
Assessment of Lung Disease in Finishing Pigs at Slaughter: Pulmonary Lesions and Implications on Productivity Parameters

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Pathologic and Immunohistochemical Examinations
2.2.1. Gross Examinations at Slaughterhouse
2.2.2. Histopathologic Examination
2.2.3. Immunohistochemical Analysis
2.3. Serologic Analysis
2.3.1. Blood Collection and Sera Preparation
2.3.2. ELISA Tests
2.4. Evaluation of Productive Parameters
2.5. Statistical Analyses
3. Results
3.1. Pathologic and Immunohistochemical Examinations
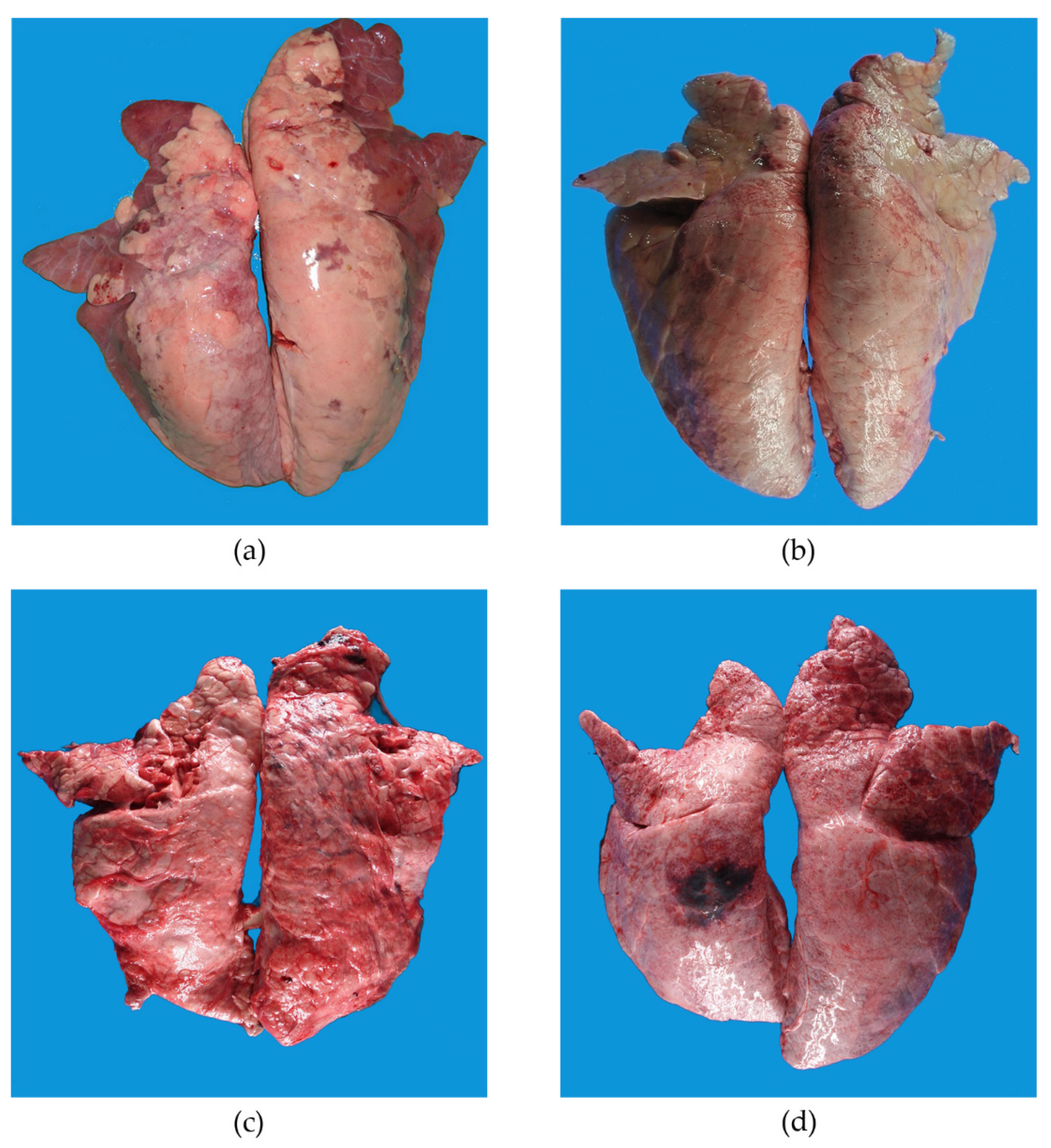
3.1.1. Gross Examinations at Slaughterhouse
3.1.2. Histopathologic Examination
3.1.3. Immunohistochemical Analysis
3.2. Serologic Analysis
3.3. Evaluation of Productive Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. Pork: Information on Pork Production, Legal Bases, Market Monitoring, Trade, Carcass Classification, Price Reporting, Regulation of Supply of PDO/PGI Ham. Available online: https://ec.europa.eu/info/food-farming-fisheries/animals-and-animal-products/animal-products/pork_en#introduction (accessed on 17 November 2021).
