Google Scholar] [CrossRef] [Green Version]
- Lobo, B.; Hermosa, C.; Abella, A.; Gordo, F. Electrical Impedance Tomography. Ann. Transl. Med. 2018, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, K.S.; Kabir, A.M. Studies on the Effect of the Third Dimension on a Two-Dimensional Electrical Impedance Tomography System. Clin. Phys. Physiol. Meas. 1991, 12, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Mülling, M. Asphyxie des neugeborenen Kalbes. Prakt. Tierarzt Coll. Vet. 1976, 58, 78–80. [Google Scholar]
- Ennen, S.; Wehrend, A. Der segmentale Untersuchungsgang für neonatale Fohlen als standardisiertes Verfahren. Prakt. Tierarzt 2010, 91, 222–229. [Google Scholar]
- Waldmann, A.; Meira, C.; Auer, U.; Böhme, S.; Braun, C.; Bohm, S.; Mosing, M. Contribution of finite element model of animals to data base. In Proceedings of the 17th International Conference on Electrical Impedance Tomography, Stockholm, Sweden, 19–23 June 2016; p. 161. [Google Scholar]
- Grychtol, B.; Lionheart, W.R.B.; Bodenstein, M.; Wolf, G.K.; Adler, A. Impact of Model Shape Mismatch on Reconstruction Quality in Electrical Impedance Tomography. IEEE Trans. Med. Imaging 2012, 31, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Abé, M. Computertomographische Untersuchung des Thorax von Kälbern von der Geburt bis zum Alter von 105 Tagen; Univesität Zürich: Zürich, Germany, 2013. [Google Scholar]
- Ohlerth, S.; Augsburger, H.; Abé, M.; Ringer, S.; Hatz, L.; Braun, U. Computed Tomography of the Thorax in Calves from Birth to 105 Days of Age. Schweiz Arch. Tierheilkd 2014, 156, 489–497. [Google Scholar] [CrossRef]
- Frerichs, I.; Hahn, G.; Golisch, W.; Kurpitz, M.; Burchardi, H.; Hellige, G. Monitoring Perioperative Changes in Distribution of Pulmonary Ventilation by Functional Electrical Impedance Tomography. Acta Anaesthesiol. Scand. 1998, 42, 721–726. [Google Scholar] [CrossRef]
- Radke, O.C.; Schneider, T.; Heller, A.R.; Koch, T. Spontaneous Breathing During General Anesthesia Prevents the Ventral Redistribution of Ventilation as Detected by Electrical Impedance Tomography: A Randomized Trial. Anesthesiology 2012, 116, 1227–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldmann, A.; Brunner, J.X.; Böhm, S. Swisstom BB2 Silent Spaces: Lung Function Parameters Explained. Available online: https://www.researchgate.net/publication/277257764_EIT_Analysis_Details_Lung_Stretch_and_Silent_Spaces_analysis#fullTextFileContent (accessed on 25 August 2020).
- Ukere, A.; März, A.; Wodack, K.H.; Trepte, C.J.; Haese, A.; Waldmann, A.D.; Böhm, S.H.; Reuter, D.A. Perioperative Assessment of Regional Ventilation During Changing Body Positions and Ventilation Conditions by Electrical Impedance Tomography. Br. J. Anaesth. 2016, 117, 228–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodadad, D.; Nordebo, S.; Müller, B.; Waldmann, A.; Yerworth, R.; Becher, T.; Frerichs, I.; Sophocleous, L.; van Kaam, A.; Miedema, M.; et al. Optimized breath detection algorithm in electrical impedance tomography. Physiol. Meas. 2018, 39, 094001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, O.; Kovac, G.; Seidel, H.; Paulikova, I. Selection of arteries for blood sampling and evaluation of blood gases and acid-base balance in cattle. Acta Vet. Brno 2002, 71, 289–296. [Google Scholar] [CrossRef] [Green Version]
- MeteoSchweiz. Messwerte. Available online: https://www.meteoschweiz.admin.ch/home/messwerte.html?param=messwerte-luftdruck-qfe-10min&station=SMA&chart=day (accessed on 25 August 2020).
