Data analyzed within this paper were derived from a single farm. This reflected the fact that the determination of daily EB values relied on the use of individual cow intake data, which are generally only recorded on research farms. However, a wide range of diet types and qualities were examined within the studies used in the analysis. Furthermore, while the genetic potential of cows in the research herd changed over time, this genetic variability within the dataset made the results highly applicable to commercial farms where genetic variation both within herds and between herds can be substantial.
In addition to characterizing the relationships between FPR and EB during the first 20 WIM, this paper sought to examine how the main drivers of EB (DMI and ECM) and other commonly used EB indicators (BW changes, BHB, and NEFA) might help explain FPR–EB relationships across the stage of lactation and parity in Holstein-Friesian dairy cows. The mean forage-to-concentrate proportion in diets offered across the 27 experiments was 49:51 (on a DM basis), with this ratio varying little across lactations; thus, minimizing possible confounding effects arising from differences in energy supply. This concentrate proportion in the diet is typical of that in diets offered to moderate–high-yielding dairy herds within grass silage-based systems in the UK.
4.1. Trends in Intakes, Milk Production, Blood Metabolites, FPR, and EB
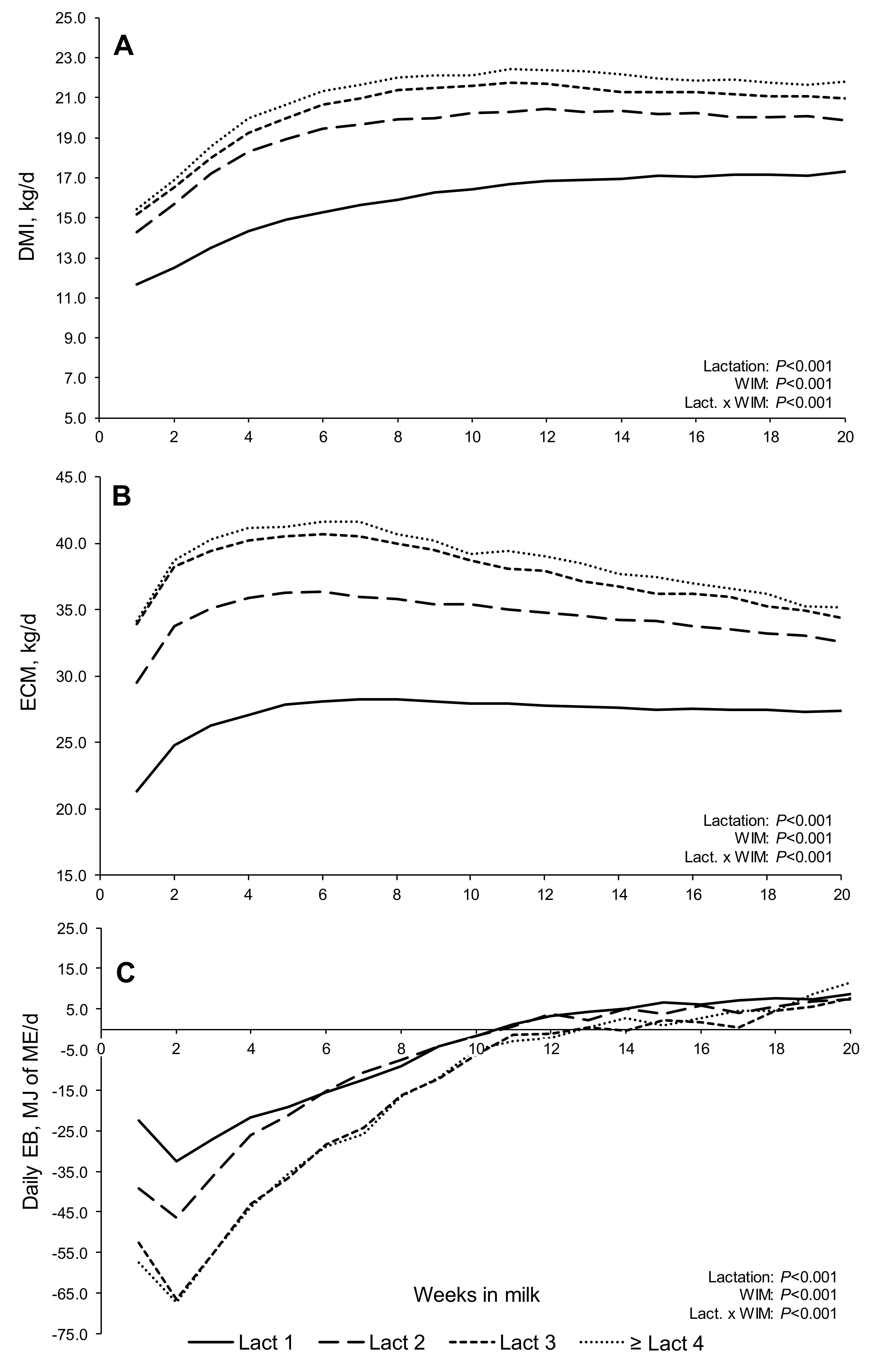
An examination of weekly trends in DMI, milk production, milk composition, EB, BW, BCS, and blood metabolites allowed changes in FPR to be placed within a wider context. The trend for DMI and ECM to increase from lactations one to three within the lactation profiles reflected the increasing size (reflected in increasing BW) of the cows with increasing parity, their correspondingly greater intake and milk production potential, and their increased capacity for tissue mobilization in support of lactation, resulting in an increased duration of NEB [
23]. In support of this, in the present study, both first and second lactation cows returned to positive EB, approximately, two weeks earlier than cows in lactations three and ≥ four (on average at 10.5 vs. 12.5 WIM, respectively). Furthermore, during the first lactation especially, cows are still growing and must partition a certain proportion of nutrients consumed to growth, as well as milk production, often resulting in lighter calves [
39].
While the EB curves in the present study were based on weekly least square means, these curves were in agreement with modelled EB curves (polynomial regressions) for Holstein cows offered grass silage-based diets [
27,
