
Effects of the Monomeric Components of Poly-hydroxybutyrate-co-hydroxyhexanoate on the Growth of Vibrio penaeicida In Vitro and on the Survival of Infected Kuruma Shrimp (Marsupenaeus japonicus)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pathogenic Bacterium and Growth Conditions
2.2. Preparation of Hydroxyalkanoic Acids and Fatty Acids
2.3. pH of the Shrimp Gut
2.4. Growth Inhibitory Effects in V. penaeicida
2.5. Shrimp Growth Experiments
2.6. In Vivo Pathogenic Bacterial Challenge Experiments
2.7. Statistics
3. Results
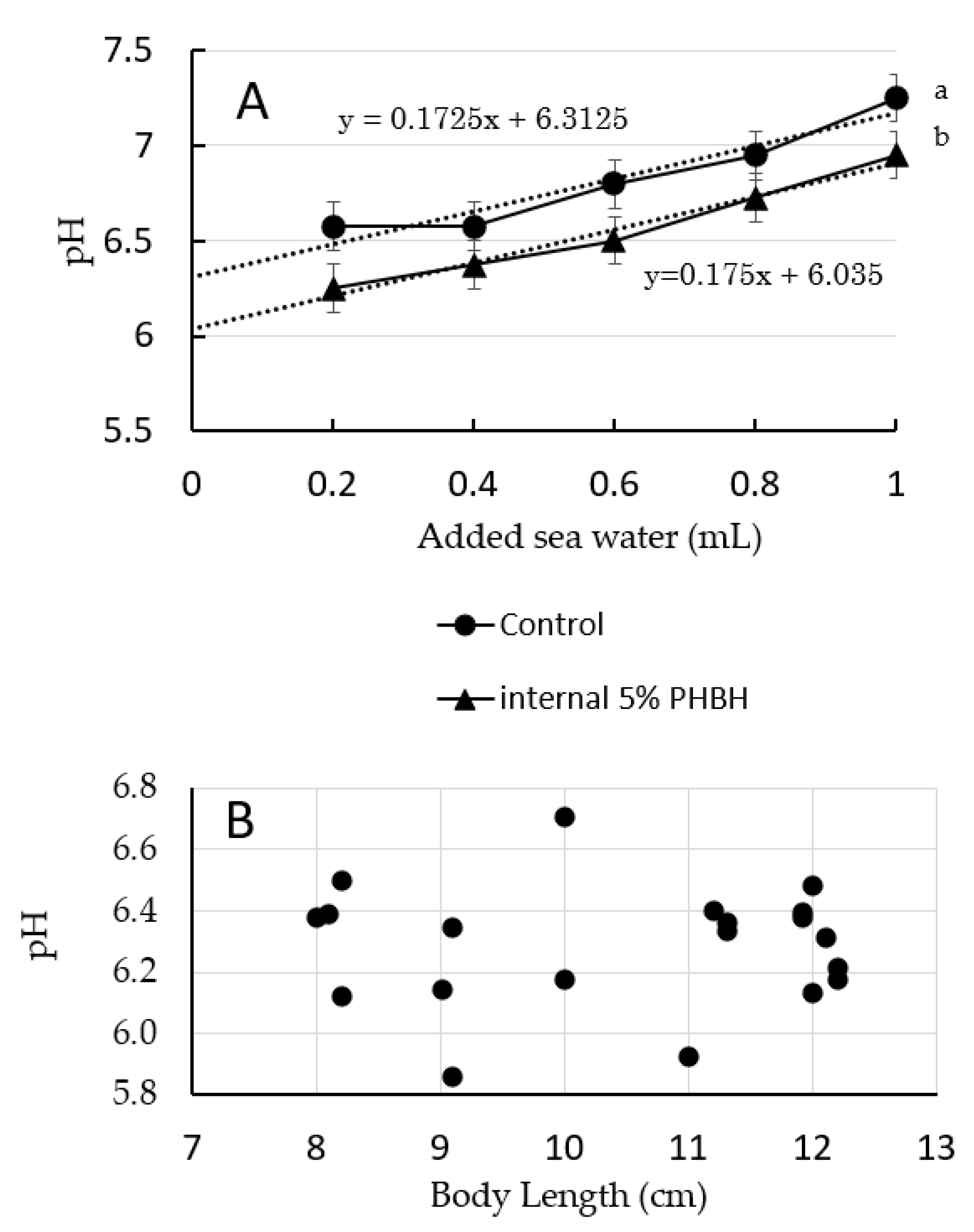
3.1. pH of the Shrimp Gut
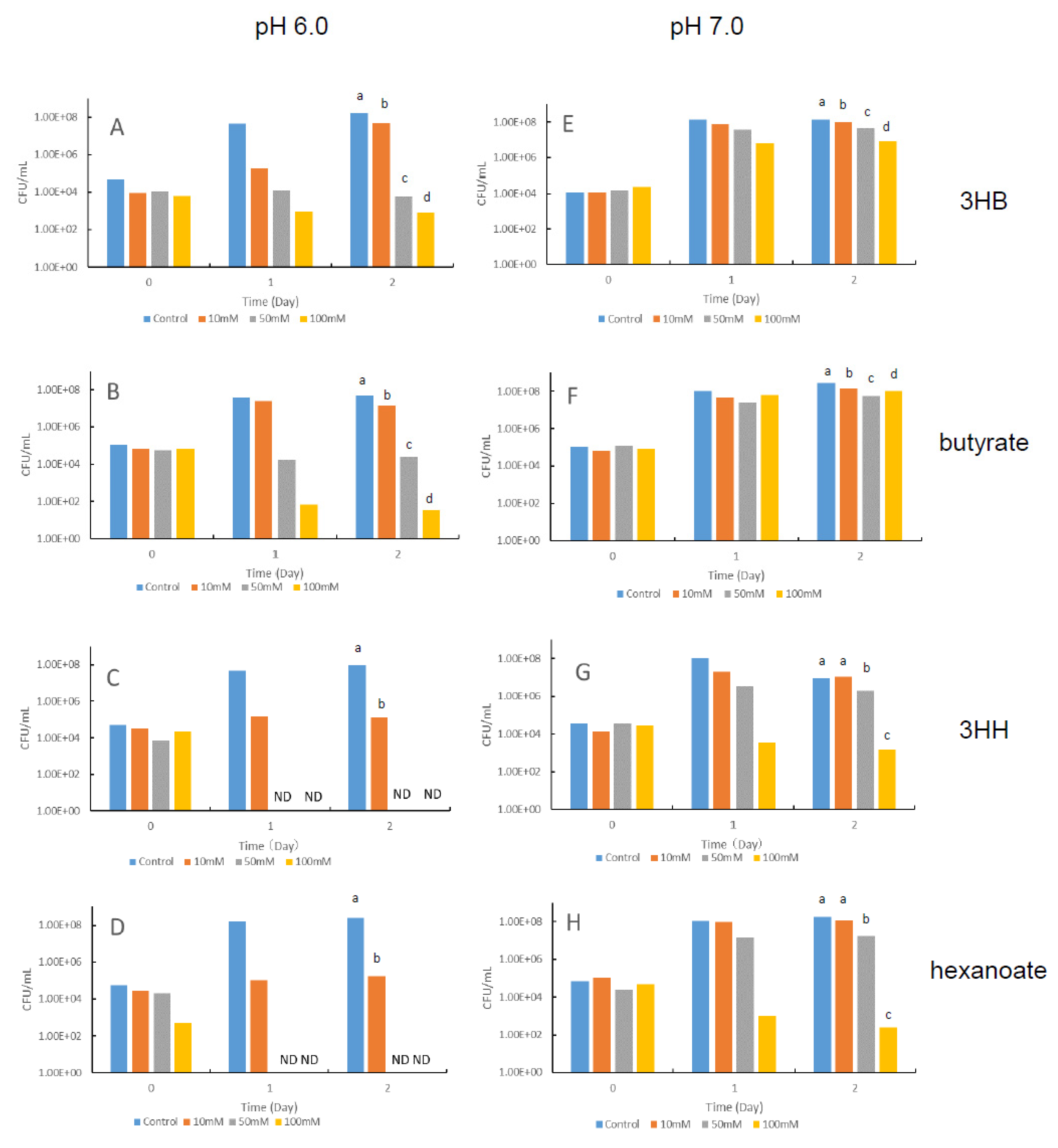
3.2. Effects of Hydroxyalkanoic Acids and Fatty Acids on the Growth of V. penaeicida
3.3. Effect of PHBH on the Growth of M. japonicus
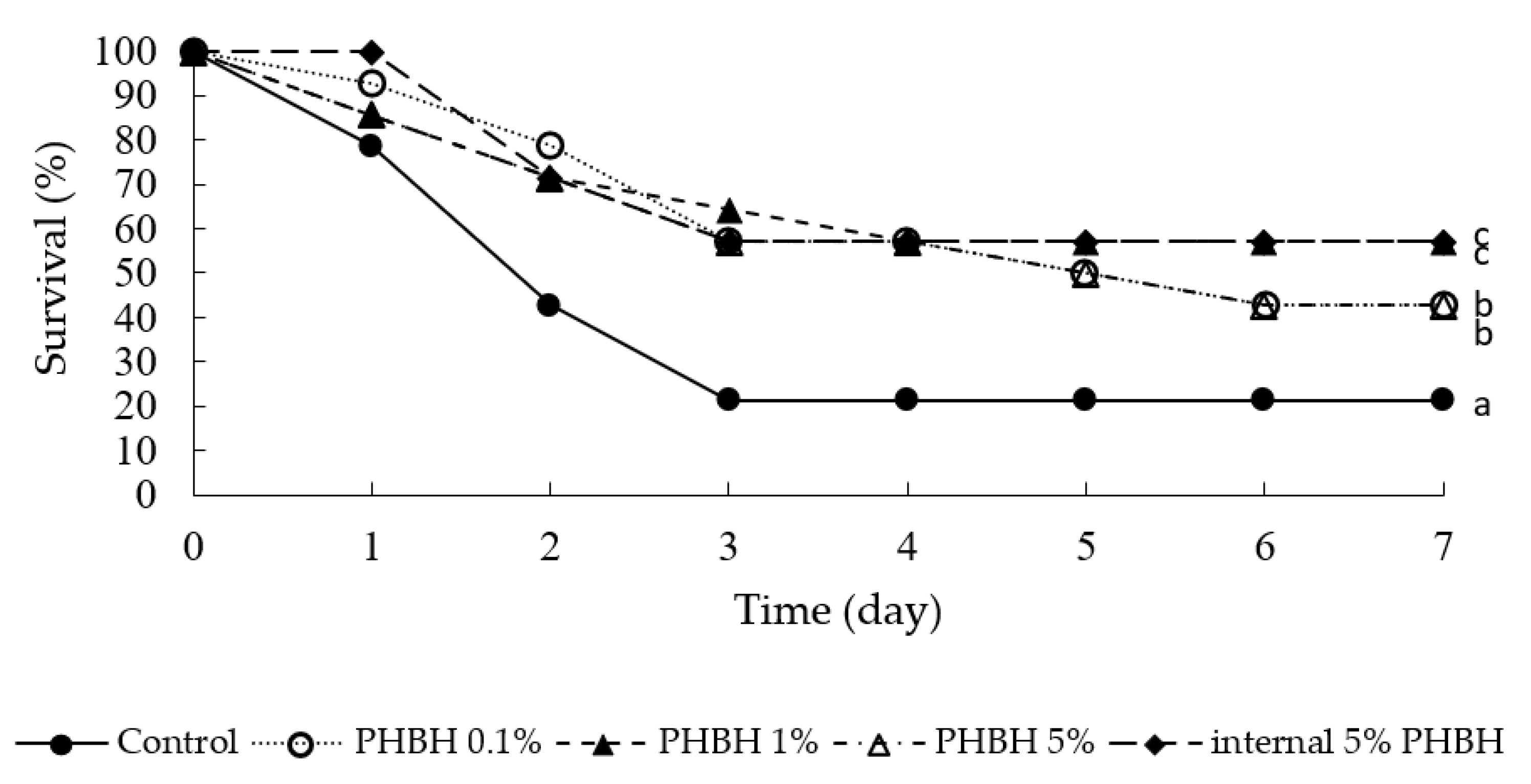
3.4. In Vivo Bacterial Challenge Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; 224p. [Google Scholar]