- Cambier, C.; Detry, B.; Beerens, D.; Florquin, S.; Ansay, M.; Frans, A.; Clerbaux, T.; Gustin, P. Effects of Hyperchloremia on Blood Oxygen Binding in Healthy Calves. J. Appl. Physiol. 1998, 85, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Araos, J.D.; Larenza, M.P.; Boston, R.C.; Monte, V.d.; Marzo, C.d.; Grasso, S.; Haskins, S.C.; Crovace, A.; Staffieri, F. Use of the Oxygen Content-Based Index, Fshunt, as an Indicator of Pulmonary Venous Admixture at Various Inspired Oxygen Fractions in Anesthetized Sheep. Am. J. Vet. Res. 2012, 73, 2013–2020. [Google Scholar] [CrossRef]
- Cambier, C.; Clerbaux, T.; Detry, B.; Marville, V.; Frans, A.; Gustin, P. Blood oxygen binding in hypoxaemic calves. Vet. Res. 2002, 33, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, R.A.; Davison, R.; Shapiro, B.A.; Meyers, S.N. Reassessment of the Assumed A-V Oxygen Content Difference in the Shunt Calculation. Anesth. Analg. 1975, 54, 198–202. [Google Scholar] [CrossRef]
- Kallio, M.; Rahtu, M.; van Kaam, A.H.; Bayford, R.; Rimensberger, P.C.; Frerichs, I. Electrical impedance tomography reveals pathophysiology of neonatal pneumothorax during NAVA. Clin. Case Rep. 2020, 8, 1574–1578. [Google Scholar] [CrossRef]
- Miedema, M.; Adler, A.; McCall, K.E.; Perkins, E.J.; van Kaam, A.H.; Tingay, D.G. Electrical impedance tomography identifies a distinct change in regional phase angle delay pattern in ventilation filling immediately prior to a spontaneous pneumothorax. J. Appl Physiol. 2019, 127, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Tingay, D.G.; Farrell, O.; Thomson, J.; Perkins, E.J.; Pereira-Fantini, P.M.; Waldmann, A.D.; Rüegger, C.; Adler, A.; Davis, P.G.; Frerichs, I. Imaging the Respiratory Transition at Birth: Unravelling the Complexities of the First Breaths of Life. Am. J. Resp. Crit. Care 2021. [Google Scholar] [CrossRef]
- Tingay, D.G.; Pereira-Fantini, P.M.; Oakley, R.; McCall, K.E.; Perkins, E.J.; Miedema, M.; Sourial, M.; Thomson, J.; Waldmann, A.; Dellaca, R.L.; et al. Gradual Aeration at Birth Is More Lung Protective Than a Sustained Inflation in Preterm Lambs. Am. J. Resp Crit Care 2019, 200, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Frerichs, I.; Hahn, G.; Hellige, G. Gravity-Dependent Phenomena in Lung Ventilation Determined by Functional EIT. Physiol. Meas. 1996, 17, A149–A157. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.W.; Counsell, S.J.; Hajnal, J.V.; Cox, P.N.; Kennea, N.L.; Thornton, A.S.; Bryan, A.C.; Edwards, A.D. Magnetic Resonance Imaging of Lung Water Content and Distribution in Term and Preterm Infants. Am. J. Respir. Crit. Care Med. 2002, 166, 397–402. [Google Scholar] [CrossRef]
- Kunst, P.W.A.; Noordegraaf, A.V.; Raaijmakers, E.; Bakker, J.; Groeneveld, A.B.J.; Postmus, P.E.; de Vries, P.M.J.M. Electrical Impedance Tomography in the Assessment of Extravascular Lung Water in Noncardiogenic Acute Respiratory Failure. Chest 1999, 116, 1695–1702. [Google Scholar] [CrossRef] [PubMed]
- Trepte, C.J.C.; Phillips, C.R.; Solà, J.; Adler, A.; Haas, S.A.; Rapin, M.; Böhm, S.H.; Reuter, D.A. Electrical impedance tomography (EIT) for quantification of pulmonary edema in acute lung injury. Crit. Care 2016, 20, 18. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, S.R.; Henderson, A.C.; Levin, D.L.; Yamada, K.; Arai, T.; Buxton, R.B.; Prisk, G.K. Vertical gradients in regional lung density and perfusion in the supine human lung: The Slinky effect. J. Appl. Physiol. 2007, 103, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Prisk, G.K. Microgravity and the respiratory system. Eur. Respir. J. 2014, 43, 1459. [Google Scholar] [CrossRef]