40]. Although no differences were observed in the timing of nadir EB across lactations, nadir EB (MJ of ME/day) became more negative with parity. This was in agreement with earlier studies with nadir EB calculated by difference (energy intake minus energy expenditure) [
40,
41,
42], and also when it was calculated from EB curves based on body energy changes [
9,
41,
43]. In general, lactation curves increase to a peak as the cow seeks to meet the increasing nutrient needs of the calf, prior to the transition to a ‘solids’ diet, and the slow process of enforced weaning by the dam [
39,
44]. This corresponds with cows reaching the point of nadir EB in early lactation.
The milk fat and milk protein content followed normal lactation trends (providing the highest quality milk to the growing calf at a time when the calf was completely dependent on milk for growth), but with subtle differences between lactations and over time, which influenced FPR. The FPR ‘curves’ in the present study, which decreased from maximum values in early lactation and ‘plateaued’ around 10–13 WIM, were broadly similar to curves observed with Holstein cows in Germany [
27], Canada [
45], and Poland [
46]. For example, in the mean FPR curves presented by Satoła and Ptak [
46], and in the modeled FPR curves presented by Buttchereit et al. [
27], the maximum FPR occurred during the second WIM, irrespective of parity. Nevertheless, a visual examination of the FPR curves in the latter study revealed a high degree of variability during the first month of lactation, with this variation being also evident during early lactation in the present study, the time when energy deficit was most pronounced. The FPR in the present study mirrored the trend observed in EB, suggesting a relationship between these two parameters. This also agreed with the ‘inverted patterns’ observed by Buttchereit et al. [
27], where FPR and EB were modelled using polynomial regressions.
Nevertheless, across published studies, there is much variation in the ratio at which the curves plateaued, with curves plateauing at, approximately, 1.20 to 1.25 in the present study, between 1.05 (first and second lactation) and 1.15 (third and further lactation) in the study by Buttchereit et al. [
21], and 1.15 in the study by Satoła and Ptak [
46]. It is possible that this was due to genetic differences in the fat and protein content of the milk produced between cows in these studies. There were no obvious reasons for the trend towards a lower FPR curve with lactation two cows, although a close examination of the curves in
Figure 2 demonstrated that relatively minor differences in milk composition at a given point in time could result in quite substantial differences in FPR.