- Aksornkoae, S.; Tokrisna, R. Overview of shrimp farming and mangrove loss in Thailand. In Shrimp Farming and Mangrove Loss in Thailand; Barbier, E.B., Sathirathai, S., Eds.; Edward Elgar: Cheltenham, UK, 2004; pp. 37–51. [Google Scholar]
- Teeyaporn, K.; Fukami, K.; Songsangjinda, P.; Muangyao, P. Isolation and characterization of Noctiluca-killing bacteria from a shrimp aquaculture pond in Thailand. Fish. Sci. 2011, 77, 657–664. [Google Scholar]
- Ferreira, J.G.; Falconer, L.; Kittiwanich, J.; Ross, L.; Saurel, C.; Wellman, K.; Zhu, C.B.; Suvanachai, P. Analysis of production and environmental effects of Nile tilapia and white shrimp culture in Thailand. Aquaculture 2015, 447, 23–36. [Google Scholar] [CrossRef]
- Muangyao, P.; Fukami, K.; Songsangjinda, P.; Predalumpaburta, Y. Stimulation by gutweed to increase the abundance of insect larvae as food for shrimp aquaculture in Thailand. Aquaculture 2020, 519, 734740. [Google Scholar] [CrossRef]
- Momoyama, K.; Muroga, K. Diseases of Cultured Kuruma Shrimp in Japan: A Review. Fish Pathol. 2005, 40, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Annual Changes in Production of Kuruma Shrimp (Marsupenaeus Japonicus) in Japan. Available online: https://ieben.net/data/catch/sea-farm/japan-tdfk/kurumaebi.html (accessed on 18 February 2021).
- Sakai, T.; Hirae, T.; Yuasa, K.; Kamaishi, T.; Matsuyama, T.; Miwa, S.; Oseko, N.; Iida, T. Mass mortality of cultured kuruma prawn Penaeus japonicus caused by Vibrio nigripulchritudo. Fish. Pathol. 2007, 42, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Mandal, A.; Das, S.K. Comparative efficacy of neem (Azadirachta indica) and non-neem supplemented biofloc media in controlling the harmful luminescent bacteria in natural pond culture of Litopenaeus vannaemei. Aquaculture 2018, 492, 157–163. [Google Scholar] [CrossRef]
- Thompson, F.L.; Iida, T.; Swings, J. Biodiversity of vibrios. Microbiol. Mol. Biol. Rev. 2004, 68, 403–431. [Google Scholar] [CrossRef] [Green Version]
- Austin, B.; Zhang, X.H. Vibrio harveyi: A significant pathogen of marine vertebrates and invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef]
- Moriarty, D.J.W. Disease control in shrimp aquaculture with probiotic bacteria. In Microbial Biosystems: New Frontiers, Proceedings of the 8th International Symposium on Microbial Ecology, Halifax, NS, Canada, 1 August 1 1998; Bell, C.R., Brylinsky, M., Johnson-Green, P., Eds.; Altantic Canada Society for Microbial Ecology: Halifax, NS, Canada, 1999; pp. 237–243. Available online: http://ag.arizona.ed/azaqua/tilapia/tilapia_shrimp/moriarty.pdf (accessed on 18 February 2021).
