
Combining Analytical Approaches and Multiple Sources of Information to Improve Interpretation of Diagnostic Test Results for Tuberculosis in Wild Meerkats

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Laboratory Processing
2.3. Data Analyses
2.4. Sensitivity Estimates in the Presence, and Absence, of a Reference Test
2.5. Survival Analyses
2.6. Relationships with Time, Age, and Predicted Risk
3. Results
3.1. Sensitivity Estimates in the Presence, and Absence, of a Reference Test
3.2. Survival Analyses
3.3. Relationships with Time, Age, and Predicted Risk
4. Discussion
4.1. Variation in Results
4.2. Test Performance
4.3. Prognostic Indicators
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruel, A.V.D.; Cleemput, I.; Aertgeerts, B.; Ramaekers, D.; Buntinx, F. The evaluation of diagnostic tests: Evidence on technical and diagnostic accuracy, impact on patient outcome and cost-effectiveness is needed. J. Clin. Epidemiol. 2007, 60, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, D.U. (Ed.) Interpretation of diagnostic tests. In Veterinary Epidemiology: An Introduction; Wiley-Blackwell: Oxford, UK, 2002. [Google Scholar]
- Dohoo, I.; Martin, W.; Stryhn, H. Veterinary Epidemiological Research; AVC Inc.: Vancouver, BC, Canada, 2003. [Google Scholar]
- Stallknecht, D.E. Impediments to wildlife disease surveillance, research, and diagnostics. In Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission; Childs, J.E., Mackenzie, J.S., Richt, J.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 445–461. [Google Scholar]
- Pollock, J.; Neill, S. Mycobacterium bovis infection and tuberculosis in cattle. Vet. J. 2002, 163, 115–127. [Google Scholar] [CrossRef]
- Vordermeier, M.; Goodchild, A.; Clifton-Hadley, R.; De La Rua, R. The interferon-gamma field trial: Background, principles and progress. Vet. Rec. 2004, 155, 37–38. [Google Scholar]
- Welsh, M.D.; Cunningham, R.T.; Corbett, D.M.; Girvin, R.M.; McNair, J.; Skuce, R.A.; Bryson, D.G.; Pollock, J.M. Influence of pathological progression on the balance between cellular and humoral immune responses in bovine tuberculosis. Immunology 2005, 114, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ritacco, V.; Lopez, B.; De Kantor, I.; Barrera, L.; Errico, F.; Nader, A. Reciprocal cellular and humoral immune responses in bovine tuberculosis. Res. Vet. Sci. 1991, 50, 365–367. [Google Scholar] [CrossRef]
- Parsons, S.D.; Drewe, J.A.; Van Pittius, N.C.G.; Warren, R.M.; Van Helden, P.D. Novel cause of tuberculosis in Meerkats, South Africa. Emerg. Infect. Dis. 2013, 19, 2004–2007. [Google Scholar] [CrossRef] [PubMed]
- Drewe, J.; Foote, A.; Sutcliffe, R.; Pearce, G. Pathology of Mycobacterium bovis infection in Wild Meerkats (Suricata suricatta). J. Comp. Pathol. 2009, 140, 12–24. [Google Scholar] [CrossRef]
- Drewe, J.A. Who infects whom? Social networks and tuberculosis transmission in wild meerkats. Proc. R. Soc. B Boil. Sci. 2009, 277, 633–642. [Google Scholar] [CrossRef]
