
Polymorphism Detection of GDF9 Gene and Its Association with Litter Size in Luzhong Mutton Sheep (Ovis aries)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and DNA Extraction
2.2. Full-Length Sequencing and Mutation Detection of GDF9 Coding Sequence
2.3. Genotyping of FecB Mutation
2.4. Statistical Analysis
2.5. Phylogenetic Analysis
2.6. Protein Structure Prediction
3. Results
3.1. Polymorphism Analysis of GDF9 Gene in Luzhong Sheep
3.2. Population Genetic Analysis of SNPs in GDF9 and FecB
3.3. Association Analysis between Eight Loci in GDF9 and FecB with Litter Size in Luzhong Sheep
3.4. Construction of Phylogenetic Tree of GDF9 Gene for 10 Sheep Breeds and Other Five Animal Species
3.5. Prediction of the Protein Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Al-Mutar, H.; Younis, L. Effect of point mutation in the growth differentiation factor 9 gene of oocytes on the sterility and fertility of Awassi sheep. Arch. Razi Inst. 2020, 75, 101–108. [Google Scholar] [CrossRef]
- Vage, D.I.; Husdal, M.; Kent, M.P.; Klemetsdal, G.; Boman, I.A. A missense mutation in growth differentiation factor 9 (GDF9) is strongly associated with litter size in sheep. BMC Genet. 2013, 14, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Hu, W.; Di, R.; Liu, Q.; Wang, X.; Zhang, X.; Zhang, J.; Chu, M. Expression analysis of the prolific candidate genes, BMPR1B, BMP15, and GDF9 in Small Tail Han ewes with three fecundity (FecB gene) genotypes. Animals 2018, 8, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, M.P.; Hanrahan, J.P. Direct evidence on the contribution of a missense mutation in GDF9 to variation in ovulation rate of Finnsheep. PLoS ONE 2014, 9, e95251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eghbalsaied, S.; Ghaedi, K.; Shahmoradi, S.; Pirestani, A.; Amini, H.; Saiedi, T.; Nicol, L.; McNeilly, A. Presence of SNPs in GDF9 mRNA of Iranian Afshari Sheep. Int. J. Fertil. Steril. 2012, 5, 225–230. [Google Scholar]
- Abdoli, R.; Ziaeddin Mirhoseini, S.; Ghavi Hossein-Zadeh, N.; Zamani, P. Screening for causative mutations of major prolificacy genes in Iranian Fat-Tailed sheep. Int. J. Fertil. Steril. 2018, 12, 51–55. [Google Scholar] [PubMed]
- Saleh, A.A.; Hammoud, M.H.; Dabour, N.A.; Hafez, E.E.; Sharaby, M.A. BMPR-1B, BMP-15 and GDF-9 genes structure and their relationship with litter size in six sheep breeds reared in Egypt. BMC Res. Notes 2020, 13, 215. [Google Scholar] [CrossRef] [Green Version]