That cows in all lactations started to gain BW from approximately 4 WIM onwards, long before reaching positive EB, has been observed in earlier studies [
47,
48], with this likely due in part to meal-related ‘gut-fill’ [
9]. However, losses of BCS continued until, approximately, 6–10 WIM in multiparous cows and 12–14 WIM in primiparous cows; thus, following broadly similar, but mirror image, patterns to the FPR curves. In addition, Holstein dairy cows mobilize significant quantities of internal body fat reserves in early lactation, but this is not always fully reflected in BCS changes [
49,
50]. In the present study, the relatively modest degree of tissue mobilization (BW and BCS losses) might be partly related to the moderate BCS observed following calving (2.70, 2.49, 2.54, and 2.52 for lactations one, two, three, and ≥ four, respectively). Cows that have a higher BCS at parturition tend to lose more BCS postpartum than those with a low–moderate BCS [
12,
51,
52].
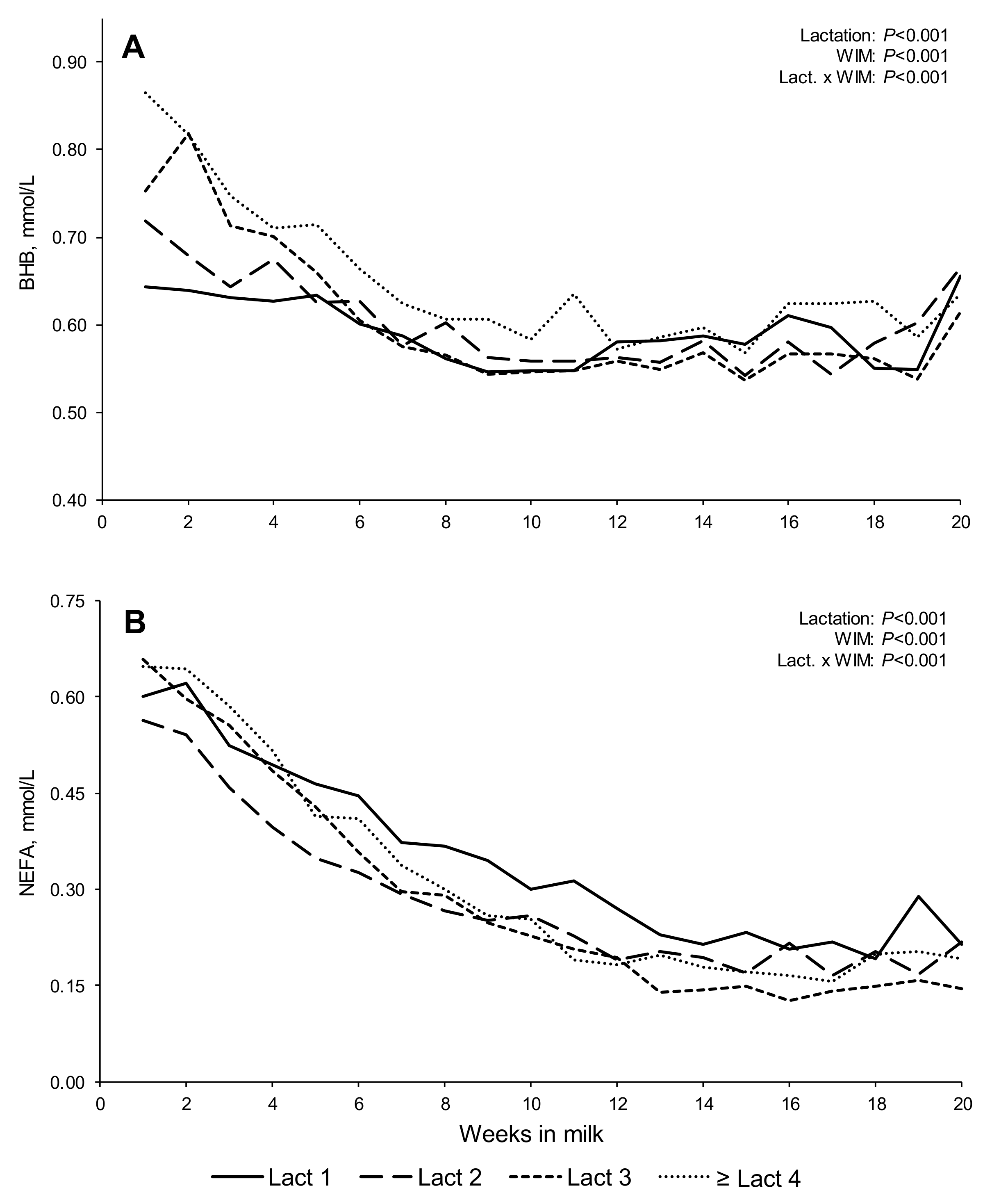
The decreasing concentrations of BHB and NEFA in blood plasma, both of which are known to be indicators of body fat mobilization [
15,
16,
53], were broadly aligned with the longitudinal weekly trends in BCS across lactations, although BHB concentrations appear to plateau earlier than NEFA concentrations. Maximum concentrations of BHB and NEFA during the first few WIM supported the occurrence of nadir EB within the second WIM for all lactations. The trend for lower weekly NEFA values early in lactation two, compared to other lactations, aligned with the trends in FPR curves and body tissue losses as described earlier.
4.2. Changes in FPR–EB Relationships with Stage of Lactation and Lactation Number
Uniquely, the present study allowed the relationships between FPR and EB to be examined at four wk. intervals during the first 20 WIM, and within each of lactations one, two, three, and > four. Irrespective of the lactation stage and parity, the partial correlations demonstrated that FPR was negatively related to EB, with the range of correlations observed broadly in agreement with those reported by Buttchereit et al. [