- Patterson, S.; Drewe, J.A.; Pfeiffer, D.U.; Clutton-Brock, T.H. Social and environmental factors affect tuberculosis related mortality in wild meerkats. J. Anim. Ecol. 2017, 86, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Clutton-Brock, T.H.; Gaynor, D.; Kansky, R.; MacColl, A.; McIlrath, G.; Chadwick, P.; Brotherton, P.N.M.; O’Riain, M.J.; Manser, M.B.; Skinner, J.D. Costs of cooperative behaviour in suricates (Suricata suricatta). Proc. R. Soc. B Boil. Sci. 1998, 265, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Clutton-Brock, T.H.; Brotherton, P.N.M.; Russell, A.F.; O’Riain, M.J.; Gaynor, D.; Kansky, R.; Griffin, A.; Manser, M.; Sharpe, L.; McIlrath, G.M.; et al. Cooperation, control, and concession in Meerkat groups. Science 2001, 291, 478–481. [Google Scholar] [CrossRef] [Green Version]
- Drewe, J.A.; Dean, G.S.; Michel, A.; Pearce, G. Accuracy of Three diagnostic tests for determining Mycobacterium bovis infection status in live-sampled wild Meerkats (Suricata Suricatta). J. Vet. Diagn. Investig. 2009, 21, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Clarke, C.; Patterson, S.J.; Drewe, J.A.; van Helden, P.D.; Miller, M.A.; Parsons, S.D.C. Development and evaluation of a diagnostic cytokine-release assay for Mycobacterium suricattae infection in meerkats (Suricata suricatta). BMC Vet. Res. 2016, 13, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwald, R.; Lyashchenko, O.; Esfandiari, J.; Miller, M.; Mikota, S.; Olsen, J.H.; Ball, R.; Dumonceaux, G.; Schmitt, D.; Moller, T.; et al. Highly accurate antibody assays for early and rapid detection of tuberculosis in african and asian elephants. Clin. Vaccine Immunol. 2009, 16, 605–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Rua-Domenech, R.; Goodchild, A.; Vordermeier, H.; Hewinson, R.; Christiansen, K.; Clifton-Hadley, R. Ante mortem diagnosis of tuberculosis in cattle: A review of the tuberculin tests, γ-interferon assay and other ancillary diagnostic techniques. Res. Vet. Sci. 2006, 81, 190–210. [Google Scholar] [CrossRef]
- Warren, R.M.; Van Pittius, N.C.G.; Barnard, M.; Hesseling, A.; Engelke, E.; De Kock, M.; Gutierrez, M.C.; Chege, G.K.; Victor, T.C.; Hoal, E.; et al. Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference. Int. J. Tuberc. Lung Dis. 2006, 10, 818–822. [Google Scholar] [PubMed]
- Branscum, A.; Gardner, I.; Johnson, W. Estimation of diagnostic-test sensitivity and specificity through Bayesian modeling. Prev. Vet. Med. 2005, 68, 145–163. [Google Scholar] [CrossRef]
- Praud, A.; Champion, J.-L.; Corde, Y.; Drapeau, A.; Meyer, L.; Garin-Bastuji, B. Assessment of the diagnostic sensitivity and specificity of an indirect ELISA kit for the diagnosis of Brucella ovis infection in rams. BMC Vet. Res. 2012, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Enøe, C.; Georgiadis, M.P.; Johnson, W.O. Estimation of sensitivity and specificity of diagnostic tests and disease prevalence when the true disease state is unknown. Prev. Vet. Med. 2000, 45, 61–81. [Google Scholar] [CrossRef]