- Bravo, S.; Larama, G.; Paz, E.; Inostroza, K.; Montaldo, H.H.; Sepulveda, N. Polymorphism of the GDF9 gene associated with litter size in Araucana Creole sheep. Anim. Genet. 2016, 47, 390–391. [Google Scholar] [CrossRef]
- Tong, B.; Wang, J.; Cheng, Z.; Liu, J.; Wu, Y.; Li, Y.; Bai, C.; Zhao, S.; Yu, H.; Li, G. Novel variants in GDF9 gene affect promoter activity and litter size in Mongolia sheep. Genes 2020, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Regan, S.L.; McFarlane, J.R.; O’Shea, T.; Andronicos, N.; Arfuso, F.; Dharmarajan, A.; Almahbobi, G. Flow cytometric analysis of FSHR, BMRR1B, LHR and apoptosis in granulosa cells and ovulation rate in merino sheep. Reproduction 2015, 150, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, G.W.; Lord, E.A.; Penty, J.M.; Dodds, K.G.; Broad, T.E.; Cambridge, L.; Sunden, S.L.; Stone, R.T.; Crawford, A.M. The Booroola fecundity (FecB) gene maps to sheep chromosome 6. Genomics 1994, 22, 148–153. [Google Scholar] [CrossRef]
- Jia, J.; Jin, J.; Chen, Q.; Yuan, Z.; Li, H.; Bian, J.; Gui, L. Eukaryotic expression, Co-IP and MS identify BMPR-1B protein-protein interaction network. Biol. Res. 2020, 53, 24. [Google Scholar] [CrossRef]
- Chu, M.X.; Liu, Z.H.; Jiao, C.L.; He, Y.Q.; Fang, L.; Ye, S.C.; Chen, G.H.; Wang, J.Y. Mutations in BMPR-IB and BMP-15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 2007, 85, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Mulsant, P. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Merino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, C.J.; MacDougall, C.; MacDougall, C.; Campbell, B.K.; McNeilly, A.S.; Baird, D.T. The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1 B (BMPR1B) gene. J. Endocrinol. 2001, 169, R1–R6. [Google Scholar] [CrossRef] [Green Version]
- Wilson, T.; Wu, X.Y.; Juengel, J.L.; Ross, I.K.; Lumsden, J.M.; Lord, E.A.; Dodds, K.G.; Walling, G.A.; McEwan, J.C.; O’Connell, A.R.; et al. Highly prolific Booroola sheep have a mutation in the intracellular kinase domain of bone morphogenetic protein IB receptor (ALK-6) that is expressed in both oocytes and granulosa cells. Biol. Reprod. 2001, 64, 1225–1235. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, Q.; Di, R.; Hu, W.; Wang, X.; He, X.; Ma, L.; Chu, M. Single nucleotide polymorphisms in BMP2 and BMP7 and the association with litter size in Small Tail Han sheep. Anim. Reprod. Sci. 2019, 204, 183–192. [Google Scholar] [CrossRef]