- Ministerio de Agricultura. Pesca y Alimentación Encuesta de Sacrificio de Ganado. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/ganaderia/encuestas-sacrificio-ganado/ (accessed on 17 November 2021).
- Maes, D.; Sibila, M.; Kuhnert, P.; Segalés, J.; Haesebrouck, F.; Pieters, M. Update on Mycoplasma hyopneumoniae infections in pigs: Knowledge gaps for improved disease control. Transbound. Emerg. Dis. 2018, 65, 110–124. [Google Scholar] [CrossRef] [Green Version]
- Karabasil, N.; Cobanovic, N.; Vucicevic, I.; Stajkovic, S.; Becskei, Z.; Forgách, P.; Aleksic-Kovacevic, S. Association of the severity of lung lesions with carcass and meat quality in slaughter pigs. Acta Vet. Hung. 2017, 65, 354–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraz, M.E.S.; Almeida, H.M.S.; Storino, G.Y.; Sonálio, K.; Souza, M.R.; Moura, C.A.A.; Costa, W.M.T.; Lunardi, L.; Linhares, D.C.L.; de Oliveira, L.G. Lung consolidation caused by Mycoplasma hyopneumoniae has a negative effect on productive performance and economic revenue in finishing pigs. Prev. Vet. Med. 2020, 182, 105091. [Google Scholar] [CrossRef]
- Sørensen, V.; Jorsal, S.E.L.; Mousing, J. Diseases of the respiratory system. In Diseases of Swine; Straw, B.E., Zimmerman, J.J., D’Allaire, S., Taylor, D.J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 149–177. ISBN 978-0-8138-1703-3. [Google Scholar]
- Brockmeier, S.L.; Halbur, P.G.; Thacker, E.L. Porcine respiratory disease complex. In Polymicrobial Diseases; ASM Press: Washington, DC, USA, 2002; pp. 231–258. [Google Scholar]
- Fraile, L.; Alegre, A.; López-Jiménez, R.; Nofrarías, M.; Segalés, J. Risk factors associated with pleuritis and cranio-ventral pulmonary consolidation in slaughter-aged pigs. Vet. J. 2010, 184, 326–333. [Google Scholar] [CrossRef]
- Meyns, T.; van Steelant, J.; Rolly, E.; Dewulf, J.; Haesebrouck, F.; Maes, D. A cross-sectional study of risk factors associated with pulmonary lesions in pigs at slaughter. Vet. J. 2011, 187, 388–392. [Google Scholar] [CrossRef]
- Merialdi, G.; Dottori, M.; Bonilauri, P.; Luppi, A.; Gozio, S.; Pozzi, P.; Spaggiari, B.; Martelli, P. Survey of pleuritis and pulmonary lesions in pigs at abattoir with a focus on the extent of the condition and herd risk factors. Vet. J. 2012, 193, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.S.; Pors, S.E.; Jensen, H.E.; Bille-Hansen, V.; Bisgaard, M.; Flachs, E.M.; Nielsen, O.L. An investigation of the pathology and pathogens associated with porcine respiratory disease complex in Denmark. J. Comp. Pathol. 2010, 143, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Fablet, C.; Marois, C.; Dorenlor, V.; Eono, F.; Eveno, E.; Jolly, J.P.; le Devendec, L.; Kobisch, M.; Madec, F.; Rose, N. Bacterial pathogens associated with lung lesions in slaughter pigs from 125 herds. Res. Vet. Sci. 2012, 93, 627–630. [Google Scholar] [CrossRef]
- Ostanello, F.; Dottori, M.; Gusmara, C.; Leotti, G.; Sala, V. Pneumonia disease assessment using a slaughterhouse lung-scoring method. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 2007, 54, 70–75. [Google Scholar] [CrossRef]
- Garcia-Morante, B.; Segalés, J.; Fraile, L.; Pérez de Rozas, A.; Maiti, H.; Coll, T.; Sibila, M. Assessment of Mycoplasma hyopneumoniae-induced Pneumonia using Different Lung Lesion Scoring Systems: A Comparative Review. J. Comp. Pathol. 2016, 154, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Van Alstine, W.G. Respiratory system. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 348–362. ISBN 978-0-8138-2267-9. [Google Scholar]
- Madec, F.; Kobisch, M. Bilan lésionnel des poumons de porcs charcutiers à l’abattoir. Journ. Rech. Porc. 1982, 14, 405–412. [Google Scholar]