- Joseph, L.; Gyorkos, T.W.; Coupal, L. Bayesian estimation of disease prevalence and the parameters of diagnostic tests in the absence of a gold standard. Am. J. Epidemiol. 1995, 141, 263–272. [Google Scholar] [CrossRef]
- McHugh, M.L. Interrater reliability: The kappa statistic. Biochem. Med. 2012, 22, 276–282. [Google Scholar] [CrossRef]
- Enderlein, G.; Cox, D.R.; Oakes, D. Analysis of Survival Data; Chapman and Hall: London, UK, 1984. [Google Scholar] [CrossRef]
- Therneau, T.; Grambsch, P.M. Modelling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Hougaard, P. Frailty models for survival data. Lifetime Data Anal. 1995, 1, 255–273. [Google Scholar] [CrossRef]
- Lunn, D.J.; Thomas, A.; Best, N.; Spiegelhalter, D. WinBUGS—A Bayesian modelling framework: Concepts, structure, and extensibility. Stat. Comput. 2000, 10, 325–337. [Google Scholar] [CrossRef]
- Sturtz, S.; Ligges, U.; Gelman, A. R2WinBUGS: A package for Running WinBUGS from R. J. Stat. Softw. 2005, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Rubin, D.B. Bayesian Data Analysis; Texts in Statistical Science Series; Chapman & Hall/CRC: Boca Raton, FL, USA, 2004. [Google Scholar]
- van Dijk, P.C.; Jager, K.J.; Zwinderman, A.H.; Zoccali, C.; Dekker, F.W. The analysis of survival data in nephrology: Basic concepts and methods of Cox regression. Kidney Int. 2008, 74, 705–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, S.D.C.; McGill, K.; Doyle, M.B.; Goosen, W.J.; Van Helden, P.D.; Gormley, E. Antigen-specific IP-10 release is a sensitive Biomarker of Mycobacterium bovis infection in cattle. PLoS ONE 2016, 11, e0155440. [Google Scholar] [CrossRef] [PubMed]
- Ruhwald, M.; Domínguez, J.; Latorre, I.; Losi, M.; Richeldi, L.; Pasticci, M.B.; Mazzolla, R.; Goletti, D.; Butera, O.; Bruchfeld, J.; et al. A multicentre evaluation of the accuracy and performance of IP-10 for the diagnosis of infection with M. tuberculosis. Tuberculosis. 2011, 91, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Cousins, D.V.; Florisson, N. A review of tests available for use in the diagnosis of tuberculosis in non-bovine species. Rev. Sci. Tech. (Int. Off. Epizoot.) 2005, 24, 1039–1059. [Google Scholar] [CrossRef]
- Chambers, M.A. Review of the diagnosis of Tuberculosis in non-bovid wildlife species using immunological methods—An update of published work since 2009. Transbound. Emerg. Dis. 2013, 60, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Greenwald, R.; Esfandiari, J.; Lesellier, S.; Houghton, R.; Pollock, J.; Aagaard, C.; Andersen, P.; Hewinson, R.; Chambers, M.; Lyashchenko, K. Improved serodetection of Mycobacterium bovis infection in badgers (Meles meles) using multiantigen test formats. Diagn. Microbiol. Infect. Dis. 2003, 46, 197–203. [Google Scholar] [CrossRef]