- Yasuda, N. HLA polymorphism information content (PIC). Jpn. J. Hum. Genet. 1988, 33, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Feng, T.; Geng, C.X.; Lang, X.Z.; Chu, M.X.; Cao, G.L.; Di, R.; Fang, L.; Chen, H.Q.; Liu, X.L.; Li, N. Polymorphisms of caprine GDF9 gene and their association with litter size in Jining Grey goats. Mol. Biol. Rep. 2011, 38, 5189–5197. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.M.; Freeny, A.; Heiberger, R.M. Analysis of variance. Designed experiments. In Statistical Models in S; Chambers, J.M., Hastie, T.J., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 145–193. [Google Scholar]
- Trifinopoulos, J.; Nguyen, L.T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Z.; Li, S.; Liu, Q.; Wang, Z.; Zhou, Z.; Di, R.; Miao, B.; Hu, W.; Wang, X.; Hu, X.; et al. Whole-genome sequences of 89 Chinese sheep suggest role of RXFP2 in the development of unique horn phenotype as response to semi-feralization. GigaScience 2018, 7, giy019. [Google Scholar] [CrossRef]
- Geourjon, C.; Deléage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Comput. Appl. Biosci. CABIOS 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef] [Green Version]
- UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [CrossRef] [PubMed] [Green Version]
- Demars, J.; Fabre, S.; Sarry, J.; Rossetti, R.; Gilbert, H.; Persani, L.; Tosser-Klopp, G.; Mulsant, P.; Nowak, Z.; Drobik, W.; et al. Genome-wide association studies identify two novel BMP15 mutations responsible for an atypical hyperprolificacy phenotype in sheep. PLoS Genet. 2013, 9, e1003482. [Google Scholar] [CrossRef]
- Lassoued, N.; Benkhlil, Z.; Woloszyn, F.; Rejeb, A.; Aouina, M.; Rekik, M.; Fabre, S.; Bedhiaf-Romdhani, S. FecX (Bar) a Novel BMP15 mutation responsible for prolificacy and female sterility in Tunisian Barbarine Sheep. BMC Genet. 2017, 18, 43. [Google Scholar] [CrossRef] [Green Version]
- Zamani, P.; Nadri, S.; Saffaripour, R.; Ahmadi, A.; Dashti, F.; Abdoli, R. A new mutation in exon 2 of the bone morphogenetic protein 15 gene is associated with increase in prolificacy of Mehraban and Lori sheep. Trop. Anim. Health Prod. 2015, 47, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.J.; McNeilly, A.S.; Benavides, M.V.; Melo, E.O.; Moraes, J.C. Mutation in the protease cleavage site of GDF9 increases ovulation rate and litter size in heterozygous ewes and causes infertility in homozygous ewes. Anim. Genet. 