- Decamp, O.; Moriarty, D.J.W.; Lavens, P. Probiotics for shrimp larviculture: Review of field data from Asia and Latin America. Aquac. Res. 2008, 39, 334–338. [Google Scholar] [CrossRef]
- Wang, H.; Wang, C.; Tang, Y.; Sun, B.; Huang, J.; Song, X. Pseudoalteromonas probiotics as potential biocontrol agents improve the survival of Penaeus vannamei challenged with acute hepatopancreatic necrosis disease (AHPND)-causing Vibrio parahaemolyticus. Aquaculture 2018, 494, 30–36. [Google Scholar] [CrossRef]
- Teo, J.W.P.; Tan, T.M.C.; Poh, C.L. Genetic determinants of tetracycline resistance in Vibrio harveyi. Antimicrob. Agents Chemother. 2002, 46, 1038–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, Y.; Gao, J.; Jiang, H.; Yao, J.; Gong, G.; Chen, X.; Xu, W.; He, X. Antimicrobial activity and virulence attenuation of citral against the fish pathogen Vibrio alginolyticus. Aquaculture 2020, 515, 734578. [Google Scholar] [CrossRef]
- Defoirdt, T.; Boon, N.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Short-chain fatty acids and poly-β-hydroxyalkanoates: (new) biocontrol agents for a sustainable animal production. Biotech. Adv. 2009, 27, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T.; Halet, D.; Sorgeloos, P.; Bossier, P.; Verstraete, W. Short-chain fatty acids protect gnotobiotic Artemia franciscana from pathogenic Vibrio campbellii. Aquaculture 2006, 261, 804–808. [Google Scholar] [CrossRef]
- Nhan, D.T.; Wille, M.; De Schryver, P.; Defoirdt, T.; Bossier, P.; Sorgeloos, P. The effect of poly-β-hydroxybutyrate on larviculture of the giant freshwater shrimp Macrobrachium rosenbergii. Aquaculture 2010, 302, 76–81. [Google Scholar] [CrossRef]
- Defoirdt, T.; Halet, D.; Vervaeren, H.; Boon, N.; Van De Wiele, T.; Sorgeloos, P.; Bossier, P.; Verstraete, W. The bacterial storage compound poly-ß-hydroxybutyrate protects Artemia franciscana, from pathogenic Vibrio campbellii. Environ. Microbiol. 2007, 9, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Suriyamongkol, P.; Weselake, R.; Narine, S.; Moloney, M.; Shah, S. Biotechnological approaches for the production of polyhydroxyalkanoates in microorganisms and plants—A review. Biotechnol. Adv. 2007, 25, 148–175. [Google Scholar] [CrossRef] [PubMed]
- Sui, L.; Cai, J.; Sun, H.; Wille, M.; Bossier, P. Effect of poly-β-hydroxybutyrate on Chinese mitten crab, Eriocheir sinensis, larvae challenged with pathogenic Vibrio anguillarum. J. Fish Dis. 2012, 35, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Leopoldo, J.; Laranja, Q.; Ludevese-Pascual, G.L.; Amar, E.C.; Sorgeloos, P.; Bossier, P.; De Schryver, P. Poly-β-hydroxybutyrate (PHB) accumulating Bacillus spp. Improve the survival, growth and robustness of Penaeus mondon (Fabricius, 1798) postlarvae. Vet. Microbiol. 2014, 173, 310–317. [Google Scholar]
- Thai, T.Q.; Wille, M.; Garcia-Gonzalez, L.; Sorgeloos, P.; Bossier, P.; De Schryver, P. Poly-ß-hydroxybutyrate content and dose of the bacterial carrier for Artemia enrichment determine the performance of giant freshwater shrimp larvae. Appl. Microbiol. Biotechnol. 2014, 98, 5205–5215. [Google Scholar] [CrossRef] [PubMed]
- Hung, N.V.; De Schryver, P.; Tran, T.T.; Garcia-Gonzalez, L.; Bossier, P.; Nevejan, N. Application of poly-β-hydroxybutyrate (PHB) in mussel larviculture. Aquaculture 2015, 446, 318–324. [Google Scholar] [CrossRef]