27] for primiparous cows (r = −0.15 to −0.43) and for multiparous cows by Grieve et al. [
25] (r = −0.36 to −0.74) and Reist et al. [
54] (r = 0.50), respectively. In the study by Buttchereit et al. [
27], phenotypical correlations between FPR and EB decreased as the lactation progressed (r = −0.42, −0.28, −0.20, at 35, 55, and 75 DIM, respectively), in agreement with genotypic correlations found later by the same authors [
55], and in a more recent study by Harder et al. [
56]. Thus, based on these correlations, FPR appears to be a stronger indicator of EB at the beginning of lactation when the cow experiences the greatest metabolic stress.
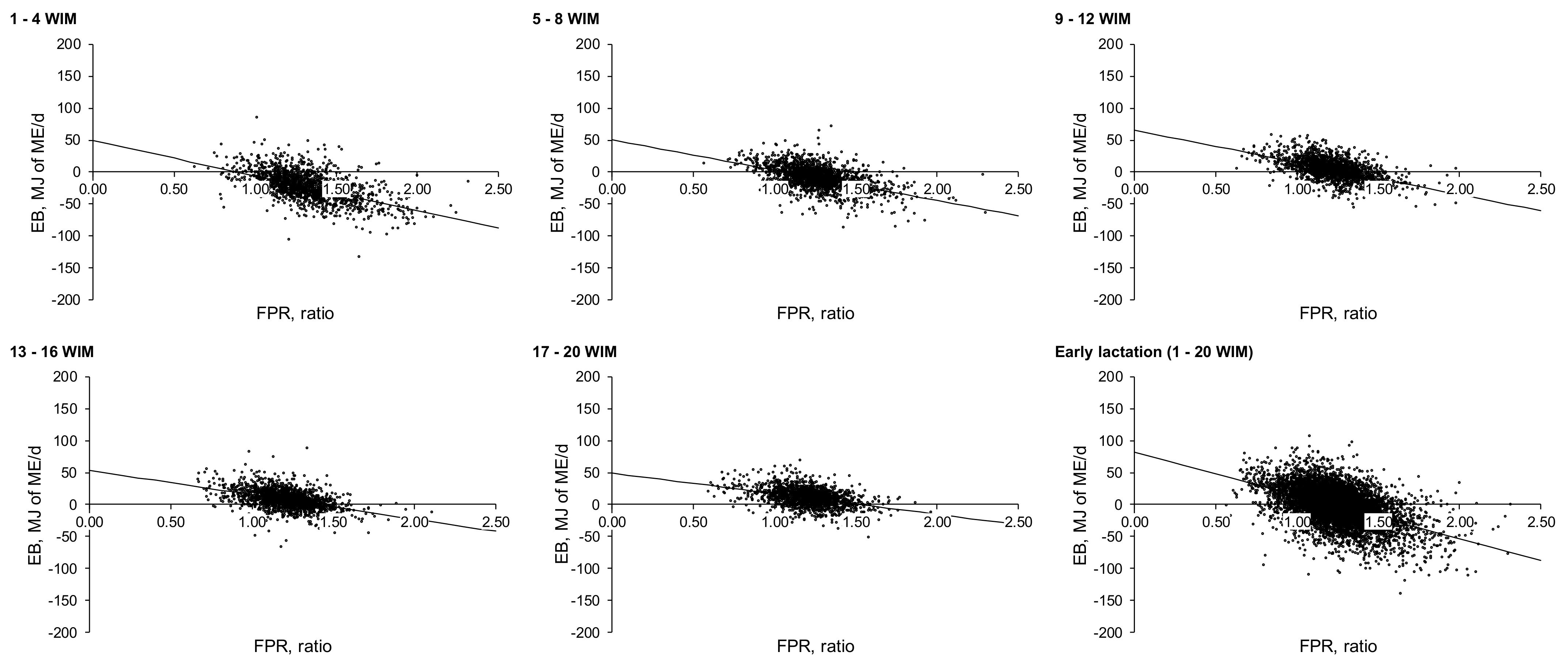
To the best of our knowledge, no previous study has examined FPR–EB relationships using a Linear Mixed Model approach evaluating between and within lactation responses as shown in the present study. The slopes of these regression models provide an estimate of the rate of decrease in EB per unit of increase in FPR. In the present study, when data from all 20 WIM were combined (slopes for the entire 20 WIM period for each lactation), the slope of the regression in each of lactations one to ≥ four was considerably more negative than for any of the individual four WIM periods within that lactation. An examination of the
Figure A1,
Figure A2,
Figure A3 and
Figure A4 (
Appendix B) highlights that this was caused by the vertical ‘stacking’ of data clusters from each four wk. period, creating an apparent response line that was very different from responses within individual four WIM periods. This demonstrates the danger of combining data for multiple weeks (1–20 WIM) for a parameter (EB) which changes over time as shown in the EB curves. Nevertheless, the examination of
Figure 1C, and
Figure A1,
Figure A2,
Figure A3 and
Figure A4 (