2014, 45, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Drouilhet, L.; Mansanet, C.; Sarry, J.; Tabet, K.; Bardou, P.; Woloszyn, F.; Lluch, J.; Harichaux, G.; Viguie, C.; Monniaux, D.; et al. The highly prolific phenotype of Lacaune sheep is associated with an ectopic expression of the B4GALNT2 gene within the ovary. PLoS Genet. 2013, 9, e1003809. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.; Raoul, J.; Bodin, L. Effects of the FecL major gene in the Lacaune meat sheep population. Genet. Sel. Evol. 2014, 46, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, G.H.; Dodds, K.G.; Wheeler, R.; Jay, N.P. Evidence that an imprinted gene on the X chromosome increases ovulation rate in sheep. Biol. Reprod. 2001, 64, 216–221. [Google Scholar] [CrossRef] [Green Version]
- Al-Samerria, S.; Al-Ali, I.; McFarlane, J.R.; Almahbobi, G. The impact of passive immunisation against BMPRIB and BMP4 on follicle development and ovulation in mice. Reproduction 2015, 149, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Lima, I.M.; Brito, I.R.; Rossetto, R.; Duarte, A.B.; Rodrigues, G.Q.; Saraiva, M.V.; Costa, J.J.; Donato, M.A.; Peixoto, C.A.; Silva, J.R.; et al. BMPRIB and BMPRII mRNA expression levels in goat ovarian follicles and the in vitro effects of BMP-15 on preantral follicle development. Cell Tissue Res. 2012, 348, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Juengel, J.L.; Davis, G.H.; McNatty, K.P. Using sheep lines with mutations in single genes to better understand ovarian function. Reproduction 2013, 146, R111–R123. [Google Scholar] [CrossRef] [PubMed]
- Fabre, S.; Pierre, A.; Pisselet, C.; Mulsant, P.; Lecerf, F.; Pohl, J.; Monget, P.; Monniaux, D. The Booroola mutation in sheep is associated with an alteration of the bone morphogenetic protein receptor-IB functionality. J. Endocrinol. 2003, 177, 435–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Wang, X.; Di, R.; Liu, Q.; Hu, W.; He, X.; Yu, J.; Zhang, X.; Zhang, J.; Broniowska, K.; et al. Metabolic effects of FecB gene on follicular fluid and ovarian vein serum in sheep (Ovis aries). Int J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; La, Y.; Zhou, X.; Zhang, X.; Li, F.; Liu, B. The genetic polymorphisms of TGFbeta superfamily genes are associated with litter size in a Chinese indigenous sheep breed (Hu sheep). Anim. Reprod. Sci. 2018, 189, 19–29. [Google Scholar] [CrossRef]