- Christensen, G.; Sørensen, V.; Mousing, J. Diseases of the respiratory system. In Diseases of Swine; Straw, B.E., D’Allaire, S., Mengeling, W.L., Taylor, D.J., Eds.; Iowa State University Press: Ames, IA, USA, 1999; pp. 913–939. ISBN 0-8138-0338-1. [Google Scholar]
- Piffer, I.A.; Brito, J.R.F. Descricao de um Modelo para Avaliacao e Quantificacao de Lesoes Pulmonares de Suinos e Formulacao de um Indice para Classificacao de Rebanhos; Embrapa: Brasília, Brazil, 1991. [Google Scholar]
- Fablet, C.; Marois-Créhan, C.; Simon, G.; Grasland, B.; Jestin, A.; Kobisch, M.; Madec, F.; Rose, N. Infectious agents associated with respiratory diseases in 125 farrow-to-finish pig herds: A cross-sectional study. Vet. Microbiol. 2012, 157, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Thacker, E.L.; Minion, F.C. Mycoplasmosis. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 779–797. ISBN 978-0-8138-2267-9. [Google Scholar]
- Thacker, E.L.; Thanawongnuwech, R. Porcine respiratory disease complex. Thai J. Vet. Med. 2002, 32, 126–134. [Google Scholar]
- Livingston, C.W.; Stair, E.L.; Underdahl, N.R.; Mebus, C.A. Pathogenesis of mycoplasmal pneumonia in swine. Am. J. Vet. Res. 1972, 33, 2249–2258. [Google Scholar]
- Pallarés, F.; Añón, J.; Rodríguez-Gómez, I.; Gómez-Laguna, J.; Fabré, R.; Sánchez-Carvajal, J.; Ruedas-Torres, I.; Carrasco, L. Prevalence of mycoplasma-like lung lesions in pigs from commercial farms from Spain and Portugal. Porc. Health Manag. 2021, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jirawattanapong, P.; Stockhofe-Zurwieden, N.; van Leengoed, L.; Wisselink, H.; Raymakers, R.; Cruijsen, T.; van der Peet-Schwering, C.; Nielen, M.; van Nes, A. Pleuritis in slaughter pigs: Relations between lung lesions and bacteriology in 10 herds with high pleuritis. Res. Vet. Sci. 2010, 88, 11–15. [Google Scholar] [CrossRef]
- López-Lorenzo, G.; Prieto, A.; López-Novo, C.; Díaz, P.; López, C.M.; Morrondo, P.; Fernández, G.; Díaz-Cao, J.M. Efficacy of Two Commercial Ready-To-Use PCV2 and Mycoplasma hyopneumoniae Vaccines under Field Conditions. Animals 2021, 11, 1553. [Google Scholar] [CrossRef] [PubMed]
- Maes, D.; Verdonck, M.; Deluyker, H.; DeKruif, A. Enzootic pneumonia in pigs. Vet. Q. 1996, 18, 104–109. [Google Scholar] [CrossRef]
- Andrada, M. Estudio Etiopatogénico de la Neumonía Enzoótica Porcina (NEP). Ph.D. Thesis, University of Las Palmas de Gran Canaria, Las Palmas de Gran Canaria, Spain, 2001. [Google Scholar]
- Sørensen, V.; Ahrens, P.; Barfod, K.; Feenstra, A.A.; Feld, N.C.; Friis, N.F.; Bille-Hansen, V.; Jensen, N.E.; Pedersen, M.W. Mycoplasma hyopneumoniae infection in pigs: Duration of the disease and evaluation of four diagnostic assays. Vet. Microbiol. 1997, 54, 23–34. [Google Scholar] [CrossRef]
- Thacker, E.; Thacker, B.J.; Janke, B.H. Interaction between Mycoplasma hyopneumoniae and swine influenza virus. J. Clin. Microbiol. 2001, 39, 2525–2530. [Google Scholar] [CrossRef] [Green Version]
- Almeida, H.M.S.; Mechler-Dreibi, M.L.; Sonálio, K.; Ferraz, M.E.S.; Storino, G.Y.; Barbosa, F.O.; Maes, D.; Montassier, H.J.; de Oliveira, L.G. Cytokine expression and Mycoplasma hyopneumoniae burden in the development of lung lesions in experimentally inoculated pigs. Vet. Microbiol. 2020, 244, 108647. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, H.; Quesada, Ó.; Assunçao, P.; Castro, A.; Rodríguez, F. Cytokine expression in porcine lungs experimentally infected with Mycoplasma hyopneumoniae. Vet. Immunol. Immunopathol. 