- Situmorang, M.L.; De Schryver, P.; Dierckens, K.; Bossier, P. Effect of poly-β-hydroxybutyrate on growth and disease resistance of Nile tilapia Oreochromis niloticus juveniles. Vet. Microbiol. 2016, 182, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Du, D.; Bo, Z.; Sui, L. Poly-βhydroxybutyrate (PHB)-accumulating Halomonas improves the survival, growth, robustness and modifies the gut microbial composition of Litopenaeus vannamei postlarvae. Aquaculture 2019, 500, 607–612. [Google Scholar] [CrossRef]
- Defoirdt, T.; Anh, N.T.M.; De Schryver, P. Virulence-inhibitory activity of the degradation product 3-hydroxybutyrate explains the protective effect of poly-β-hydroxybutyrate against the major aquaculture pathogen Vibrio campbellii. Sci. Rep. 2018, 8, 7245. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, E.; Kasuya, K.; Kobayashi, G.; Shiotani, T.; Shima, Y.; Doi, Y. Physical Properties and Biodegradability of Microbial Poly (3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules 1994, 27, 878–880. [Google Scholar] [CrossRef]
- Doi, Y.; Kitamura, S.; Abe, H. Microbial Synthesis and Characterization of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules 1995, 28, 4822–4828. [Google Scholar] [CrossRef]
- Fujiki, T. Biodegradable plastic production by microorganism. Microbiol. Cult. Coll. 2013, 29, 25–29. (In Japanese) [Google Scholar]
- Arikawa, H.; Matsumoto, K. Evaluation of gene expression cassettes and production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) with a fine modulated monomer composition by using it in Cupriavidus necator. Microb. Cell Fact. 2016, 15, 184. [Google Scholar] [CrossRef] [Green Version]
- Immanuel, G.; Sivagnanavelmurugan, M.; Palavesam, A. Antibacterial effect of short-chain fatty acids on gnotobiotic Artemia franciscana nauplii against Vibrio parahaemolyticus. Aquac. Res. 2012, 43, 518–525. [Google Scholar] [CrossRef]
- Hung, N.V.; Bossier, P.; Hong, N.T.X.; Ludeseve, C.; Garcia-Gonzalez, L.; Nevejan, N.; De Schryver, P. Does Ralstonia eutropha, rich in poly-β hydroxybutyrate (PHB), protect blue mussel larvae against pathogenic vibrios? J. Fish Dis. 2019, 42, 777–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukami, K.; Nishijima, T.; Hata, Y. Availability of deep seawater and effects of bacteria isolated from deep seawater on the mass culture of food microalga Chaetoceros ceratosporum. Nippon Suisan Gakkaishi 1992, 58, 931–936. [Google Scholar] [CrossRef]
- Ali, M.F.Z.; Yasin, I.A.; Ohta, T.; Hashizume, A.; Ido, A.; Takahashi, T.; Miura, C.; Miura, T. The silkrose of Bombyx mori effectively prevents vibriosis in penaeid prawns via the activation of innate immunity. Sci. Rep. 2018, 8, 8836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mine, S.; Boopathy, R. Effect of organic acids on shrimp pathogen, Vibrio harveyi. Curr. Microbiol. 2011, 63, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Sun, Y.-Z.; Caipang, C.M. Short-chain fatty acids as feed supplements for sustainable aquaculture: An updated view. Aquacul. Res. 2017, 48, 1380–1391. [Google Scholar] [CrossRef]