- Wang, W.; Liu, S.; Li, F.; Pan, X.; Li, C.; Zhang, X.; Ma, Y.; La, Y.; Xi, R.; Li, T. Polymorphisms of the ovine BMPR-IB, BMP-15 and FSHR and their associations with litter size in two chinese indigenous sheep breeds. Int. J. Mol. Sci. 2015, 16, 11385–11397. [Google Scholar] [CrossRef] [Green Version]
- Stocker, W.A.; Walton, K.L.; Richani, D.; Chan, K.L.; Beilby, K.H.; Finger, B.J.; Green, M.P.; Gilchrist, R.B.; Harrison, C.A. A variant of human growth differentiation factor-9 that improves oocyte developmental competence. J. Biol. Chem. 2020, 295, 7981–7991. [Google Scholar] [CrossRef]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Nicol, L.; Bishop, S.C.; Pong-Wong, R.; Bendixen, C.; Holm, L.E.; Rhind, S.M.; McNeilly, A.S. Homozygosity for a single base-pair mutation in the oocyte-specific GDF9 gene results in sterility in Thoka sheep. Reproduction 2009, 138, 921–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Ruiz, E.; Gallegos-Sanchez, J.; Cortez-Romero, C.; Segura-Leon, O.L.; Salinas-Ruiz, J.; Salazar-Ortiz, J. FecG(E) mutation in pelibuey sheep. Anim. Genet. 2020, 51, 346–347. [Google Scholar] [CrossRef] [PubMed]
- El Fiky, Z.A.; Hassan, G.M.; Nassar, M.I. Genetic polymorphism of growth differentiation factor 9 (GDF9) gene related to fecundity in two Egyptian sheep breeds. J. Assist. Reprod. Genet. 2017, 34, 1683–1690. [Google Scholar] [CrossRef]
- Heaton, M.P.; Smith, T.P.L.; Freking, B.A.; Workman, A.M.; Bennett, G.L.; Carnahan, J.K.; Kalbfleisch, T.S. Using sheep genomes from diverse U.S. breeds to identify missense variants in genes affecting fecundity. F1000Research 2017, 6, 1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.N.; Zhang, K.; Xu, T.M. The role of BMP15 and GDF9 in the pathogenesis of primary ovarian insufficiency. Hum. Fertil. 2019, 1–8. [Google Scholar] [CrossRef]
- Roy, S.; Gandra, D.; Seger, C.; Biswas, A.; Kushnir, V.A.; Gleicher, N.; Kumar, T.R.; Sen, A. Oocyte-derived factors (GDF9 and BMP15) and FSH regulate AMH expression via modulation of H3K27AC in granulosa cells. Endocrinology 2018, 159, 3433–3445. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Li, Q.; Wigglesworth, K.; Rangarajan, A.; Kattamuri, C.; Peterson, R.T.; Eppig, J.J.; Thompson, T.B.; Matzuk, M.M. Growth differentiation factor 9: Bone morphogenetic protein 15 heterodimers are potent regulators of ovarian functions. Proc. Natl. Acad. Sci. USA 2013, 110, E776–E785. [Google Scholar] [CrossRef] [Green Version]