- Bland, R.D.; Hansen, T.N.; Haberkern, C.M.; Bressack, M.A.; Hazinski, T.A.; Raj, J.U.; Goldberg, R.B. Lung Fluid Balance in Lambs Before and After Birth. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1982, 53, 992–1004. [Google Scholar] [CrossRef]
- Bland, R.D.; McMillan, D.D.; Bressack, M.A.; Dong, L. Clearance of Liquid from Lungs of Newborn Rabbits. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1980, 49, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Kutasi, O.; Horvath, A.; Harnos, A.; Szenci, O. Radiographic Assessment of Pulmonary Fluid Clearance in Healthy Neonatal Foals. Vet. Radiol. Ultrasound. 2009, 50, 584–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maamoun, W.; Fort, A.E.; Cummings, J.J. Neonatal respiratory disease. In Pediatric Critical Care; Fuhrman, B.P., Zimmerman, J.J., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2011; pp. 590–680. [Google Scholar]
- Vannucchi, C.I.; Silva, L.C.G.; Unruh, S.M.; Lúcio, C.F.; Veiga, G.A.L. Calving duration and obstetric assistance influence pulmonary function of Holstein calves during immediate fetal-to-neonatal transition. PLoS ONE 2018, 13, e0204129. [Google Scholar] [CrossRef] [Green Version]
- König, H.E.; Liebich, H.-G. Apparatus respiratorius. In Anatomie der Haussäugetiere, 6th ed.; König, H.E., Liebich, H.-G., Eds.; Schattauer: Stuttgart, Germany, 2005; pp. 373–394. [Google Scholar]
- Frerichs, I.; Schiffmann, H.; Oehler, R.; Dudykevych, T.; Hahn, G.; Hinz, J.; Hellige, G. Distribution of Lung Ventilation in Spontaneously Breathing Neonates Lying in Different Body Positions. Intensive Care Med. 2003, 29, 787–794. [Google Scholar] [CrossRef]
- Ambrisko, T.D.; Schramel, J.P.; Adler, A.; Kutasi, O.; Makra, Z.; Moens, Y.P.S. Assessment of Distribution of Ventilation by Electrical Impedance Tomography in Standing Horses. Physiol. Meas. 2016, 37, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brabant, O.; Crivellari, B.; Hosgood, G.; Raisis, A.; Waldmann, A.D.; Auer, U.; Adler, A.; Smart, L.; Laurence, M.; Mosing, M. Effects of PEEP on the relationship between tidal volume and total impedance change measured via electrical impedance tomography (EIT). J. Clin. Monit. Comput. 2021. [Google Scholar] [CrossRef]
- Lehmann, S.; Leonhardt, S.; Ngo, C.; Bergmann, L.; Schrading, S.; Heimann, K.; Wagner, N.; Tenbrock, K. Electrical impedance tomography as possible guidance for individual positioning of patients with multiple lung injury. Clin. Respir. J. 2018, 12, 68–75. [Google Scholar] [CrossRef]
- Uystepruyst, C.H.; Reinhold, P.; Coghe, J.; Bureau, F.; Lekeux, P. Mechanics of the respiratory system in healthy newborn calves using impulse oscillometry. Res. Vet. Sci. 2000, 68, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Mester, L.; Borzsonyi, L.; Erdesz, C.; Vari, A.; Kormoczi, P.S.; Szenci, O. Adaptation of respiration to extrauterine-life in healthy newborn calves. Reprod. Domest. Anim. 1999, 34, 377–379. [Google Scholar] [CrossRef]
- Reinhold, P. Atmungsphysiologie im Spezies-Vergleich. Pneumologie 2008, 62, A11. [Google Scholar] [CrossRef]
- Silva, B.; Henklein, A.; Marques, R.; Oliveira, P.; Leite, S.; Novo, S.; Baccili, C.; Reis, J.; Gomes, V. Vital Parameters of Holstein Calves from Birth to Weaning. Rev. Bras. De Med. Vet. 2016, 38, 299–304. [Google Scholar]