- De Schryver, P.; Shima, A.K.; Kunwar, P.S.; Baruah, K.; Verstraete, W.; Boon, N.; Boeck, G.; Bossier, P. Poly-β-hydroxybutyrate (PHB) increases growth performance and intestinal bacterial range-weighted richness in juvenile European sea bass, Dicentrarchs labrax. Appl. Microbiol. Biotechnol. 2010, 86, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Halet, D.; Defoirdt, T.; Van Damme, P.; Vervaeren, H.; Forrez, I.; Van de Wiele, T.; Boon, N.; Sorgeloos, P.; Bossier, P.; Verstraete, W. Poly-ß-hydroxybutyrate-accumulating bacteria protect gnotobiotic Artemia franciscana from pathogenic Vibrio campbellii. FEMS Microbiol. Ecol. 2007, 60, 363–369. [Google Scholar] [CrossRef] [PubMed]
- McHan, F.; Shotts, E.B. Effects of short-chain fatty acids on the growth of Salmonella tyrhimurium in an in vitro system. Avian Dis. 1993, 37, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Durant, J.A.; Lowry, V.K.; Nisbet, D.J.; Stanker, L.H.; Corrier, D.E.; Ricke, S.C. Late logarithmic Salmonella typhimurium HEp-2cell association and invasion response to short-chain fatty acid addition. J. Food Saf. 2000, 20, 1–11. [Google Scholar] [CrossRef]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Velge, P.; Botteau, E.; Fievez, V.; Haesebrouck, F.; Ducatelle, R. Invasion of Salmonella enteritidis in avian intestinal epithelial cells in vitro is influenced by short-chain fatty acids. Int. J. Food Microbiol. 2003, 85, 237–248. [Google Scholar] [CrossRef]
- Sun, C.Q.; O’ Connor, C.J.; Turner, S.J.; Lewis, G.D.; Stanley, R.A.; Roberton, A.M. The effect of pH on the inhibition of bacterial growth by physiological concentrations of butyric acid: Implications for neonates fed on suckled milk. Chem. Biol. Interact. 1998, 113, 117–131. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition | g/100g |
|---|---|
| Crude Protein | 57.0 |
| Crude Lipid | 9.6 |
| Fiber | 3.0 |
| Ash | 16.0 |
| Others | 5.8 |
| Water | 8.6 |
| Total | 100 |
| Feed Component | PHBH % | Internal 5% | |||
|---|---|---|---|---|---|
| Control | 0.1 | 1 | 5 | ||
| Standard Diet | 100 | 100 | 100 | 100 | 95 |
| PHBH | 0 | 0.1 | 1 | 5 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukami, K.; Takagi, F.; Sonoda, K.; Okamoto, H.; Kaneno, D.; Horikawa, T.; Takita, M. Effects of the Monomeric Components of Poly-hydroxybutyrate-co-hydroxyhexanoate on the Growth of Vibrio penaeicida In Vitro and on the Survival of Infected Kuruma Shrimp (Marsupenaeus japonicus). Animals 2021, 11, 567. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020567
Fukami K, Takagi F, Sonoda K, Okamoto H, Kaneno D, Horikawa T, Takita M. Effects of the Monomeric Components of Poly-hydroxybutyrate-co-hydroxyhexanoate on the Growth of Vibrio penaeicida In Vitro and on the Survival of Infected Kuruma Shrimp (Marsupenaeus japonicus). Animals. 2021; 11(2):567. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020567
Chicago/Turabian StyleFukami, Kimio, Fumika Takagi, Kohei Sonoda, Hiroshi Okamoto, Daisuke Kaneno, Takao Horikawa, and Masaki Takita. 2021. "Effects of the Monomeric Components of Poly-hydroxybutyrate-co-hydroxyhexanoate on the Growth of Vibrio penaeicida In Vitro and on the Survival of Infected Kuruma Shrimp (Marsupenaeus japonicus)" Animals 11, no. 2: 567. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020567