- Monestier, O.; Servin, B.; Auclair, S.; Bourquard, T.; Poupon, A.; Pascal, G.; Fabre, S. Evolutionary origin of bone morphogenetic protein 15 and growth and differentiation factor 9 and differential selective pressure between mono- and polyovulating species. Biol. Reprod. 2014, 91, 83. [Google Scholar] [CrossRef]
- Ahmadi, A.; Afraz, F.; Talebi, R.; Farahavar, A.; Vahidi, S.M.F. Investigation of GDF9 and BMP15 polymorphisms in Mehraban sheep to find the missenses as impact on protein. Iran. J. Appl. Anim. Sci. 2016, 6, 863–872. [Google Scholar]




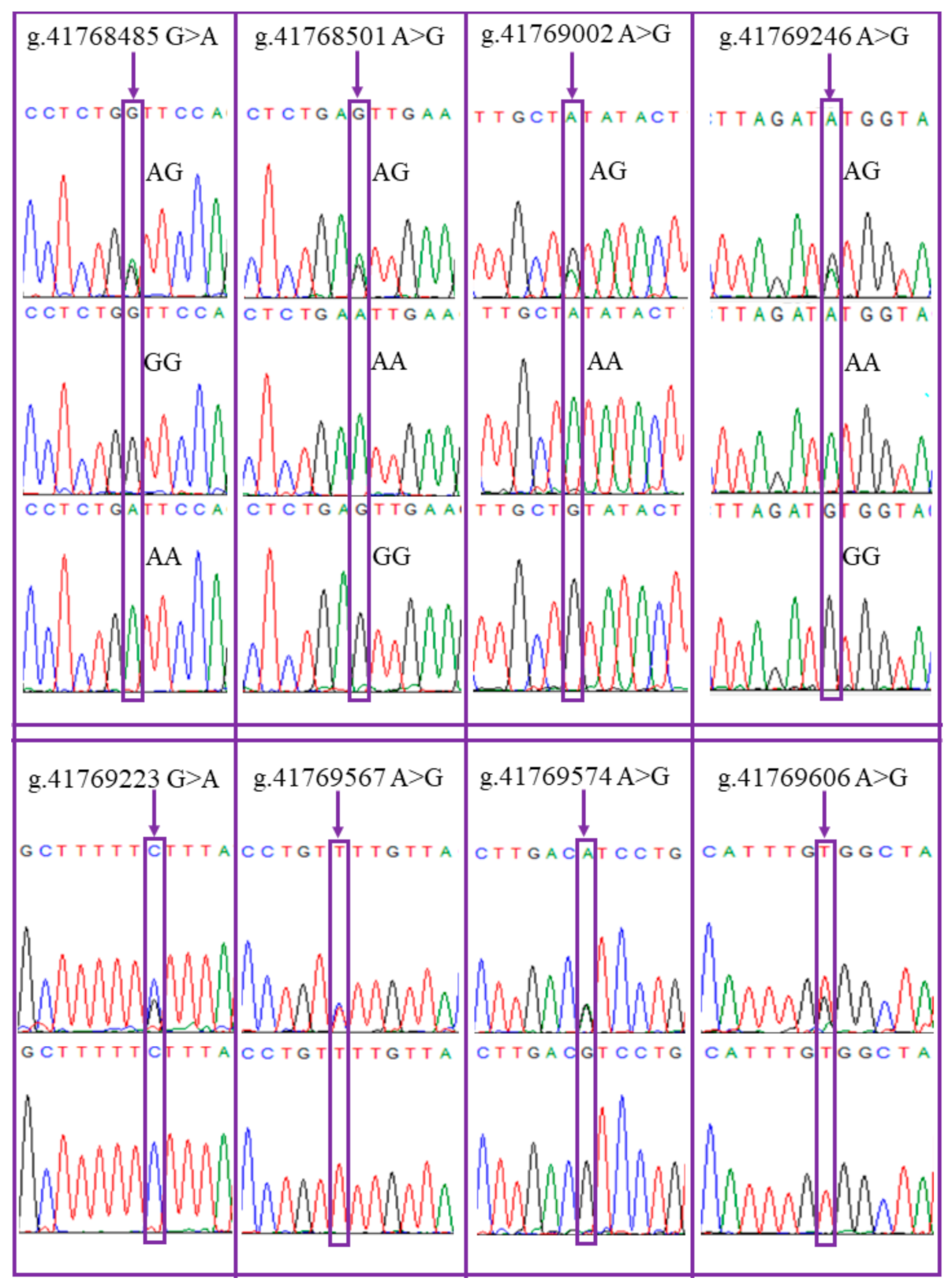{kind=link}
{kind=link}
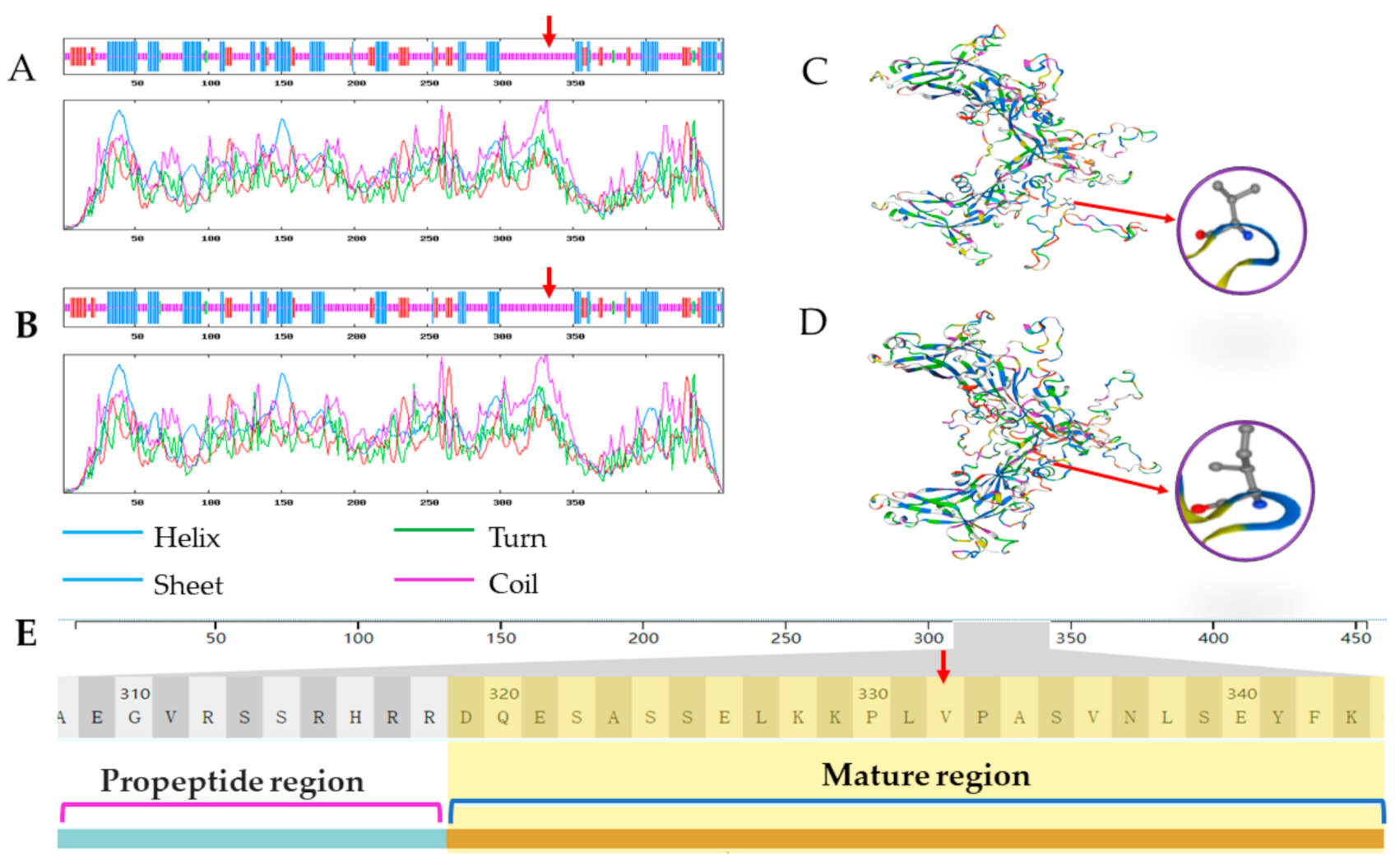{kind=link}
| Primer Name | Primer Sequence (5′–3′) | Annealing Temperature/°C | Amplified Fragment/bp |
|---|---|---|---|
| GDF9-F1 | ACTGAATGAATAGGGTGTTG | 58 | 1553 |
| GDF9-R1 | ATCTGTACCATATCTAAGTCC | 57 | |
| GDF9-F2 | GTTTTGTTACAGTGGGTTTAGAGC | 63 | 1507 |
| GDF9-R2 | GGCGCGGCATTTACACTGG | 58 | |
| GDF9-F3 | TCAGCTGAAGTGGGACAA | 60 | 694 |
| GDF9-R3 | ACACAGAAAATTTATGCCACTCAC | 62 |
| Region | Genomic Location (Chr5: Oar_v4.0/Oar_v3.1) | Wild-Type | Mutant | Mutation in Ensembl Database | AA Coding Residue | Amino Acid Change |
|---|---|---|---|---|---|---|
| Intron 1 | 41769606/41842523 | T | G | No | - | - |
| 41769574/41842491 | G | A | Yes | - | - | |
| 41769567/41842484 | T | C | Yes | - | - | |
| 41769246/41842163 | A | G | Yes | - | - | |
| 41769223/41842140 | C | G | Yes | - | - | |
| Exon 2 | 41769002/41841919 | A | G | Yes | 159 | Unchanged Leu (L) |
| 41768501/41841418 | A | G | Yes | 326 | Unchanged Glu (E) | |
| 41768485/41841402 | G | A | Yes | 332 | Val (V)–Ile (I) |
| Gene | Locus | Genotype Frequency | Allele Frequency | PIC | He | Ne | Chi-Square Test (p-Value) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| GDF9 | g.41769606 T > G | TT | GT | GG | T | G | 0.013 | 0.013 | 1.013 | 0.935 |
| 0.987 | 0.013 | 0.000 | 0.994 | 0.007 | ||||||
| g.41769574 G > A | GG | AG | AA | G | A | 0.013 | 0.013 | 1.013 | 0.935 | |
| 0.987 | 0.013 | 0.000 | 0.994 | 0.007 | ||||||
| g.41769567 T > C | TT | CT | CC | T | C | 0.083 | 0.087 | 1.095 | 0.555 | |
| 0.909 | 0.091 | 0.000 | 0.955 | 0.046 | ||||||
| g.41769246 A > G | AA | AG | GG | A | G | 0.325 | 0.408 | 1.690 | 0.821 | |
| 0.078 | 0.416 | 0.506 | 0.286 | 0.714 | ||||||