2006, 109, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Chae, C. Porcine respiratory disease complex: Interaction of vaccination and porcine circovirus type 2, porcine reproductive and respiratory syndrome virus, and Mycoplasma hyopneumoniae. Vet. J. 2016, 212, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Thacker, E.L.; Halbur, P.G.; Ross, R.F.; Thanawongnuwech, R.; Thacker, B.J. Mycoplasma hyopneumoniae potentiation of porcine reproductive and respiratory syndrome virus-induced pneumonia. J. Clin. Microbiol. 1999, 37, 620–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanawongnuwech, R.; Thacker, B.; Halbur, P.; Thacker, E.L. Increased production of proinflammatory cytokines following infection with porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae. Clin. Diagn. Lab. Immunol. 2004, 11, 901–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaniyi, M.O.; Awoyomi, O.J.; Akinniyi, O.; Adebiyi, A.A.; Alaka, O.O.; Ajayi, O.L.; Jubril, A.J.; Jarikre, T.A.; Emikpe, B.O. Retrospective study of swine respiratory diseases in Ogun and Oyo States, Nigeria: Immunohistochemical detection of Mycoplasma hyopneumoniae. Sokoto J. Vet. Sci. 2020, 18, 72–82. [Google Scholar] [CrossRef]
- Sibila, M.; Nofrarías, M.; López-Soria, S.; Segalés, J.; Riera, P.; Llopart, D.; Calsamiglia, M. Exploratory field study on Mycoplasma hyopneumoniae infection in suckling pigs. Vet. Microbiol. 2007, 121, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Redondo, E.; Masot, A.J.; Fernández, A.; Gázquez, A. Histopathological and Immunohistochemical Findings in the Lungs of Pigs Infected Experimentally with Mycoplasma hyopneumoniae. J. Comp. Pathol. 2009, 140, 260–270. [Google Scholar] [CrossRef]
- Hansen, M.S.; Pors, S.E.; Bille-Hansen, V.; Kjerulff, S.K.J.; Nielsen, O.L. Occurrence and tissue distribution of porcine circovirus type 2 identified by immunohistochemistry in Danish finishing pigs at slaughter. J. Comp. Pathol. 2010, 142, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Haruna, J.; Hanna, P.; Hurnik, D.; Ikede, B.; Miller, L.; Yason, C. The role of immunostimulation in the development of postweaning multisystemic wasting syndrome in pigs under field conditions. Can. J. Vet. Res. 2006, 70, 269–276. [Google Scholar] [PubMed]
- Halbur, P.G.; Paul, P.S.; Frey, M.L.; Landgraf, J.; Eernisse, K.; Meng, X.-J.; Andrews, J.J.; Lum, M.A.; Rathje, J.A. Comparison of the antigen distribution of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 1996, 33, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Van Alstine, W.G.; Popielarczyk, M.; Albregts, S.R. Effect of formalin fixation on the immunohistochemical detection of PRRS virus antigen in experimentally and naturally infected pigs. J. Vet. Diagn. Investig. 2002, 14, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Yaeger, M.; Funk, N.; Hoffman, L. A survey of agents associated with neonatal diarrhea in Iowa swine including Clostridium difficile and porcine reproductive and respiratory syndrome virus. J. Vet. Diagn. Investig. 2002, 14, 281–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, J.; Kneucker, A.; Liebler-Tenorio, E.; Fachinger, V.; Glaser, M.; Pesch, S.; Murtaugh, M.P.; Reinhold, P. Respiratory function and pulmonary lesions in pigs infected with porcine reproductive and respiratory syndrome virus. Vet. J. 2011, 187, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Flores, J.; Calle, S.; Falcón, N.; Torres, M.; Morales, S.; Acosta, F. Evaluación de una bacterina de dosis única contra Mycoplasma hyopnemoniae en porcinos de madres no vacunadas. Rev. Investig. Vet. Peru 2006, 17, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Pepovich, R. Efficacy of single-dose Mycoplasma hyopneumoniae vaccine for the control of enzootic pneumonia in pigs. Bulg. J. Agric. Sci. 2019, 25, 1039–1043. [Google Scholar]