- Lyashchenko, K.P.; Greenwald, R.; Esfandiari, J.; O’Brien, D.; Schmitt, S.M.; Palmer, M.V.; Waters, W.R. Rapid detection of serum antibody by Dual-Path platform VetTB assay in white-tailed deer infected with Mycobacterium bovis. Clin. Vaccine Immunol. 2013, 20, 907–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, S.; Holder, T.; Clifford, D.; Dexter, I.; Brewer, J.; Smith, N.; Waring, L.; Crawshaw, T.; Gillgan, S.; Lyashchenko, K.; et al. Evaluation of gamma interferon and antibody Tuberculosis tests in Alpacas. Clin. Vaccine Immunol. 2012, 19, 1677–1683. [Google Scholar] [CrossRef] [PubMed]
- Buzdugan, S.N.; Chambers, M.A.; Delahay, R.J.; Drewe, J.A. Diagnosis of tuberculosis in groups of badgers: An exploration of the impact of trapping efficiency, infection prevalence and the use of multiple tests. Epidemiol. Infect. 2016, 144, 1717–1727. [Google Scholar] [CrossRef] [Green Version]
- Kerkhoff, A.D.; Cattamanchi, A.; Muyoyeta, M.; Denkinger, C.M.; Dowdy, D.W. Validating novel diagnostic assays for tuberculosis in the context of existing tools. Lancet Glob. Health 2021, 9, e1209. [Google Scholar] [CrossRef]
- Abbate, J.M.; Arfuso, F.; Iaria, C.; Arestia, G.; Lanteri, G. Prevalence of Bovine Tuberculosis in slaughtered Cattle in Sicily, Southern Italy. Animal 2020, 10, 1473. [Google Scholar] [CrossRef]
- Pascual-Linaza, A.V.; Gordon, A.W.; Stringer, L.A.; Menzies, F.D. Efficiency of slaughterhouse surveillance for the detection of bovine tuberculosis in cattle in Northern Ireland. Epidemiol. Infect. 2017, 145, 995–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himsworth, C.G.; Elkin, B.T.; Nishi, J.S.; Epp, T.; Lyashchenko, K.P.; Surujballi, O.; Turcotte, C.; Esfandiari, J.; Greenwald, R.; Leighton, F.A. Comparison of test performance and evaluation of novel immunoassays for Tuberculosis in a captive herd of wood bison naturally infected with Mycobacterium bovis. J. Wildl. Dis. 2010, 46, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Aim of the Analysis | Approach Used | Inclusion Criteria | Time Period | References |
|---|---|---|---|---|
| 1. To determine the ability of each testing regime to correctly identify infected animals (test sensitivity) | Comparison with a reference test | All animals from which M. suricattae was confirmed on culture of post-mortem material, at any point during the study. All samples collected prior to death were included. | September 2014-September 2016 inclusive (Full) | [3] |
| 2. To determine each testing regime’s ability to correctly identify animals as infected (sensitivity) or uninfected (specificity) | Latent Class Analysis | The first sample collected for each individual was used. The time-period was limited to minimise the impact of changes over time on prevalence. | September 2014–March 2015 inclusive | [20,21,22,23] |
| 3. To quantify the agreement between the results from different tests | Calculation of the kappa statistic | All sampling events where there was more than one test were used. | Full | [24] |
| 4. To investigate whether test results were predictive of mortality | Survival analysis of time to death | All samples collected over the study period from wild individuals were used. | Full | [25] |