| g.41769223 C > G | CC | GC | GG | C | G | 0.083 | 0.087 | 1.095 | 0.555 | |
| 0.909 | 0.091 | 0.000 | 0.955 | 0.046 | ||||||
| g.41769002 A > G | AA | AG | GG | A | G | 0.325 | 0.408 | 1.690 | 0.821 | |
| 0.078 | 0.416 | 0.506 | 0.286 | 0.714 | ||||||
| g.41768501 A > G | AA | AG | GG | A | G | 0.229 | 0.263 | 1.357 | 0.874 | |
| 0.714 | 0.260 | 0.026 | 0.844 | 0.156 | ||||||
| g.41768485 G > A | GG | AG | AA | G | A | 0.229 | 0.263 | 1.357 | 0.874 | |
| 0.714 | 0.260 | 0.026 | 0.844 | 0.156 | ||||||
| FecB | g.29315643 A > G | AA | AG | GG | A | G | 0.340 | 0.434 | 1.766 | 0.005 |
| 0.416 | 0.532 | 0.052 | 0.682 | 0.318 | ||||||
| Gene | Locus | Genotype | Litter Size (Mean ± SD) |
|---|---|---|---|
| GDF9 | g.41769606 T > G | TT | 1.612 a ± 0.680 |
| GT | 2.000 a ± 0.000 | ||
| g.41769574 G > A | GG | 1.612 a ± 0.680 | |
| AG | 2.000 a ± 0.000 | ||
| g.41769567 T > C | TT | 1.614 a ± 0.684 | |
| CT | 1.642 a ± 0.621 | ||
| g.41769223 C > G | CC | 1.614 a ± 0.684 | |
| CG | 1.642 a ± 0.621 | ||
| g.41769246 A > G | AA | 1.750 a ± 0.608 | |
| GG | 1.603 a ± 0.688 | ||
| AG | 1.609 a ± 0.679 | ||
| g.41769002 A > G | AA | 1.750 a ± 0.608 | |
| GG | 1.603 a ± 0.688 | ||
| AG | 1.609 a ± 0.679 | ||
| g.41768501 A > G | AA | 1.564 a ± 0.656 | |
| AG | 1.825 a ± 0.708 | ||
| GG | 1.000 b ± 0.000 | ||
| g.41768485 G > A | GG | 1.564 a ± 0.656 | |
| AG | 1.825 a ± 0.708 | ||
| AA | 1.000 b ± 0.000 | ||
| FecB | g.29315643 A > G | AA | 1.094 b ± 0.342 |
| AG | 1.988 a ± 0.596 | ||
| GG | 2.000 a ± 0.730 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Chu, M.; Pan, L.; Wang, X.; He, X.; Zhang, R.; Tao, L.; La, Y.; Ma, L.; Di, R. Polymorphism Detection of GDF9 Gene and Its Association with Litter Size in Luzhong Mutton Sheep (Ovis aries). Animals 2021, 11, 571. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020571
Wang F, Chu M, Pan L, Wang X, He X, Zhang R, Tao L, La Y, Ma L, Di R. Polymorphism Detection of GDF9 Gene and Its Association with Litter Size in Luzhong Mutton Sheep (Ovis aries). Animals. 2021; 11(2):571. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020571
Chicago/Turabian StyleWang, Fengyan, Mingxing Chu, Linxiang Pan, Xiangyu Wang, Xiaoyun He, Rensen Zhang, Lin Tao, Yongfu La, Lin Ma, and Ran Di. 2021. "Polymorphism Detection of GDF9 Gene and Its Association with Litter Size in Luzhong Mutton Sheep (Ovis aries)" Animals 11, no. 2: 571. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11020571