- Thacker, E.L.; Thacker, B.J.; Young, T.F.; Halbur, P.G. Effect of vaccination on the potentiation of porcine reproductive and respiratory syndrome virus (PRRSV)-induced pneumonia by Mycoplasma hyopneumoniae. Vaccine 2000, 18, 1244–1252. [Google Scholar] [CrossRef]
- Martelli, P.; Terreni, M.; Guazzetti, S.; Cavirani, S. Antibody response to Mycoplasma hyopneumoniae infection in vaccinated pigs with or without maternal antibodies induced by sow vaccination. J. Vet. Med. Ser. B 2006, 53, 229–233. [Google Scholar] [CrossRef]
- Hodgins, D.C.; Shewen, P.E.; Dewey, C.E. Influence of age and maternal antibodies on antibody responses of neonatal piglets vaccinated against Mycoplasma hyopneumoniae. J. Swine Health Prod. 2004, 12, 10–16. [Google Scholar]
- Maes, D.; Segales, J.; Meyns, T.; Sibila, M.; Pieters, M.; Haesebrouck, F. Control of Mycoplasma hyopneumoniae infections in pigs. Vet. Microbiol. 2008, 126, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Kraft, C.; Hennies, R.; Dreckmann, K.; Noguera, M.; Rathkjen, P.H.; Gassel, M.; Gereke, M. Evaluation of PRRSv specific, maternally derived and induced immune response in Ingelvac PRRSFLEX EU vaccinated piglets in the presence of maternally transferred immunity. PLoS ONE 2019, 14, 1–21. [Google Scholar] [CrossRef]
- Cho, J.G.; Dee, S.A. Porcine reproductive and respiratory syndrome virus. Theriogenology 2006, 66, 655–662. [Google Scholar] [CrossRef]
- Tomás, A.; Fernandes, L.T.; Valero, O.; Segalés, J. A meta-analysis on experimental infections with porcine circovirus type 2 (PCV2). Vet. Microbiol. 2008, 132, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Larochelle, R.; Magar, R.; D’Allaire, S. Comparative serologic and virologic study of commercial swine herds with and without postweaning multisystemic wasting syndrome. Can. J. Vet. Res. 2003, 67, 114–120. [Google Scholar] [PubMed]
- Rodríguez-Arrioja, G.M.; Segalés, J.; Calsamiglia, M.; Resendes, A.R.; Balasch, M.; Plana-Durán, J.; Casal, J.; Domingo, M. Dynamics of porcine circovirus type 2 infection in a herd of pigs with postweaning multisystemic wasting syndrome. Am. J. Vet. Res. 2002, 63, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Carasova, P.; Celer, V.; Takacova, K.; Trundova, M.; Molinkova, D.; Lobova, D.; Smola, J. The levels of PCV2 specific antibodies and viremia in pigs. Res. Vet. Sci. 2007, 83, 274–278. [Google Scholar] [CrossRef]
- Grau-Roma, L.; Fraile, L.; Segalés, J. Recent advances in the epidemiology, diagnosis and control of diseases caused by porcine circovirus type 2. Vet. J. 2011, 187, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.A.; Scheidt, A.B.; Teclaw, R.F.; Clark, L.K.; Knox, K.E.; Jordan, M. Association between growth indicators and volume of lesions in lungs from pigs at slaughter. Am. J. Vet. Res. 1992, 53, 2221–2223. [Google Scholar] [PubMed]
- Pagot, E.; Pommier, P.; Keïta, A. Relationship between growth during the fattening period and lung lesions at slaughter in swine. Rev. Med. Vet. 2007, 158, 253. [Google Scholar]
- Straw, B.E.; Tuovinen, V.K.; Bigras-Poulin, M. Estimation of the cost of pneumonia in swine herds. J. Am. Vet. Med. Assoc. 1989, 195, 1702–1706. [Google Scholar]
- Permentier, L.; Maenhout, D.; Deley, W.; Broekman, K.; Vermeulen, L.; Agten, S.; Verbeke, G.; Aviron, J.; Geers, R. Lung lesions increase the risk of reduced meat quality of slaughter pigs. Meat Sci. 2015, 108, 106–108. [Google Scholar] [CrossRef] [PubMed]