| 5. To investigate whether test results were predictive of clinical signs of disease | Survival analysis of time to showing clinical signs | All samples collected over the study period from wild individuals were used. | Full | [25] |
| 6. To investigate whether individuals’ test results fluctuate over time | Analysis of consistency over time | All individuals sampled on more than one occasion were included. | Full | - |
| 7. To investigate whether test result is influenced by age | Analysis for association with age | All samples from individuals with a known date of birth were included. | Full | - |
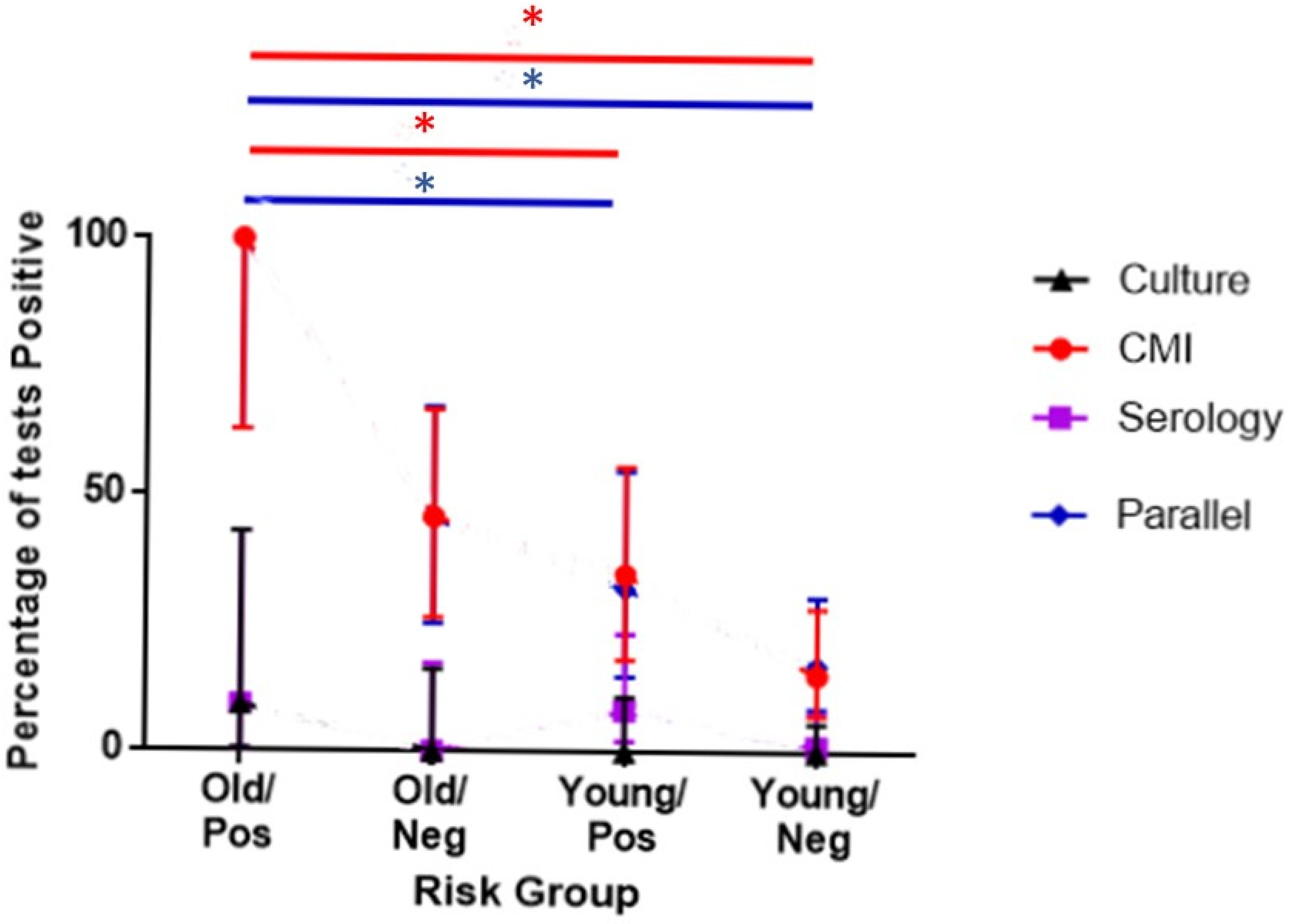
| 8. To Investigate whether test results were detecting infection more often in the most susceptible animals | Analysis for association with risk | Only the first sample from individuals with a known date of birth were included. The time-period was limited to minimise the impact of changes over time. | September 2014–March 2015 inclusive | [12] |
| Individual Test Interpretations | Multiple Test Interpretations | |||||
|---|---|---|---|---|---|---|
| Time before Euthanasia (days) | n | Serology (DPP) (%, 95CI *) | Cell-Mediated Immunity (IPRA) (%, 95CI *) | Tracheal Wash Culture (%, 95CI *) | Parallel (%, 95CI *) | Ever Positive (%, 95CI *) |
| 0 | 23 | 3/22 (13, 3–35) | 10/22 (45, 25–67) | 3/23 (13, 3–35) | 13/22 (59, 37–79) | 18/22 (82, 59–95) |
| 1–90 | 10 | 0/10 (0, 0–35) | 4/10 (40, 14–73) | 0/10 (0, 0–35) | 4/9 (44, 15–77) | 6/9 (67, 31–91) |
| 91–180 | 7 | 0/7 (0, 0–44) | 4/7 (57, 20–88) | 0/7 (0, 0–44) | 4/7 (57, 20–88) | 5/7 (71, 30–95) |
| 181–365 | 11 | 0/11 (0, 0–32) | 1/8 (13, 1–53) | 0/11 (0, 0–32) | 1/8 (13, 1–53) | 2/8 (25, 5–64) |
| >365 | 11 | 0/11 (0, 0–32) | 1/8 (13, 1–53) | 0/11 (0, 0–32) | 1/8 (13, 1–53) | 1/8 (13, 1–53) |
| Serology (DPP) | Cell-Mediated Immunity (IPRA) | Tracheal Wash Culture | Parallel Interpretation | |
|---|---|---|---|---|
| Sensitivity (%, Credible interval) | 12.0 (4.0–33.6) | 58.6 (35.1–80.6) | 2.2 (0.1–10.9) | 65.7 (42.7–84.7) |
| Specificity (%, Credible interval) | 97.9 (92.8–99.9) | 85.0 (68.4–99.4) | 99.2 (95.7–99.9) | 81.9 (65.2–96.7) |
| Model a—All Animals (n = 126) | |||||
| Univariable | Hazard Ratio | 95% Confidence Interval | p-Value | ||
| Dominance | No | 0.966 | |||
| Yes | 0.99 | 0.64–1.53 | |||
| Sex | Female | 0.813 | |||
| Male | 1.05 | 0.711.54 | |||
| Age | <6 months | 0.678 | |||
| 6–12 months | 1.03 | 0.55–1.92 | |||
| >12 months | 0.85 | 0.49–1.45 | |||
| Serology | Neg a | 0.039 | |||
| Pos b | 2.44 | 1.04–5.70 | |||
| Cell-Mediated immunity | Neg a | 0.107 | |||
| Pos b | 1.45 | 0.92–2.28 | |||
| Culture | Neg a | 0.013 | |||
| Pos b | 14.84 | 1.76–125.5 | |||
| Parallel | Neg a | 0.010 | |||
| Pos b | 1.75 | 1.14–2.67 | |||
| Multivariable | Hazard Ratio | 95% Confidence Interval | p-value | Likelihood ratio for model, p value | |
| Serology + Frailty Sampling group | Neg a | <0.0001 | |||