Appendix B), suggests that the increase in the slope with increasing parity (largely consistent when data from each four wks. period were compared across parity) was not primarily due to a greater range in FPR with increasing parity, but rather a greater range in EB values.
When data were compared across time points within any lactation, the
Figure A1,
Figure A2,
Figure A3 and
Figure A4 demonstrated that, in general, the data points showed a general ‘contraction’ from the ‘right’ to the ‘left’ as lactation progressed (i.e., a decreasing number of FPR points > 1.5). In addition, the overall dataset shifted upwards as EB increased, while the range of EB values encountered also appeared to decrease with the stage of lactation. However, the overall trends in slopes within each lactation were inconsistent, with the most negative slope occurring at 1– 4 WIM in lactations one and ≥ four, and at the WIM intervals of 5–8, 9–12 in lactations two and three, respectively. Reasons for the inconsistent trends across lactations are unclear, and our findings contrasted with the findings of Buttchereit et al. [
27], based on polynomial regressions of relationships between FPR and EB, that, irrespective of parity, the most NEB responses per unit of FPR were observed only within the first month of lactation.
The data presented in
Appendix B clearly highlight the wide range in individual cow FPR values, something that was not reflected in the relatively narrow range in mean values presented in
Figure 2C. Furthermore, the linear models suggest that the interaction between the stage of lactation and parity would need to be considered if attempting to use FPR as a single predictor (indicator) of EB in dairy cows during early lactation. Earlier studies indicated that differences in genetic correlations between FPR and EB as lactation progressed may be due to different genes regulating physiological mechanisms affecting EB at different times during early lactation [
42,
55,
57,
58].