- Gros, G. Atmung: Regulation der Atmung. In Physiologie der Haustiere, 5th ed.; Engelhardt, W.V., Breves, G., Diener, M., Gäbel, G., Eds.; Enke: Stuttgart, Germany, 2015; pp. 288–291. [Google Scholar]
- Taeusch, H.W.; Carson, S.; Frantz, I.D.; Milic-Emili, J. Respiratory regulation after elastic loading and CO2 rebreathing in normal term infants. J. Pediatr. 1976, 88, 102–111. [Google Scholar] [CrossRef]
- Fisher, J.T.; Mortola, J.P.; Smith, J.B.; Fox, G.S.; Weeks, S. Respiration in Newborns: Development of the Control of Breathing. Am. Rev. Respir. Dis. 1982, 125, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.H.; Rose, R.J.; Barko, A.M. Respiratory Studies in Foals from Birth to Seven Days Old. Equine Vet. J. 1984, 16, 323–328. [Google Scholar] [CrossRef]
- Mosing, M.; Waldmann, A.D.; Raisis, A.; Böhm, S.H.; Drynan, E.; Wilson, K. Monitoring of Tidal Ventilation by Electrical Impedance Tomography in Anaesthetised Horses. Equine Vet. J. 2019, 51, 222–226. [Google Scholar] [CrossRef]
- Victorino, J.A.; Borges, J.B.; Okamoto, V.N.; Matos, G.F.J.; Tucci, M.R.; Caramez, M.P.R.; Tanaka, H.; Sipmann, F.S.; Santos, D.C.B.; Barbas, C.S.V.; et al. Imbalances in Regional Lung Ventilation: A Validation Study on Electrical Impedance Tomography. Am. J. Respir. Crit. Care Med. 2004, 169, 791–800. [Google Scholar] [CrossRef]
- Oczenski, W. Atmen—Atemhilfen: Atemphysiologie und Beatmungstechnik; Thieme: Stuttgart, Germany, 2012. [Google Scholar]
- Brisville, A.C.; Fecteau, G.; Boysen, S.; Dorval, P.; Buczinski, S.; Blondin, P.; Smith, L.C. Respiratory Disease in Neonatal Cloned Calves. J. Vet. Intern. Med. 2011, 25, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekeux, P. Pulmonary Function in Healthy, Exercising and Diseased Animals; Gent State University: Gent, Belgium, 1993; p. 402. [Google Scholar]
- Heymann, M.A. Control of the Pulmonary Circulation in the Fetus and During the Transitional Period to Air Breathing. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 84, 127–132. [Google Scholar] [CrossRef]
- Rudolph, A.M. Fetal and Neonatal Pulmonary Circulation. Annu. Rev. Physiol. 1979, 41, 383–395. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 min (v) | 15 min (a) | 2 h (a) | 6 h (a) | 12 h (a) | 24 h (a) | |
|---|---|---|---|---|---|---|
| pH | 7.21 ± 0.08 | 7.29 ± 0.06 | 7.33 ± 0.04 | 7.34 ± 0.04 * | 7.39 ± 0.03 * | 7.41 ± 0.03 * |
| pO2 (mmHg) | 26.77 ± 5.94 | 51.93 ± 12.54 | 62.87 ± 12.89 | 69.32 ± 10.58 * | 74.01 ± 12.27 * | 71.49 ± 12.50 * |
| pCO2 (mmHg) | 67.53 ± 8.62 | 55.29 ± 4.01 | 54.36 ± 5.27 | 50.42 ± 6.61 | 45.33 ± 8.37 * | 47.15 ± 4.47 * |
| sO2 (%) | 50.70 ± 14.86 | 85.93 ± 7.71 | 93.05 ± 3.69 * | 94.80 ± 2.79 * | 95.67 ± 1.78 * | 94.93 ± 2.09 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bleul, U.; Wey, C.; Meira, C.; Waldmann, A.; Mosing, M. Assessment of Postnatal Pulmonary Adaption in Bovine Neonates Using Electric Impedance Tomography (EIT). Animals 2021, 11, 3216. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113216
Bleul U, Wey C, Meira C, Waldmann A, Mosing M. Assessment of Postnatal Pulmonary Adaption in Bovine Neonates Using Electric Impedance Tomography (EIT). Animals. 2021; 11(11):3216. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113216
Chicago/Turabian StyleBleul, Ulrich, Corina Wey, Carolina Meira, Andreas Waldmann, and Martina Mosing. 2021. "Assessment of Postnatal Pulmonary Adaption in Bovine Neonates Using Electric Impedance Tomography (EIT)" Animals 11, no. 11: 3216. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113216