| Pos b | 1.29 | 0.51–3.31 | 0.59 | ||
| Variance of frailty term | 1.126 | ||||
| Cell-Mediated immunity + Frailty Social group | Neg a | <0.0001 | |||
| Pos b | 1.10 | 0.65–1.87 | 0.72 | ||
| Variance of frailty term | 1.939 | ||||
| Culture + Frailty Social group | Neg a | <0.0001 | |||
| Pos b | 1.92 | 0.22–16.60 | 0.55 | ||
| Variance of frailty term | 1.075 | ||||
| Parallel + Frailty Social group | Neg a | <0.0001 | |||
| Pos b | 1.30 | 0.80–2.11 | 0.29 | ||
| Variance of frailty term | 1.028 | ||||
| Model b–Excluding euthanised animals (n = 112) | |||||
| Univariable | Hazard Ratio | 95% Confidence Interval | p-value | ||
| Dominance | No | 0.780 | |||
| Yes | 1.06 | 0.69–1.65 | |||
| Sex | F | 0.820 | |||
| M | 1.05 | 0.70–1.56 | |||
| Age | <6 months | 0.897 | |||
| 6–12 months | 0.93 | 0.48–1.79 | |||
| >12 months | 0.88 | 0.51–1.53 | |||
| Serology | Neg a | 0.116 | |||
| Pos b | 2.09 | 0.83–5.27 | |||
| CMI | Neg a | 0.09 | |||
| Pos b | 1.48 | 0.93–2.34 | |||
| Culture | Neg a | 0.008 | |||
| Pos b | 18.66 | 2.13–163.60 | |||
| Parallel | Neg a | 0.011 | |||
| Pos b | 1.75 | 1.14–2.68 | |||
| Multivariable | Hazard Ratio | 95% Confidence Interval | p-value | Likelihood ratio for model, p value | |
| Serology + Frailty Sampling group | Neg a | <0.0001 | |||
| Pos b | 1.04 | 0.38–2.86 | 0.94 | ||
| Variance of frailty term | 1.126 | ||||
| CMI + Frailty Social group | Neg a | <0.0001 | |||
| Pos b | 1.17 | 0.68–1.99 | 0.57 | ||
| Variance of frailty term | 1.809 | ||||
| Culture + Frailty Social group | Neg a | <0.0001 | |||
| Pos b | 2.37 | 0.26–21.39 | 0.44 | ||
| Variance of frailty term | 1.013 | ||||
| Parallel + Frailty Social group | Neg a | <0.0001 | |||
| Pos b | 1.33 | 0.81–2.17 | 0.26 | ||
| Variance of frailty term | 0.972 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patterson, S.J.; Clarke, C.; Clutton-Brock, T.H.; Miller, M.A.; Parsons, S.D.C.; Pfeiffer, D.U.; Vergne, T.; Drewe, J.A. Combining Analytical Approaches and Multiple Sources of Information to Improve Interpretation of Diagnostic Test Results for Tuberculosis in Wild Meerkats. Animals 2021, 11, 3453. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11123453
Patterson SJ, Clarke C, Clutton-Brock TH, Miller MA, Parsons SDC, Pfeiffer DU, Vergne T, Drewe JA. Combining Analytical Approaches and Multiple Sources of Information to Improve Interpretation of Diagnostic Test Results for Tuberculosis in Wild Meerkats. Animals. 2021; 11(12):3453. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11123453
Chicago/Turabian StylePatterson, Stuart J., Charlene Clarke, Tim H. Clutton-Brock, Michele A. Miller, Sven D. C. Parsons, Dirk U. Pfeiffer, Timothée Vergne, and Julian A. Drewe. 2021. "Combining Analytical Approaches and Multiple Sources of Information to Improve Interpretation of Diagnostic Test Results for Tuberculosis in Wild Meerkats" Animals 11, no. 12: 3453. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11123453