4.3. Performance Indicators Associated with ‘Low’ and ‘High’ FPR Scores
While a ‘baseline’ FPR score between 1.0 and 1.5 is considered to represent normal physiological conditions in Holstein cows during early lactation [
59], an FPR outside of this range has been associated with the occurrence of metabolic and health disorders during early lactation. In particular, cows with an FPR greater > 1.5 are associated with an increased risk of ketosis, displaced abomasum, ovarian cysts, lameness, and mastitis [
24,
28,
59,
60], while those with an FPR of less than 1.0 have been found to have an increased risk of sub-acute ruminal acidosis (SARA) [
61,
62]. In the present study, the incidence of ‘High’ FPR scores decreased as lactation progressed (17.2, 12.9, 6.9, 4.4, and 4.2% at 1–4, 5–8, 9–12, 13–16, and 17–20 WIM, respectively; mean data across all lactations). A similar trend was observed in the study by Vlček et al. [
63], where the frequency of ‘High’ FPR scores (ketosis risk; >1.5 FPR) of 908 test-day records (208 Holstein cows) was found to be slightly higher during the first month of lactation (≈22%) and similar decreases observed with advanced lactation (≈10, 8, 4, and 3% for the second, third, fourth, and fifth month of lactation, respectively). In contrast, there was a small increase in the incidence of ‘Low’ FPR scores (<1.0) as lactation progressed (6.4, 7.2, 9.0, 9.7, and 8.8% at 1–4, 5–8, 9–12, 13–16, and 17–20 WIM, respectively; mean data across all lactations), with the incidence tending to reach a plateau around the time cows returned to positive EB. In contrast, in the study by Vlček et al. [
63], a more pronounced shift towards a greater incidence of ‘Low’ FPR scores (<1.0; risk of SARA) as lactation progressed was observed (≈11.0, 16.0, 19.0, 24.0, and 27.0% of cows at months 1–5 of lactations, respectively). Although the composition of diets offered in the latter study was not reported, these frequencies might suggest that the mean concentrate proportion in the diet offered may have been higher than in the present study. It is well known that an increased energy supply (via highly fermentable carbohydrates) in the diet of dairy cows is related to the increased risk of SARA, and this is reflected in low FPR values in milk [
64,
65].
The results presented in
Table 4,
Table 5,
Table 6 and
Table 7 allow an examination of how performance and blood parameters associated with each FPR score differed, with most of the traits presented either drivers of, or indicators of, EB. Across all parities and across all stages of lactation, EB showed a consistent decrease with an increasing FPR score, cows with an FPR score > 1.5 always having a significantly lower EB than those with an FPR score < 1.0. Both DMI and ECM are considered as primary factors influencing EB responses in dairy cows during early lactation, although their individual importance may vary according to parity and the stage of lactation. For example, studies by Hüttmann et al. [
58] and Krattenmacher et al. [
66] reported that EB was more genetically correlated to DMI than ECM across the first 180 DIM in primiparous cows. In contrast, Buttchereit et al. [
24] reported that genetic correlations between DMI and EB only became more important than those for ECM and EB during mid lactation (≥120 DIM). Within the present study, differences in EB between scores were due in part to differences in DMI, with a trend, although not always consistent, for cows with ‘High’ FPR scores to have lower DMI. In contrast, ECM appeared to be a more consistent driver of differences in EB between FPR scores, increasing significantly across each of the three FPR score bands during every 4 wk. period in all parities. Trends for decreased milk production associated with ‘Low’ FPR scores in the present study were in good agreement with results presented by Toni et al. [
28], who evaluated FPR at 7 DIM (test-day).