- Donkó, T.; Kovács, M.; Magyar, T. Association of growth performance with atrophic rhinitis and pneumonia detected at slaughter in a conventional pig herd in Hungary. Acta Vet. Hung. 2005, 53, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Dailidavičienė, J.; Januškevičienė, G.; Milius, J.; Mieželienė, A.; Alenčikienė, G. Influence of lung pathology on pig carcasses’ microbiological quality and sensory parameters. Bull. Vet. Inst. Puławy 2009, 53, 433–438. [Google Scholar]
- Arsenakis, I. Different Vaccination Protocols with Bacterins as a Means to Control Mycoplasma hyopneumoniae Infections in Peri-weaned and Fattening Pigs. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2018. [Google Scholar]
- Kritas, S.K.; Alexopoulos, C.; Kyriakis, C.S.; Tzika, E.; Kyriakis, S.C. Performance of fattening pigs in a farm infected with both porcine reproductive and respiratory syndrome (PRRS) virus and porcine circovirus type 2 following sow and piglet vaccination with an attenuated PRRS vaccine. J. Vet. Med. Ser. A 2007, 54, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Chiers, K.; Donné, E.; van Overbeke, I.; Ducatelle, R.; Haesebrouck, F. Actinobacillus pleuropneumoniae infections in closed swine herds: Infection patterns and serological profiles. Vet. Microbiol. 2002, 85, 343–352. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Category | Frequency (%) | p Value |
|---|---|---|---|
| Gross lesions (n = 108) | Animals with lesions | 79 (73.1%) | < 0.001 |
| Animals without lesions | 29 (26.9%) | ||
| Lung lesion patterns 1 (n = 79) | CBP | 50 (63.3%) | < 0.001 |
| Pleuritis | 19 (24.1%) | ||
| FNP | 7 (8.9%) | ||
| IP | 2 (2.5%) | ||
| EP | 1 (1.3%) | ||
| Percentage of affected lung parenchyma 1 (n = 79) | (0.1–10%) | 41 (51.9%) | < 0.001 |
| (10.1–25%) | 23 (29.1%) | ||
| (25.1–50%) | 7 (8.9%) | ||
| (50.1–75%) | 2 (2.5%) | ||
| (75.1–100%) | 6 (7.6%) |
| Global Sample (n = 108) | Animals with Lesions 1 (n = 76) | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Statistic | Animals without Lesions (n = 29) | Animals with Lesions (n = 79) | p Value | CBP (n = 50) | Pleuritis (n = 19) | FNP (n = 7) | p Value |
| Age of slaughter (days) | Mean (SD) | 200.8 (12.4) | 206 (11.4) | 0.035 | 206.7 (11) | 206 (12.2) | 201.4 (15.1) | 0.571 |
| Carcass weight (kg) | Mean (SD) | 77.7 (9.5) | 75.5 (8.5) | 0.248 | 75.5 (9) | 74.2 (7.7) | 80.6 (6.7) | 0.235 |
| Affected lung parenchyma(%) | Mean (SD) | - | 17.5 (25.3) | - | 16.4 (22.7) | 25.3 (35.1) | 8.5 (7.2) | 0.823 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paz-Sánchez, Y.; Herráez, P.; Quesada-Canales, Ó.; Poveda, C.G.; Díaz-Delgado, J.; Quintana-Montesdeoca, M.d.P.; Plamenova Stefanova, E.; Andrada, M. Assessment of Lung Disease in Finishing Pigs at Slaughter: Pulmonary Lesions and Implications on Productivity Parameters. Animals 2021, 11, 3604. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11123604
Paz-Sánchez Y, Herráez P, Quesada-Canales Ó, Poveda CG, Díaz-Delgado J, Quintana-Montesdeoca MdP, Plamenova Stefanova E, Andrada M. Assessment of Lung Disease in Finishing Pigs at Slaughter: Pulmonary Lesions and Implications on Productivity Parameters. Animals. 2021; 11(12):3604. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11123604
Chicago/Turabian StylePaz-Sánchez, Yania, Pedro Herráez, Óscar Quesada-Canales, Carlos G. Poveda, Josué Díaz-Delgado, María del Pino Quintana-Montesdeoca, Elena Plamenova Stefanova, and Marisa Andrada. 2021. "Assessment of Lung Disease in Finishing Pigs at Slaughter: Pulmonary Lesions and Implications on Productivity Parameters" Animals 11, no. 12: 3604. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11123604