In agreement with the differences in EB between FPR scores, cows with an FPR score > 1.5 had a consistently greater BW loss, or lower BW gain, than cows with an FPR score < 1.0 throughout the first 12 WIM in all parities (except for 9–12 WIM in lactation ≥ four). In contrast, there were few differences in actual BCS between FPR score groups, and while this does not imply that changes in BCS did not follow a similar pattern for BW changes, it does suggest that cows with a different BCS were not more likely to be in one FPR score band than in another. This is perhaps surprising as there is ample evidence that cows with a high BCS are likely to have lower DMI and, subsequently, greater tissue loss in early lactation [
12,
49]. However, the present study has shown that lower DMI was not a consistent driver of NEB of cows having a ‘High’ FPR score. Despite this, Løvendahl et al. [
67] found that EB estimated from milk composition variables generally follows a time profile similar to EB estimated from BCS and BW.
The increasing concentrations of both BHB and NEFA (ln transformed) in blood metabolites at higher FPR scores in early lactation reflected the greater NEB and greater BW loss with higher FPR scores. These metabolites, produced from the mobilization of body tissue reserves to produce glycerol for use as an energy resource, are normally associated with high milk fat concentrations and, as such, ‘High’ FPR scores (>1.5) [
68]. Greater concentrations of each of these metabolites have been associated with an increased risk of subclinical ketosis, with risk thresholds for both plasma BHB and NEFA concentrations identified [
16,
17,
69]. For example, a NEFA threshold of 0.57 mEq/L was identified by Ospina et al. [
16] within the first 30 DIM for subclinical ketosis in North America. Untransformed NEFA concentrations in the present study at 1–4 WIM were 0.60, 0.52, 0.67, 0.68 mEq/L for lactations one, two, three, and ≥ four, respectively, greater than the threshold identified by Ospina et al. [
16] in all parities, except for the second lactation cows. However, none of these mean values were higher than 0.70 mEq/L during the first 20 DIM, a value recently identified as being indicative of excessive NEB in commercial dairy herds in the UK [
17]. Although BHB has been used as an indicator of NEB, high blood butyrate concentrations can reflect the rumen production of butyrate associated with an increased concentrate proportion in the diet [
70]. In contrast, NEFA is less influenced by diet and is considered as a better indicator of adipose tissue lipolysis [
10,
71]. Concentrations of NEFA and BHB have been reported not to be well correlated during the transition period [
72], which may partly explain discrepancies between these metabolites among FPR scores in the present study. The lowest incidences of ‘High’ FPR values observed with second lactation cows, especially during the first two WIM intervals, aligned with the smaller BW losses and lower NEFA concentrations when compared to cows in other lactations. Although data from the pre-partum period were not available in the present study, a smaller loss of BCS in second lactation cows during the entire transition period (including 3 wks. after parturition), compared to the other lactation groups, could partly explain these trends [
73].
While this paper established clear relationships between FPR and EB, it is important to examine the utility of FPR as a prediction tool of EB for individual cows. Partial correlations and linear regression models highlighted strong relationships between EB and FPR, while data presented in
Table 4,
Table 5,
Table 6 and
Table 7 show that cows within each FPR score differed in performance and EB. Nevertheless, a visual examination of individual cow data in the
Figure A1,
Figure A2,
Figure A3 and
Figure A4 (
Appendix B) did not suggest that FPR could be used to accurately predict the EB of individual cows. For example, during the first 8 WIM (across all parities), almost all observations (95.4%) with an FPR score > 1.5 were related to NEB (mean and S.D.; −56 ± 32.4 MJ of ME/day). However, during the same period, 79.2% of observations that had a ‘Normal’ FPR (between 1.0 and 1.5) were also in NEB (−33.3 ± 23.4 MJ of ME/day). Therefore, the large overlap in EB values between these two FPR score categories (1.0–1.5, and >1.5) confirms that FPR alone cannot accurately predict EB of individual cows. Furthermore, between 9 and 16 WIM, the majority of cows had an FPR score of between 1 and 1.5, while 47.9% of the observations with this score remained in NEB (−18 ± 16.6 MJ of ME/day). This supports the findings of previous studies that FPR had a limited predictive power and lacks precision to estimate the EB of individual cows [
18,
25,
67]. Rather, FPR may be a more useful tool at a herd level, providing information on the general energy status of a group of cows [
25,
74]. Novel approaches such as the definition of metabolic clusters [
75,
76,
77,
78] could be useful for characterizing individuals on the basis of their metabolic efficiency.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}